黄河三角洲芦苇湿地生态系统碳通量动态特征及其影响因素
2021-08-07王永志刘胜林
王永志 ,刘胜林
1.河南科技大学应用工程学院,河南 三门峡 472000;2.四川农业大学管理学院,四川 成都 611130;3.三门峡职业技术学院,河南 三门峡 472000
湿地是陆地生态系统的重要组成部分。全球现有湿地面积占陆地总面积的2%—6%,但是,储存在湿地泥炭中的碳总量为 120—260 Pg,约占地球碳总量的15%(韩广轩,2017;陈小平等,2018;罗琪等,2017)。全球碳循环中,湿地生态系统作为全球生态系统的重要类型,其碳循环及碳收支的动态变化研究在全球碳收支平衡中扮演着重要角色(Duman et al.,2018)。湿地作为陆地生态系统最重要的碳库之一,虽然仅占陆地表面的6%左右,但其土壤碳储量占陆地土壤总碳储量的 10%—30%,其碳贮存能够消减大气日益增加的 CO2,在稳定全球气候、减缓温室效应方面发挥着重要作用(Meng et al.,2016;Cao et al.,2017)。然而,近年来由于气候变化和人类活动的干扰,湿地面积大幅度萎缩,其正常的生态系统碳循环过程也发生了巨大的变化(Miao et al.,2017)。因此,研究不同湿地的碳收支状况及其运行机制对精确估算全球碳收支具有重要意义。
目前已有的关于湿地方面的碳源/汇的研究尚无统一的结论(Duman et al.,2018;Meng et al.,2016)。大部分的研究结果证明湿地生态系统是碳汇,但随着气候环境变化以及人类活动的影响,部分湿地呈现出碳源的现象。芦苇湿地作为世界上分布最广泛的湿地类型之一,在湿地碳收支的研究中占有重要的地位(Zhuang et al.,2015;Pugh et al.,2018)。已经开展的关于芦苇湿地的研究极少采用直接观测CO2在大气和湿地之间的交换量,这就造成了无法定量分析CO2交换量与环境因子之间变化的相关关系(Zhong et al.,2016)。随着涡度相关技术的发展,使得直接测定陆地生态系统与大气间的CO2和水热通量成为可能。近年来,涡度相关技术已经成为直接测定大气与群落CO2交换通量的主要方法,也是世界上CO2和水热通量测定的标准方法,所观测的数据已经成为检验各种模型估算精度的最权威的资料(Neubauer et al.,2019;Fellman et al.,2017)。该方法已经得到微气象学家和生态学家的广泛认可,成为目前通量观测网络 FlUXNET的主要技术手段,已经在世界范围内被广泛用来测量大气和地球表面碳、水、热通量的交换,用这种微气象学方法观测到的净生态系统 CO2交换(Net ecosystem CO2exchange,NEE)能够为在生态系统尺度上了解光合、呼吸提供重要信息。
国外对湿地净生态系统CO2交换的研究已有很多报道(Wilson et al.,2018;Chu et al.,2018;Han et al.,2015)。中国的湿地生态系统碳收支研究主要集中在青藏高原的若尔盖高原草丛湿地、三江平原草丛湿地等河口和内陆湿地,但是对黄河三角洲湿地碳通量的研究相对较少。黄河三角洲地区湿地面积大,类型多,结构复杂,独特的自然地理位置和气候特征使该地区蕴藏着丰富的湿地资源,是世界上生物多样性最丰富的地区之一(Strachan et al.,2015)。由于自身的典型性和特殊性,加之地貌和人为作用,该地区发育了多种多样的湿地生态系统,成为陆-海相互作用研究的热点地区(Hanis et al.,2015)。前人对黄河三角洲湿地生态系统的研究主要集中在生态系统植物群落分布、生态系统演变以及人类活动的影响上,对湿地净生态系统CO2交换(NEE)的研究还鲜有报道。为了更深入地了解黄河三角洲芦苇湿地碳的生物地球化学循环特征及其关键机制,本研究选取黄河三角洲芦苇湿地作为研究对象,结合涡度相关技术,利用长期的通量观测数据和生物量等野外监测数据,探讨生态系统尺度芦苇湿地净生态系统CO2交换量的季节变异特征及其环境控制机制,希望能为区域的碳收支预算和为全球碳循环模型的进一步完善提供理论基础,为重新评价芦苇湿地对全球变化的贡献提供重要的科学依据。
1 材料与方法
1.1 研究区概况
研究区位于黄河三角洲国家自然保护区(37°40′—38°10′N,118°41′—119°16′E)。该湿地自然保护区地处中国山东省东营市黄河入海口,总面积15.3×104hm2,是以保护黄河口新生湿地生态系统和珍稀濒危鸟类为主体的自然保护区。研究区四季分明,属北温带亚湿润气候区,年平均气温12.1 ℃,年均降水量551.6 mm,无霜期196 d,年蒸发量为1962 mm。土壤类型为潮土、盐土和滨海盐土,土壤表层多以轻沙壤土和沙壤土为主。有机质含量一般在0.6%—1.0%之间,土壤pH值为7.6—8.5。主要植被有芦苇(Phragmites australis)、穗状狐尾藻(Myriophyllum spicatum)、荻(Triarrhena sacchariflora)、蒲草(Typha angustifolia)、补血草(Limonium sinense)、翅碱蓬(Suaeda salsa)、柽柳(Tamarix chinensis)等,其中芦苇、翅碱蓬和柽柳分布较广(Strachan et al.,2015)。
1.2 观测方法与数据处理
通量观测设备主要包括一套开路涡度相关系统和常规气象要素测量系统。涡度相关系统主要测量离地面4.5 m高的CO2通量、潜热和感热通量,由一个开路远红外CO2/H2O气体分析仪(IRGA,LI 7500,LI-COR Inc.NE,USA)和一个三维超声波测风仪(CSAT3,Campbell Scientific,MS,USA)组成。仪器采样频率为10 Hz,每半小时自动将平均值记录在数据采集器中(CR5000,Campbell Scientific)。
常规气象要素测量系统包括安装在离地面 1.5 m的辐射测定仪(CNR-1,NY,USA)和光量子测定仪(LI190SB,Li-COR,Lincoln,NE,USA),用于测量净辐射和光合有效辐射。同时在离地面4.5 m处测量相对湿度(HMP45C,Vaisala,Woburn,MA,USA)和风速。土壤温度(地面以下 0.05、0.10、0.20、0.5、1.0 m)、土壤热通量(0.05 m,HFP01,HUKESEFLUX,Delft,Netherlands),降水量等要素也同时监测。每半小时输出1组平均值记录在数据采集器中。
为了减少因观测引起的不确定性,我们对数据进行了质量控制和处理。利用涡度相关数据处理软件(Edire软件)对数据进行坐标轴旋转和WPL校正,以消除地形倾斜对通量计算的影响,同时也校正了由于空气水热传输引起的 CO2和水汽密度波动造成的通量计算误差。由于降雨、标定和仪器故障(如系统维护、电压不稳、断电等)等原因必然会造成数据缺失和一些异常点的出现。同时,较低的摩擦风速(v)和夜间低湍流也会低估系统的净 CO2交换速率。为了消除这些影响,对数据进行了筛选和质量控制,大约30%的数据被剔除掉(Li et al.,2016)。
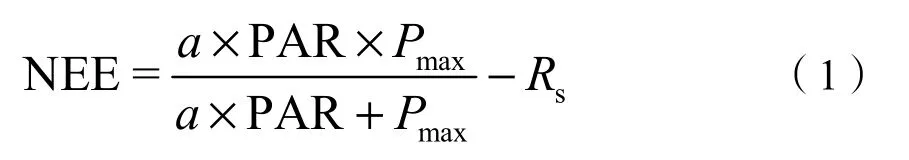
式中:a(mg·μmol−1)表示生态系统表观光量子效率,Pmax表示最大CO2同化效率,Rs表示生态系统呼吸速率(mg·μmol−1),PAR 表示光合有效辐射(μmol·m−2·s−1)。
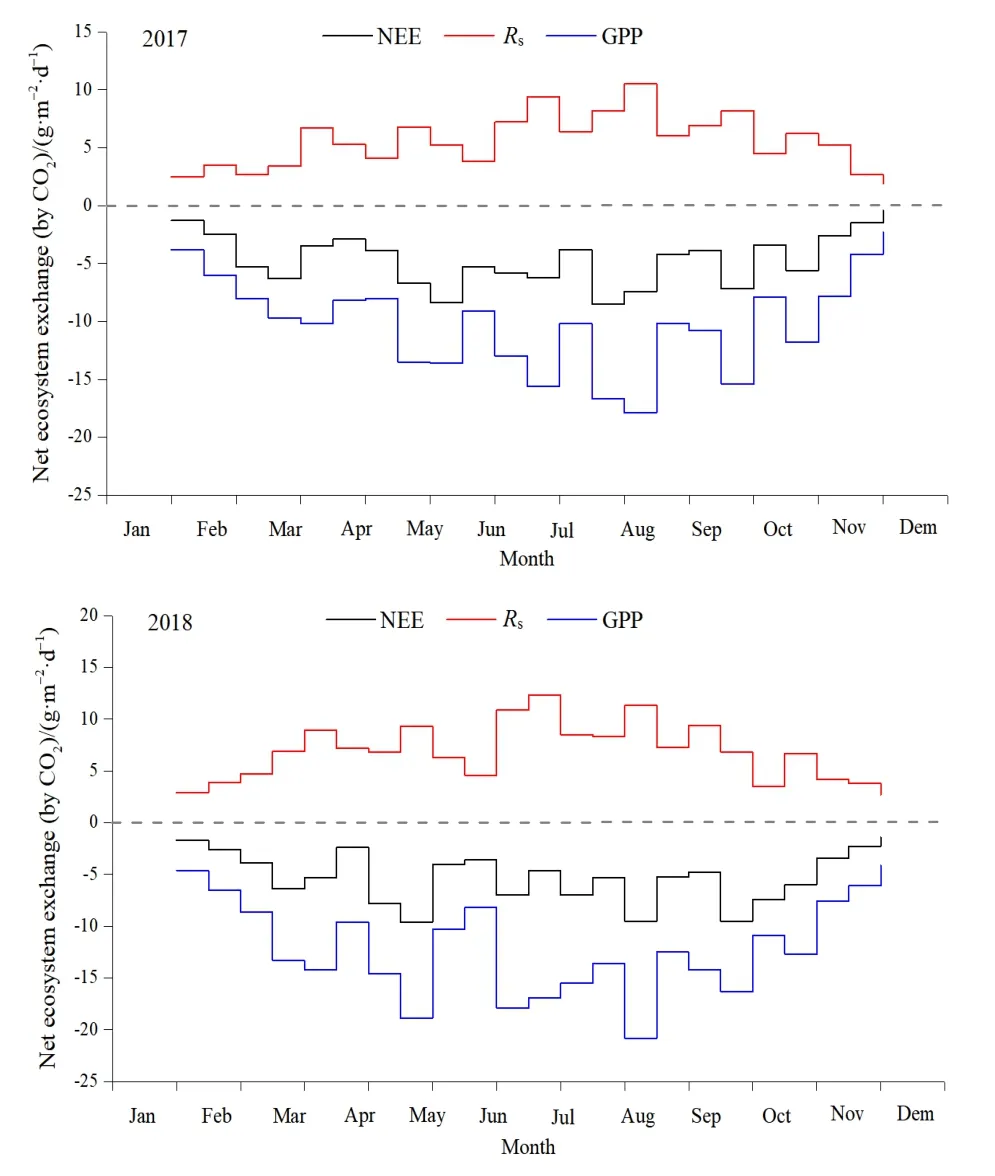
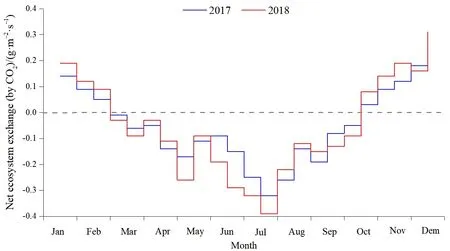
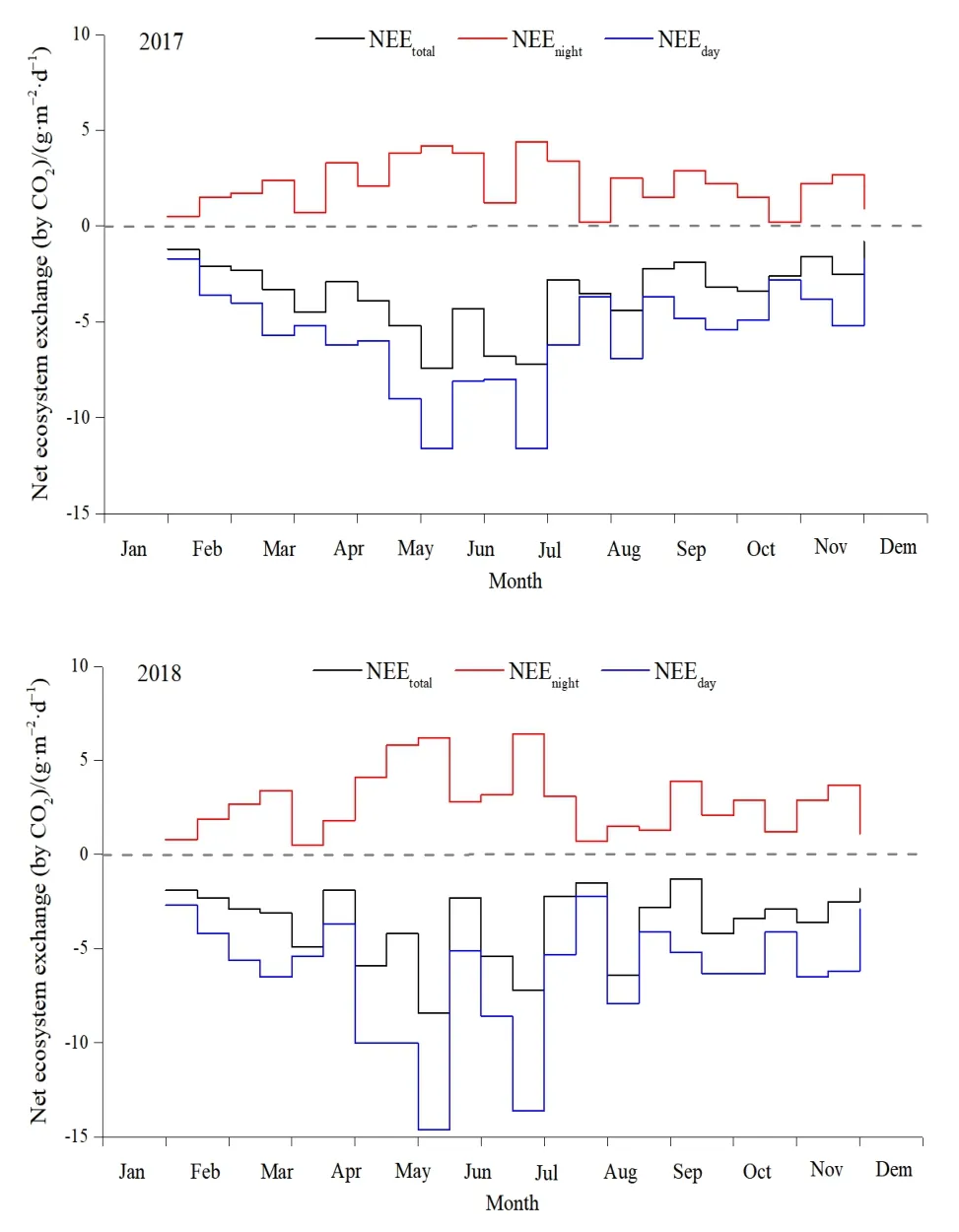
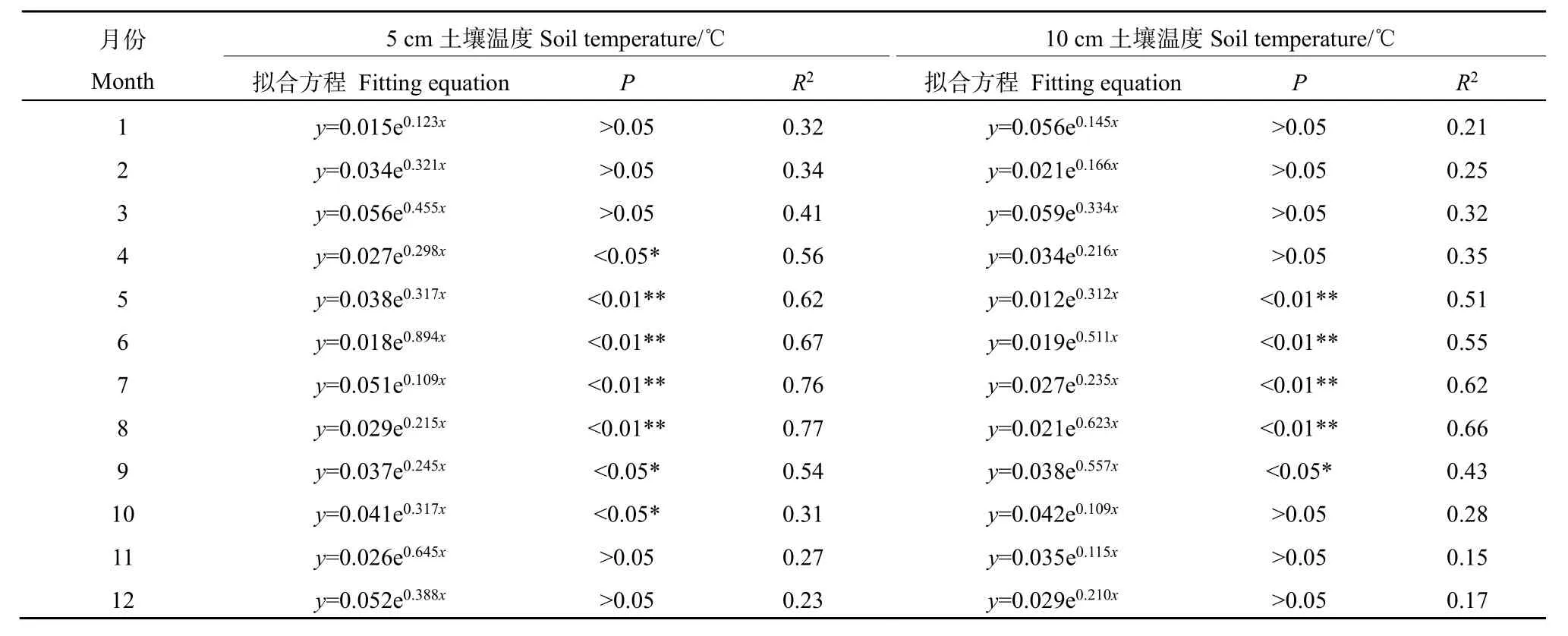
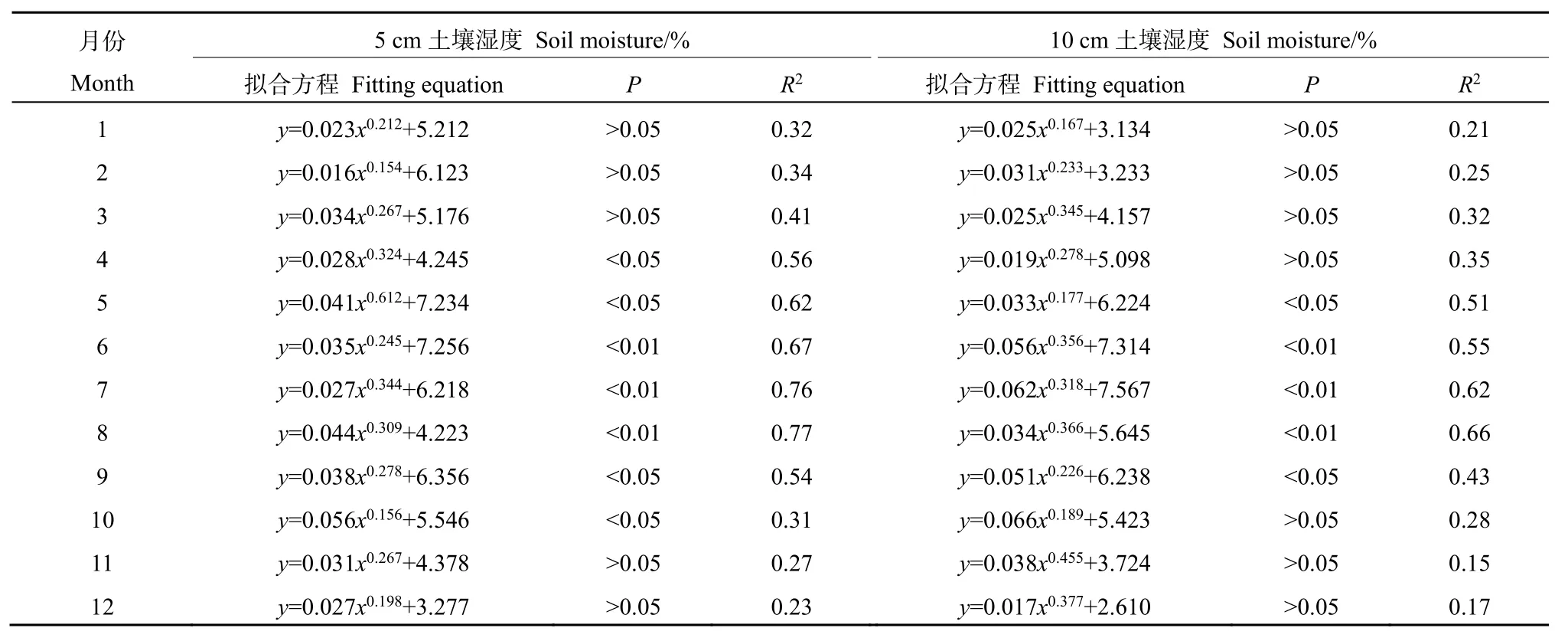
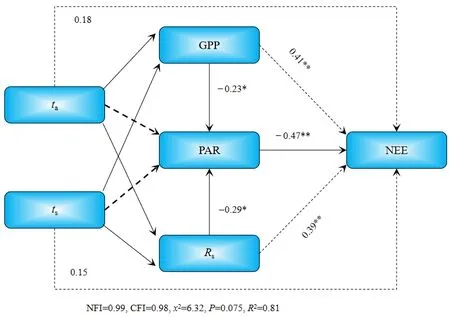
对插补后的数据划分白天(总辐射≥l W·m−2)和晚上(总辐射 通量塔直接观测到的CO2通量代表了净生态系统CO2交换(NEE),正值代表系统净排放CO2,负值代表系统净吸收CO2,是生态系统呼吸(Rs)和总初级生产力(GPP)之间的差值。因此,GPP可以定义为: GPP=NEE−Rs NEEtotal=NEEnight+NEEday 本研究利用路径分析的方法来评价 2017—2018年各个环境因子对CO2通量的影响。路径分析已经被广泛用于评价多个环境变量对碳通量季节和年际动态的相对重要性。这种方法是一种增强型的多元回归分析,能够用来评价各个环境变量之间的相互关系及对碳通量的直接和间接影响程度。本研究主要关注3个环境因子(空气温度ta、土壤温度ts和光合有效辐射PAR)对芦苇湿地CO2通量的影响。对于建立的模型,利用3个拟合度指数进行模型拟合度评估,若模型拟合度越高,则代表模型可用性越高,参数的估计越具有其涵义。3个拟合度指数分别为卡方统计量(x2)、基准化适合度指标(NFI,normed fit index)和比较适合度指标(CFI,comparative fit index)。其中,x2一般以卡方值P>0.05作为判断,意即模型具有良好的拟合度;NFI和CFI越接近于1表示模型拟合度越好。路径分析的软件是 AMOS 20.0(Analysis of Moment Structures)。 研究期间,环境因子表现出明显的季节变化特征。由图1可知,2017—2018年平均空气温度与降雨量基本呈一致的变化趋势,随着月份的增加呈倒“V”型,其中6月和7月有所降低;两年的平均空气温度最高值在5月(21.3 ℃)和8月(20.1 ℃),相对应降水量分别为86 mm和77 mm;最低气温出现在12月,为2.2 ℃;最低降水量也出现在12月,为9 mm。2017—2018年平均降水主要集中在4—8月,占全年总降水的50%以上。 图1 气温一年内动态变化特征Fig.1 The dynamic change characteristics of temperature in a year 由图2可知,2017年生态系统碳交换(NEE)变化范围在−0.4— −8.5 g·m−2·d−1(以CO2计,下同)之间,生态系统总初级生产力(GPP)变化范围在−2.3— −17.9 g·m−2·d−1之间,生态系统呼吸(Rs)变化范围在 1.9— −10.5 g·m−2·d−1之间。其中生态系统呼吸(Rs)随着月份的增加呈倒“V”型变化特征,在8月达到最高;生态系统碳交换(NEE)和生态系统总初级生产力(GPP)随着月份的增加呈“V”型变化特征,在8月达到最高。2018年不同月份生态系统碳交换(NEE)、生态系统总初级生产力(GPP)、生态系统呼吸(Rs)均高于2017年,局部有所差异,其变化趋势与 2017年总体保持一致;其中生态系统呼吸(Rs)随着月份的增加呈倒“V”型变化特征,在8月达到最高;生态系统碳交换(NEE)和生态系统总初级生产力(GPP)随着月份的增加呈“V”型变化特征,在8月达到最高。 图2 生态系统碳交换(NEE)、生态系统总初级生产力(GPP)、生态系统呼吸(Rs)Fig.2 Ecosystem carbon exchange (NEE),gross primary productivity (GPP),ecosystem respiration (Rs) 光合有效辐射(PAR)是控制光合作用的主要因素之一,因此PAR的大小也强烈影响NEE的大小。图3给出了2017—2018年净生态系统碳交换(NEE)的日动态,其中相同时间2018年净生态系统碳交换(NEE)基本高于 2017年。日出后(大约07:00)生态系统开始吸收CO2,随着PAR的增加,光合作用逐渐增强,NEE逐渐由净排放(正值)转为净吸收(负值),固碳速率逐渐增大,大约在10:00达到CO2吸收峰值。但随后随着PAR的继续增加,系统固碳速率开始逐渐降低。14:00左右生态系统固碳能力降低,出现“午休”现象。到午后15:00左右,生态系统固碳能力又开始增强,达到第2个CO2吸收高峰,19:00左右NEE接近于0,生态系统开始向大气中排放 CO2。通过对连续2 a的观测数据分析发现,CO2的最大吸收速率为0.32(2017 年)和 0.35(2018 年)g·m−2·s−1。 图3 2017—2018年净生态系统碳交换日动态Fig.3 Net ecosystem carbon exchange day dynamics from 2017 to 2018 由图4可知,2017年NEEtotal变化范围在−0.4— −8.5 g·m−2·d−1之间,NEEnight变化范围在−2.3—−17.9 g·m−2·d−1之间,NEEday变化范围在 1.9—10.5 g·m−2·d−1之间。其中 NEEnight随着月份的增加呈倒“V”型变化特征,在8月达到最高;NEEtotal和NEEday随着月份的增加呈“V”型变化特征,在8月达到最高。2018年不同月份NEEnight、NEEtotal和NEEday均高于2017年,局部有所差异,其变化趋势与2017年总体保持一致;其中 NEEnight随着月份的增加呈倒“V”型变化特征,在8月达到最高;NEEtotal和NEEday随着月份的增加呈“V”型变化特征,在 8月达到最高。 图4 2017—2018年净生态系统碳交换季节动态特征Fig.4 Seasonal dynamics of carbon exchange in net ecosystems from 2017 to 2018 利用 NEE数据与不同深度土壤温度和土壤湿度进行回归分析,筛选出最优模式;与土壤温度的回归分析表明(表1),4—10月NEE通量与5 cm土壤温度呈显著或极显著的指数关系(P<0.05,P<0.01),而5—9月NEE通量与10 cm土壤温度呈显著或极显著的指数关系(P<0.05,P<0.01);同时NEE通量与5 cm土壤温度的R2均高于NEE通量与10 cm土壤温度的R2,由此说明NEE通量与5 cm土壤温度的回归关系更好;NEE通量与土壤湿度的回归分析表明(表2),4—10月NEE通量与 5 cm土壤湿度呈显著或极显著的幂数关系(P<0.05,P<0.01),而5—9月NEE通量与10 cm土壤湿度呈显著或极显著的幂数关系(P<0.05,P<0.01);同时 NEE通量与5 cm土壤湿度的 R2均高于NEE通量与10 cm土壤湿度的R2,由此说明NEE通量与5 cm土壤湿度的回归关系更好。综合分析表明5 cm土壤温度和湿度能够更好地指示NEE通量的变化。 表1 土壤温度与NEE通量的指数关系Table 1 Exponential relationship between soil temperature and NEE flux 表2 土壤湿度与NEE通量的指数关系Table 2 Exponential relationship between soil moisture and NEE flux 在黄河三角洲芦苇湿地生态系统,多个环境因子会影响 NEE通量。利用路径分析的方法探讨各个环境因子对芦苇湿地碳收支的直接影响和间接影响(图5)。由图可知,模型的拟合度较好,NFI和CFI均高于0.9,且P>0.05,模型的可用性较高。其中ta和ts对NEE的路径系数分别为0.18和0.15,而GPP和Rs对NEE的路径系数分别为0.41和0.39,表明GPP和Rs直接影响黄河三角洲芦苇湿地NEE通量,而ta和ts对NEE通量起到间接的影响作用;此外,光合有效辐射与NEE呈显著的负相关关系,即随着PAR的增强,芦苇湿地生态系统的固碳能力在逐渐增强。 图5 路径分析图Fig.5 The path analysis diagram 本研究中,生长季芦苇湿地各月 NEE的日均值呈现“U”形变化曲线,这与以往的研究结果一致(Fang et al.,2018;Zhang et al.,2016)。湿地净生态系统CO2交换的年际变化可以反映特定生态系统的碳源/汇功效。由于生态系统覆盖的植被及气候环境的不同,各湿地生态系统的碳收支状况往往表现不一(Rankin et al.,2018)。在本研究中,2017年生态系统碳交换(NEE)变化范围在−0.4—8.5 g·m−2·d−1之间,生态系统总初级生产力(GPP)变化范围在−2.3—17.9 g·m−2·d−1之间,生态系统呼吸(Rs)变化范围在 CO21.9—10.5 g·m−2·d−1之间。其中生态系统呼吸(Rs)随着月份的增加呈倒“V”型变化特征,在 8月达到最高;生态系统碳交换(NEE)和生态系统总初级生产力(GPP)随着月份的增加呈“V”型变化特征,在8月达到最高。2018年不同月份生态系统碳交换(NEE)、生态系统总初级生产力(GPP)、生态系统呼吸(Rs)均高于2017年,局部有所差异,其变化趋势与2017年总体保持一致;黄河三角洲芦苇湿地和国外及国内芦苇湿地比较,其生态系统碳通量的大体特征一致,均为碳汇,然而其日动态值低于若尔盖高寒湿地和辽河三角洲芦苇湿地,故三角洲芦苇湿地碳汇功能弱于其它同类型的湿地系统,说明黄河三角洲是一个净固碳量相对较高的地区(Zhou et al.,2015;Fleischer et al.,2016)。与此同时,特定的生态系统可能由于降水格局等的改变而表现出碳源/汇的不确定性。 PAR是影响植物光合作用的主要限制因子。当太阳辐射强度较低时,净生态系统 CO2交换随着PAR的增加而增加,其变化趋势符合双曲线方程(Fleischer et al.,2016)。有研究表明,生态系统呼吸速率(Rs)随着ts的升高而增加。本研究中,4—10月NEE通量与5 cm土壤温度呈显著或极显著的指数关系(P<0.05,P<0.01),而5—9月NEE通量与 10 cm土壤温度呈显著或极显著的指数关系(P<0.05,P<0.01);同时NEE通量与5 cm土壤温度的R2均高于NEE通量与10 cm土壤温度的R2,由此说明NEE通量与5 cm土壤温度的回归关系更好;NEE通量与土壤湿度的回归分析表明(表2),4—10月NEE通量与5 cm土壤湿度呈显著或极显著的幂数关系(P<0.05,P<0.01),而5—9月NEE通量与10 cm土壤湿度呈显著或极显著的幂数关系(P<0.05,P<0.01);同时 NEE通量与5 cm土壤湿度的R2均高于NEE通量与10 cm土壤湿度的R2,由此说明NEE通量与5 cm土壤湿度的回归关系更好,水分条件对NEE的影响仅次于温度,这与前人的研究相一致(Zhou et al.,2015;Fleischer et al.,2016)。相对于其他陆地生态系统,湿地通常不会因土壤水分过低而影响土壤呼吸,反而会由于土壤水分过高,导致呼吸产物CO2的排放受到限制。Zhou et al.(2015)研究认为,在7、8月,土壤水分饱和导致NEE主要来自植被呼吸,从而影响NEE与ts的关系。由此说明黄河三角洲湿地生态系统呼吸对温度变化的敏感性相对较低,温度与水分对生态系统 CO2通量起着重要作用,而5 cm土壤温度和湿度能够更好的指示NEE通量的变化。 此外,黄河三角洲芦苇湿地夏季由于高温、强光、低湿等环境条件引起部分气孔关闭,或光合作用被抑制,在14:00左右出现“午休”现象,时间晚于草原生态系统(例如内蒙古羊草草原 08:00—10:00,青海湖草甸草原11:30—13:00) (Sharifi et al.,2016;Kelsey et al.,2016)。植物光合作用过程中这种午间降低现象已被许多研究证实,是较为普遍的现象。而黄河三角洲芦苇湿地,由于其外在气候、地理环境条件及自身生理条件与草原生态系统的差异,导致其“午休”现象出现时间的推迟。从季节动态来看,由路径分析发现土壤温度是黄河三角洲芦苇湿地CO2通量变化的主要影响因子,而降水量和PAR对CO2通量的变化影响次之,这若尔盖高原高寒湿地的研究结论基本一致,认为PAR、温度和降水显著影响湿地生态系统的CO2通量。其它湿地的研究也发现了温度在控制碳平衡中的重要性。而在高海拔或者高寒地区,昼夜温差以及温度的季节变化非常大,因此也成为控制CO2通量的重要环境变量。然而,这些研究缺乏各个环境因子对CO2通量相对重要性和贡献量的比较研究。本研究的路径分析则提供了一些新的信息,明确反映了温度及其它环境因子对湿地碳通量的直接影响、间接影响及影响程度。 季节尺度上,芦苇湿地生长季具有明显的碳汇功能,生态系统呼吸(Rs)随着月份的增加呈倒“V”型变化特征,在 8月达到最高;生态系统碳交换(NEE)和生态系统总初级生产力(GPP)随着月份的增加呈“V”型变化特征,在8月达到最高。2018年不同月份生态系统碳交换(NEE)、生态系统总初级生产力(GPP)、生态系统呼吸(Rs)均高于2017年,局部有所差异,其变化趋势与2017年总体保持一致。在日尺度上,2017—2018年芦苇湿地NEE日变化特征表现为两个CO2吸收高峰,分别出现在11:00和16:00左右,其特点是在午间出现了碳交换通量的降低,CO2排放的日最大值两个生长季均出现在8月。 2017年NEEnight随着月份的增加呈倒“V”型变化特征,在8月达到最高;而NEEtotal和NEEday随着月份的增加呈“V”型变化特征,在8月达到最高;2018年不同月份NEEnight、NEEtotal和NEEday均高于2017年,局部有所差异,其变化趋势与2017年总体保持一致。 回归分析显示生态系统的CO2交换受到光合有效辐射(PAR)、土壤温度(ts)和土壤体积含水量(Ta)的共同影响,生长季NEE通量与5 cm土壤温度和土壤湿度呈显著或极显著的指数关系(P<0.05,P<0.01),同时生长季NEE通量与5 cm土壤温度和土壤湿度的R2均高于NEE通量与10 cm土壤温度和土壤湿度的R2,由此说明5 cm土壤温度和湿度能够更好的指示NEE通量的变化。 从季节动态来看,路径分析发现土壤温度是黄河三角洲芦苇湿地CO2通量变化的主要影响因子,而降水量和PAR对CO2通量的变化影响次之,在高海拔或者高寒地区,昼夜温差以及温度的季节变化非常大,因此也成为控制CO2通量的重要环境变量。然而,这些研究缺乏各个环境因子对CO2通量相对重要性和贡献量的比较研究。1.3 路径分析
2 结果与分析
2.1 环境因子动态特征

2.2 生态系统碳交换(NEE)、生态系统总初级生产力(GPP)、生态系统呼吸(Rs)
2.3 净生态系统碳交换日变化特征
2.4 净生态系统碳交换季节动态特征
2.5 环境因子对NEE通量的影响
3 讨论
4 结论
