间作对咖啡和澳洲坚果根系形态、分布及养分累积的影响
2021-08-06董云萍朱华康赵青云孙燕林兴军张玲艳龙宇宙
董云萍 朱华康 赵青云 孙燕 林兴军 张玲艳 龙宇宙


摘 要:為探明澳洲坚果与咖啡间作对地上部生长及地下部互作效应,为生产推广复合高效栽培模式提供理论依据,布置了咖啡和澳洲坚果间作盆栽实验,以二者分别单作为对照,分析植株的生长量、生物量、养分含量、根系形态以及土壤养分的差异。结果表明:(1)间作咖啡和间作澳洲坚果根干重、茎干重、单株总干重分别为40.86、50.38、148.34 g,22.38、33.19、111.97 g,间作咖啡显著大于间作澳洲坚果,叶干重二者差异不显著;幼龄期间作条件下,咖啡生长速率快于澳洲坚果。间作咖啡、间作澳洲坚果与各自单作根、茎、叶、单株干物质累积量差异均不显著。(2)间作澳洲坚果叶氮累积量2135.27 mg显著大于间作咖啡的1957.60 mg,除此之外,间作咖啡根氮、磷、钾,茎氮、磷、钾,叶磷、钾,单株总氮、磷、钾累积量均显著大于间作澳洲坚果,表明幼龄期间作条件下,咖啡养分累积量占优势;间作澳洲坚果根氮、钾累积量742.80、103.22 mg显著大于单作的535.00、37.77 mg,可见,间作根系竞争促进澳洲坚果增加根部氮的分配,提高根钾的吸收累积。(3)间作咖啡和间作澳洲坚果根长、根表面积、根体积在本株侧面根系生态位分布量占总取样量的62.63%~65.67%,其次是在株间根系生态位分布量占总取样量的16.01%~27.53%,在对应本株近侧和对应本株远侧面2个根系生态位分布较少,可见间作后二者根系主要分布于自身根系生态位,在株间位置根系交叉重叠,未在土壤深度和广度空间形成差异分布,属于间作竞争型根系统。咖啡与澳洲坚果均属于浅根系,侧根直径小,成龄澳洲坚果是高大乔木,随着时间的推移,与咖啡间作将处于地上、地下部竞争优势。
关键词:咖啡;澳洲坚果;间作;根系形态;分布;养分累积
Abstract: In order to explore the effects on plant growth, root morphology, distribution and nutrient accumulation based on coffee-macadamia intercropping systems, an experiment was performed with three planting patterns: monoculture coffee (MC), monoculture macadamia (MM), and coffee-macadamia intercropping (IMC). The results showed that: (1) in IMC, the coffee leaves dry weight was no significantly compared with macadamia, but the root dry weight, stem dry weight and total dry weight per plant of coffee and Macadamia were as follows 40.86, 50.38, 148.34 g and 22.38, 33.19, 111.97 g. It showed those of coffee were significantly higher than those of Macadamia. the young coffee plant growth rate was faster than that of young Macadamia plant. The dry weight of coffee root, stem, leaves and total per plant dry weight were not significantly different in MC and in IMC, and so as the macadamia. (2) in IMC, the N accumulation of macadamia leaves was 2135.27 mg, significantly higher than that of coffee (1957.60 mg). Beyond that, the N,P and K accumulation of stem and root; the P and K accumulation of leaf; total N, P and K accumulation per plant for coffee were significantly higher than those of macadamia. As a result, the young coffee plant had more nutrient accumulation compared with macadamia. The N and K accumulation of macadamia root was 742.80, 103.22 mg in IMC, higher than that of 535.00, 37.77 mg in MM, indicating that the macadamia N allocation to root increased due to the root competition between the two, and root K accumulation increased. (3) The root length, root surface area and root volume near the side of coffee and macadamia root distribution position was accounted for 62.63%~65.67% of the total sampled, The second root distribution position was between the two plants, accounted for 16.01%~27.53% of the total sampled. The least was near the side of the correspond plant and beyond the side of the correspond plant. So more root of coffee and macadamia was observed atthere-self root niche, and the root was overlapping between the two plant root space, the spatial distribution of the two plants root system was not separated in soil depth and width, showing that the two crops intercropping had a competition root system. Coffee and macadamia are shallow rooted plants with, thin root diameter. Macadamia is a large tree, so it will be in dominant position in field intercropping.
Keywords: Coffea Arabica L.; Macadamia ternifolia F. Muell; intercropping; root morphology; distribution; nutrient accumulation
澳洲坚果(Macadamia spp.),俗称夏威夷果,是山龙眼科(Proteaceae)澳洲坚果属(Macadamia F. Muell)的常绿果树,喜光照充足、温凉、湿润的热带亚热带气候条件。咖啡为茜草科(Rubiaceae)咖啡属(Coffea arabica L.)常绿灌木或小乔木,适宜半阴性、温凉、湿润的环境。这2种作物是云南省热区农民脱贫致富的支柱产业,特别是澳洲坚果,由于其经济价值高,产业发展迅速。截至2017年底,云南省澳洲坚果和咖啡种植面积分别为17.5万hm2、12.5万hm2,产值分别为8.22亿美元、3.16亿美元[1-2]。适宜种植区为临沧、德宏、普洱、保山等地。近年来,咖啡收购价格持续低迷,为了提高种植咖啡的经济效益和解决澳洲坚果非生产期长的产业问题,云南省各级政府积极鼓励发展咖啡与澳洲坚果间作,并得到了种植户和企业的积极响应,目前澳洲坚果、咖啡间作面积已超过3.3万hm2,但二者间作种间互作效应方面的研究鲜见报道。
间作群體中,作物之间存在地上部和地下部相互作用,从地上部来看,主要是相互之间对光照的竞争作用,具体表现为株高对相邻作物的影响。从地下部来看,主要是对土壤养分和水分的竞争。研究表明,地下部根系间的相互作用在间作产量优势中起了很重要的作用,根长密度和根表面积对主要借助扩散到达根表面的养分有效性有着决定性作用[3-7]。在发生竞争时,有的植物调节向根系分配的氮量,使根系变得细长,增加根系与土壤接触面积,进而增加竞争能力[7],有的植物会庞大自己的根系,占据更多的土壤空间[6],如小麦/玉米、大豆/小麦间作中小麦对其他物种竞争作用强烈,蚕豆/玉米间作中蚕豆对玉米呈现较弱的竞争甚至促进作用[8-9],槟榔/咖啡间作,咖啡根系延伸到槟榔根系生态位,能使该生态位槟榔根长密度增加[10]。合理间作可有效缓解作物之间的竞争,Du等[11]研究表明玉米大豆间作系统种间氮竞争通过减少氮投入促进大豆生物固氮,与相应的单作相比促进了氮素肥料的利用。养分利用能力不同的作物之间所表现出的互馈效应可以使土壤资源利用率得到提高[10,12-14],玉米/大豆间作土壤总氮、有效磷和土壤有机质含量均显著高于相应的单作作物[14]。
本研究拟对澳洲坚果、咖啡间作地上部及地下部互作效应进行初步的研究,为生产合理间作配置和有效管理间作作物提供理论依据。
1 材料与方法
1.1 材料
于2017年1月中旬从农业农村部瑞丽咖啡种质资源圃采摘小粒种咖啡‘德热155(Coffea arabica L.cv. DTARI No.155),鲜果经脱皮、脱胶、晾干,3月20日在中国热带农业科学院香料饮料研究所(简称“香饮所”)进行沙床催芽;澳洲坚果于2016年11月中旬从云南省热带作物研究所试验基地采收充分成熟的果实,品种为M. ternifolia F. Muell. cv.H2,剥去外果皮后,用干净的干河沙贮藏,于2017年2月13日在香饮所进行沙床催芽。4月30日分别移栽到12 cm×16 cm营养钵中培育2个月,选取长势一致的咖啡和澳洲坚果苗备用。
1.2 方法
1.2.1 试验设计 营养土配比为体积比表土∶ 牛粪∶沙=8∶1∶1,挖取香饮所防风林下0~ 20 cm表土,按比例加入充分堆沤腐熟的牛粪肥和粗河沙,混合均匀,按四分法取样并测定盆栽营养土养分含量,有机质22.5 g/kg、碱解氮142.33 mg/kg、速效磷34.15 mg/kg、速效钾188.76 mg/kg、pH 5.51。
盆栽试验于2017年7月1日布置,采用长×宽×高为48 cm×32 cm×26 cm长方形塑料花盆,每盆装配制好的营养土25 kg。设3个处理:咖啡单作,Monoculture coffee(MC),每盆种2株咖啡;澳洲坚果单作,Monoculture macadamia(MM),每盆种2株澳洲坚果;澳洲坚果间作咖啡,Coffee-Macadamia Intercropping(IMC),每盆种1株澳洲坚果和1株咖啡。随机区组设计,每个处理6盆为1个小区,每个区组包括3个小区,排成1列,共3列。在香饮所遮阳网大棚内(荫蔽度60%)培养,日常管理包括淋水,拔杂草,分别于培养3、6、10个月时施浓度为0.5%的市售复合肥(N∶P2O5∶K2O=15∶15∶15),每次每盆施2 L。培育12个月采样分析。
1.2.2 测定项目与方法 (1)植株生长量、氮平衡指数和叶绿素测定。每个小区选取长势一致的植株3株。株高,用钢卷尺测量从盆土表面处到植株顶端的距离;茎粗,用游标卡尺测量植株茎干距离土面5 cm处直径;氮平衡指数和叶绿素,在晴天上午的8:00—10:00进行,选取咖啡和澳洲坚果植株东、南、西、北4个方位顶芽往下数第3片成熟叶片,用植物氮平衡指数测量仪(法国F0RCE-A公司生产,型号SCIENTIFIC+)测定氮平衡指数和叶绿素相对值。
(2)植株生物量测定及植株样、土样采集。每个处理每小区收获2盆,共6盆,将盆连同植株放平,敲击盆外壁,使土与盆壁产生空隙,手握植株基部往外拉将整株根部取出,小心抖动,粗略去除根部泥土,用自来水洗净根部,带回实验室再用纯水冲洗根部,注意收集清洗过程中断的根系。分成叶片、茎、根三部分,用吸水纸擦干水分。分别称量各部分鲜重,然后置烘箱中105 ℃杀青30 min,75 ℃烘至恒重,称量各部分干重。将各处理植株干样用植物样品粉碎机粉碎,装入自封袋中,做好标记,用于植物养分分析。取完根后相同处理的盆栽土经充分混匀后按四分法取样装在自封袋中,做好标记,带回实验室风干,过筛后分装于样品袋中并做好标记。用于土壤养分分析。
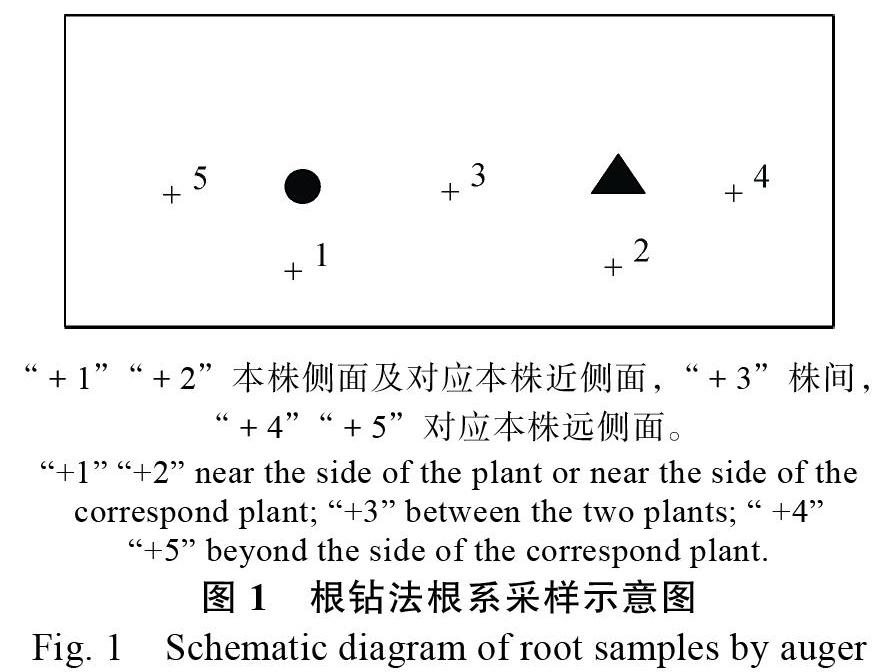
(3)根系取样。从每处理每小区长势一致的植株中选取2盆,以取样植株为中心,取样位置由近到远分成4个取样点,分别为本株侧面(near the side of the plant,SP)、株间(between the two plants,BP)、对应本株近侧面(near the side of the correspond plant,SCP)和对应本株远侧面(beyond the side of the correspond plant,BSCP)(图1)。钻头直径3.3 cm,长13 cm,每钻土约为94.03 cm3。取出的带土根样装入塑料自封袋,做好标签,带回实验室放入?20 ℃冰箱暂存。检测时取出放室温解冻后用水洗法将根从土壤中分离出来,平铺于透明的有机玻璃根盒内,并注入3 mm左右深的水,置于Epson根系扫描仪上用透视扫描法扫描,间作处理的根样将澳洲坚果和咖啡根系分开扫描,WINRhizo软件分析根系各生长参数。
(4)植株养分含量测定。全氮用有机元素分析仪(意大利Euro Vector E EA3000)测定;全磷用H2SO4-H2O2消煮,钼锑抗比色法测定;全钾用H2SO4-H2O2消煮,火焰光度法测定[15]。
(5)土壤養分含量、pH测定。碱解氮用碱解扩散法测定;速效磷用HCl-H2SO4浸提分光光度法测定;速效钾用NH4OAC浸提,火焰光度法测定;有效Fe、Mn、Cu、Zn用0.1 mol/L盐酸浸提,原子吸收光谱法测定;交换性Ca、Mg用1 mol/L中性醋酸铵提取,原子吸收光谱法测定[15]。土壤pH测定利用玻璃电极酸度计测定,水土比为2.5∶1(V/ω),土壤悬液值即为土壤pH[15]。
1.3 数据处理
数据均采用SPSS软件(SPSS 19.0)进行ANOVA方差分析和多重比较(LSD,P<0.05)。
2 结果与分析
2.1 间作对咖啡和澳洲坚果生长的影响
由图2可见,间作咖啡株高、茎粗分别比咖啡单作增加5.5%、4.5%,差异不显著;间作澳洲坚果株高比单作增加5.5%,茎粗减少8.4%,差异不显著。可见,间作不影响咖啡和澳洲坚果株高和茎粗的生长。在间作和单作下澳洲坚果株高均显著高于咖啡,茎粗两者差异不显著。
2.2 间作对咖啡和澳洲坚果生物量的影响
由图3可见,间作咖啡根鲜重、干重,茎鲜重、干重,叶片鲜重、干重分别比单作增加了5.5%、18.0%,19.3%、22.5%,19.6%、27.4%,其中叶鲜重差异达显著水平;澳洲坚果根鲜重、干重比单作增加了49.3%和49.4%,而茎鲜重、干重,叶片鲜重、干重分别比单作减少了18.1%、1.0%和17.3%、15.1%,差异均未达到显著水平。咖啡单株总鲜重、干重间作大于单作,澳洲坚果总鲜重、干重间作小于单作。可见,咖啡和澳洲坚果间作后咖啡的生物量比单作有所增加;而澳图中数据为各处理的平均值±标准偏差,不同小写字母表示差异性分析达显著水平(P<0.05)。
单作咖啡根干物质积累量显著大于单作澳洲坚果,叶干物质积累量则是单作澳洲坚果显著大于咖啡,茎干物质积累量和单株总干物质积累量二者差异不显著;间作咖啡根干重、茎干重和单株总干物质积累量显著大于间作澳洲坚果,叶干重两者差异不显著。
2.3 间作对咖啡和澳洲坚果植株养分累积的影响
从图4可见,相比单作,间作显著促进咖啡叶氮、根氮和澳洲坚果根氮累积量,显著减少咖啡茎氮和澳洲坚果叶氮、茎氮积累量(图4C);间作咖啡叶片磷累积量显著大于单作,而咖啡根磷、澳洲坚果叶磷、茎磷显著低于单作(图4B);除澳洲坚果叶钾累积量间作与单作差异不显著外,咖啡叶钾、茎钾、根钾和澳洲坚果茎钾、根钾累积量均是间作高于单作(图4A);从单株养分累积量可见,间作显著促进咖啡植株对氮、钾的累积,显著促进澳洲坚果植株对钾的累积,但显著降低澳洲坚果植株对氮、磷的累积(图4D)。
单作和间作下,叶氮累积量澳洲坚果显著高于咖啡,而茎氮、根氮、叶磷、茎磷、根磷、叶钾、茎钾、根钾,单株总磷、总钾以及间作下单株总氮累积量均是咖啡显著高于澳洲坚果。
2.4 间作对咖啡和澳洲坚果氮平衡指数及叶绿素含量的影响
由图5可见,间作咖啡氮平衡指数和叶绿素相对值与单作相比差异不显著。间作澳洲坚果氮平衡指数和叶绿素相对值显著低于单作。可见间作对咖啡氮素利用效率影响小,但间作显著抑制了澳洲坚果氮素利用效率。单作和间作下,氮平衡指数澳洲坚果显著高于咖啡,单作下叶绿素相对值咖啡和澳洲坚果差异不显著,间作下咖啡叶绿素相对值显著高于澳洲坚果。
2.5 间作对咖啡和澳洲坚果根系形态的影响
由表1和表2可见,单作咖啡和单作澳洲坚果根长、根表面积、根体积在株间、本株侧面、对应本株近侧面、对应本株远侧面4个根系生态位均匀分布。间作咖啡和间作澳洲坚果根长、根表面积、根体积在本株侧面根系生态位显著大于其他根系生态位,其次为株间根系生态位,其中间作咖啡在本株侧面和株间根长、根表面积、根体积占取样总根长、总根表面积、总根体积的63.49%、63.61%、63.40%,16.01%、18.04%、20.38%。间作澳洲坚果为62.63%、64.36%、65.67%,27.53%、25.91%、24.55%;间作下株间根系生态位澳洲坚果根长、根表面积大于咖啡,图中数据为各处理的平均值±标准偏差,不同小写字母表示差异性分析达显著水平(P<0.05)。
而对应本株近侧面和对应本株远侧面咖啡根长、根表面积、根体积大于澳洲坚果,差异未达显著。由此可见,间作后咖啡和澳洲坚果根系主要分布在各自的根系生态位,部分根系主要在株间根系生态位交叉生长。间作咖啡对应本株近侧面和对应本株远侧面侧根直径显著小于株间和本株侧面根系直径,说明咖啡根系生长到更远处的侧根变得更细。澳洲坚果侧根直径比咖啡侧根直径小,说明澳洲坚果侧根更细。
2.6 间作对土壤养分含量的影响
由表3和表4可见,单作和间作下土壤有机质、碱解氮、速效磷、速效钾、交换性钙、交换性镁、有效铁、有效铜差异不显著。间作土壤中有效锌显著降低;有效锰显著增加。
2.7 间作对土壤pH的影响
由图6可见,咖啡间作澳洲坚果后,土壤pH比单作咖啡和单作澳洲坚果略有降低,差异不显著。
3 讨论
单作和间作下澳洲坚果株高显著高于咖啡,这与澳洲坚果为高大乔木的生长习性有关。间作咖啡根干重、茎干重和单株总干物质积累量显著大于间作澳洲坚果,可见在间作条件下,咖啡生长速率快于澳洲坚果。
间作产量优势是以作物养分吸收优势为物质基础的[16],咖啡和澳洲坚果养分累积量大小与其干物质累积量趋势一致。作物对三大矿质养分(N、P、K)的吸收依靠扩散和集体流动过程的共同作用[17]。由于1年龄咖啡生长速率快于澳洲坚果,在间作系统中地下部生长处于优势,有利于吸收以质流运输为主的土壤氮源[18-19],因而其根、茎干物质和根氮、茎氮养分累积量显著大于澳洲坚果,间作咖啡叶氮、根氮累积量也显著大于单作咖啡。澳洲坚果植株较高,占据着地上部光热资源优势,因此澳洲坚果叶氮累积量显著大于咖啡。在间作下,虽然澳洲坚果地下部生长处于劣势,但为了与咖啡争夺氮源,加大了根部氮的分配,减少了叶片氮的分配,因而根氮累积量显著大于单作,而叶氮累积量显著小于单作。从间作澳洲坚果氮平衡指数和叶绿素含量显著小于单作也可说明间作使澳洲坚果减少了叶片氮的分配。叶磷、茎磷、根磷,叶钾、茎钾、根钾的累积量均是咖啡占显著优势。
根系分析表明,间作咖啡和间作澳洲坚果根长、根表面积、根体积在本株侧面根系生态位显著大于其他根系生态位,说明间作后二者根系主要分布在各自的根系生态位。二者根长、根表面积、根体积在取样的4个根系生态位差异不显著,在株间位置根系交叉重叠,未在土壤深度和广度空间形成差异分布,说明二者间作属于竞争型根系统。分析发现间作后咖啡根系生长到更远处的侧根变得更细,以增加根系与土壤接触面积,从而增加竞争能力,这与前人研究结果相一致[7]。Fitter等[20]研究得出,根系统的空间结构对土壤里的水分和礦物质吸收具有重要意义。澳洲坚果侧根比咖啡更细,从二者根长、根表面积在不同的根系生态位差异不显著,咖啡根干重显著大于澳洲坚果,可得出澳洲坚果相对根长和相对根表面积大于咖啡,也就是澳洲坚果根系与土壤接触面积会更大。磷和钾元素扩散迁移速度极慢,根系的广泛分布在磷、钾吸收上就显得尤为重要[18-19],这就有利于根系吸收大部分束缚在固相土壤里的钾和磷酸盐,这可能是间作显著促进澳洲坚果对钾的累积而减少咖啡根系对磷累积的原因之一。间作显著促进澳洲坚果和咖啡对钾的累积的原因也可能是根系分泌物活化钾,将土壤中的非交换性钾转变为能被作物吸收利用的有效钾[21-23],这有待后续进一步研究。
间作土壤有效锰显著增加,有效锌显著低降低,推测是间作降低了土壤的pH,使得Mn2+离子浓度增加。相比单作,间作促进了咖啡和澳洲坚果根对氮的累积,作物对氮、锌养分吸收存在协同效应[24],因而加大了二者对锌的吸收,使得间作土壤有效锌显著低于单作土壤。
在时间上作物种间对养分和水分的需求差异,也是影响根系对水分、养分吸收的重要原因[4],本盆栽试验通过分析得出在幼龄期咖啡生长速率快于澳洲坚果,在地下部干物质累积量和养分吸收累积方面占优势,但在成龄期,二者间作竞争关系可能发生改变。咖啡根系主要分布在0~ 30 cm土层[25],澳洲坚果根系主要分布在0~40 cm土层[26],由此可见,咖啡和澳洲坚果根系在土壤深度上将会形成长期的竞争关系[27]。成龄澳洲坚果是高大乔木,对地上部光热资源的利用占据优势,根系分布范围、相对根长和相对根表面积大于咖啡,这些有利因素将为大田成龄澳洲坚果带来间作竞争优势,而咖啡将处于竞争劣势。本研究仅仅是1年期的盆栽试验,咖啡和澳洲坚果间作长期的田间效应,根系形态及其相互作用等方面,有待进一步深入研究。
参考文献
杜 杉. 云南已成澳洲坚果种植老大, 第八届国际澳洲坚果大会将首次走进中国[J]. 时代金融, 2018 (19): 52-54.
刘艺卓, 焦 点. 自由贸易区建设对中国云南咖啡产业的影响[J]. 世界农业, 2017(3): 181-184.
安颖蔚, 冯良山, 张 鹏. 间作群体作物根系营养竞争与互作效应[J]. 江苏农业科学, 2017, 45(5): 26-28.
陈 伟,薛 立. 根系间的相互作用?竞争与互利[J]. 生态学报, 2004, 24(6): 1243-1251.
Schenk H J. Root competition: beyond resource depletion[J]. Journal of Ecology, 2006, 94(4): 725-739.
Craine J M. Competition for nutrients and optimal root allocation[J]. Plant and Soil, 2006, 285(1/2): 171-185.
Fransen B, Blijjenberg J, de Kroon H. Root morphological and physiological plasticity of perennial grass species and the exploitation of spatial and temporal heterogeneous nutrient patches[J]. Plant and Soil, 1999, 211(2): 179-189.
李 隆. 间套作体系豆科作物固氮生态学原理与应用[M]. 北京: 中国农业大学出版社, 2013.
Li L, Yang S C, Li X L, et al. Interspecific complementary and competitive interactions between intercropped maize and fababean[J]. Plant Soil, 1999, 212(2): 105-114.
王曉阳, 董云萍, 邢诒彰, 等. 单作和间作对槟榔和咖啡生长、根系形态及养分利用的影响[J]. 热带作物学报, 2018, 39(10): 1906-1912.
Du Q, Zhou L, Chen P, et al. Relay-intercropping soybean with maize maintains soil fertility and increases nitrogen recovery efficiency by reducing nitrogen input[J]. The Crop Journal, 2020, 8(1): 140-152.
余常兵, 孙建好, 李 隆. 种间相互作用对作物生长及养分吸收的影响[J]. 植物营养与肥料学报, 2009, 15(1): 1-8.
王 灿, 杨建峰, 祖 超, 等. 胡椒园间作槟榔对胡椒产量及养分利用的影响[J]. 热带作物学报, 2015, 36(7): 1191-1196.
Fu Z D, Zhou L, Chen P, et al. Effects of maize-soybean relay intercropping on crop nutrient uptake and soil bacterial community[J]. Journal of Integrative Agriculture, 2019, 18(9): 2006-2018.
鲍士旦. 土壤农化分析[M]. 北京: 中国农业出版社, 2000.
李 隆, 杨思存, 孙建好, 等. 春小麦大豆间作条件下作物养分吸收积累动态的研究[J]. 植物营养与肥料学报, 1999, 5(2): 68.
Casper B B, Jackson R B. Plant competition underground[J]. Annual Review of Ecology and Systematis, 1997, 28: 545-570.
Natarajan M, Willey R W. The effects of water—stress on yield advantages of intercropping systems[J]. Field Crops Research, 1986, 13(2): 117-131.
蔡昆争. 作物根系生理生态学[M]. 北京: 化学工业出版社, 2011.
Fitter A H, Stickland T R. Architectural analysis of plant root systems. II. Influence of nutrient supply on architecture in contrasting plant species[J]. New Phytologist, 1991, 118: 383-389.
付学鹏, 吴凤芝, 吴 瑕, 等. 间套作改善作物矿质营养的机理研究进展[J]. 植物营养与肥料学报, 2016, 22(2): 525-535.
Kraffczyk I, Trolldenier G, Beringer H. Soluble root exudates of maize: influence of potassium supply and Rhizosphere microorganisms[J]. Soil Biology and Biochemistry, 1984, 16(4): 315-322.
Li T X, Ma G R, Wang C Q, et al. Mineral potassium activation in rhizosphere soils and root exudates of grain amaranth[j]. Joural of Soil Science, 2003, 34(1): 48-51.
何忠俊, 华 珞, 白玲玉, 等. 土壤?植物系统中氮锌交互作用研究进展[J]. 土壤与环境, 2001, 10(2): 133-137.
董云萍, 龙宇宙, 孙 燕, 等. 咖啡高产栽培技术[M]. 北京: 中国农业出版社, 2009.
陈 菁, 陆超忠, 石伟琦, 等. 澳洲坚果根系分布特点[J]. 热带农业科学, 2017, 37(6): 23-26.
Rubio G, Walk T, Ge Z Y, et al. Root gravitropism and below-ground competition among neighbouring plants: a modelling approach[J]. Annals of Botany, 2001, 88: 929- 940.
责任编辑:沈德发
