重庆30份茶树种质资源农艺性状与生化成分多样性
2021-08-05翟秀明胡方洁侯渝嘉
翟秀明,李 解,唐 敏,胡方洁,张 军,侯渝嘉,*,徐 泽
(1.重庆市农业科学院 茶叶研究所,重庆 402160; 2.重庆市茶叶工程技术研究中心,重庆 402160)
重庆是茶树原产地之一,也是人工种植茶树最早的地区[1]。重庆地处中国西南部,属于亚热带季风性湿润气候,降水丰富,冬暖春早,多云雾,少霜雪,立体气候明显,具有发展茶产业的先天优越气候环境条件,是我国最古老的茶区之一,也是我国茶树生长最适宜地区之一。重庆茶树种质资源极为丰富,有上千年历史的野生大茶树,也有经过长期栽培、驯化形成的各色地方品种,为茶树品种的培育与改良提供了丰富的材料。目前,茶已成为我国的重要经济作物,也是南方丘陵山区农民脱贫致富的支柱产业之一[2]。茶树种质资源是品种创新、新品种培育的物质基础,对茶叶生产和科学研究具有十分重要的价值和意义。“第三次全国农作物种质资源调查与收集”重庆项目组的启动,对重庆茶树种质资源的保护、发掘和利用奠定了坚实的基础。
农艺性状是与作物品种特性息息相关的性状,是判别种质资源鉴定分类和多样性研究的重要依据之一,具有直接、简便、与育种目标联系紧密等优点,农艺性状比较是目前种质资源研究最基本的方法和途径[3-5]。生化成分是决定茶叶适制性和制茶品质的物质基础[6]。利用农艺性状和生化成分对茶树种质资源进行分类是一种结果稳定、客观真实的研究方法,在茶树种质资源鉴定分类中应用较为广泛[7-9]。本研究采用农艺性状与生化成分相结合的方法,对保存于重庆市农业科学院茶叶研究所资源圃内的30份茶树资源进行鉴定、评价,挖掘其育种潜力,为优质特异新品种的选育提供研究基础。
1 材料与方法
1.1 材料
研究材料来源于“第三次全国农作物种质资源调查与收集”行动从重庆江津区、武隆区、酉阳县、南川区等地收集的地方品种、野生种单株或变异单株,材料保存于重庆市农业科学院茶叶研究所资源圃。1~15 号、21~30 号材料为重庆地方品种或资源,16~20号材料为在重庆栽种25 a的福鼎大白茶与川茶群体种杂交后代单株选择资源。材料名称与收集地见表1。
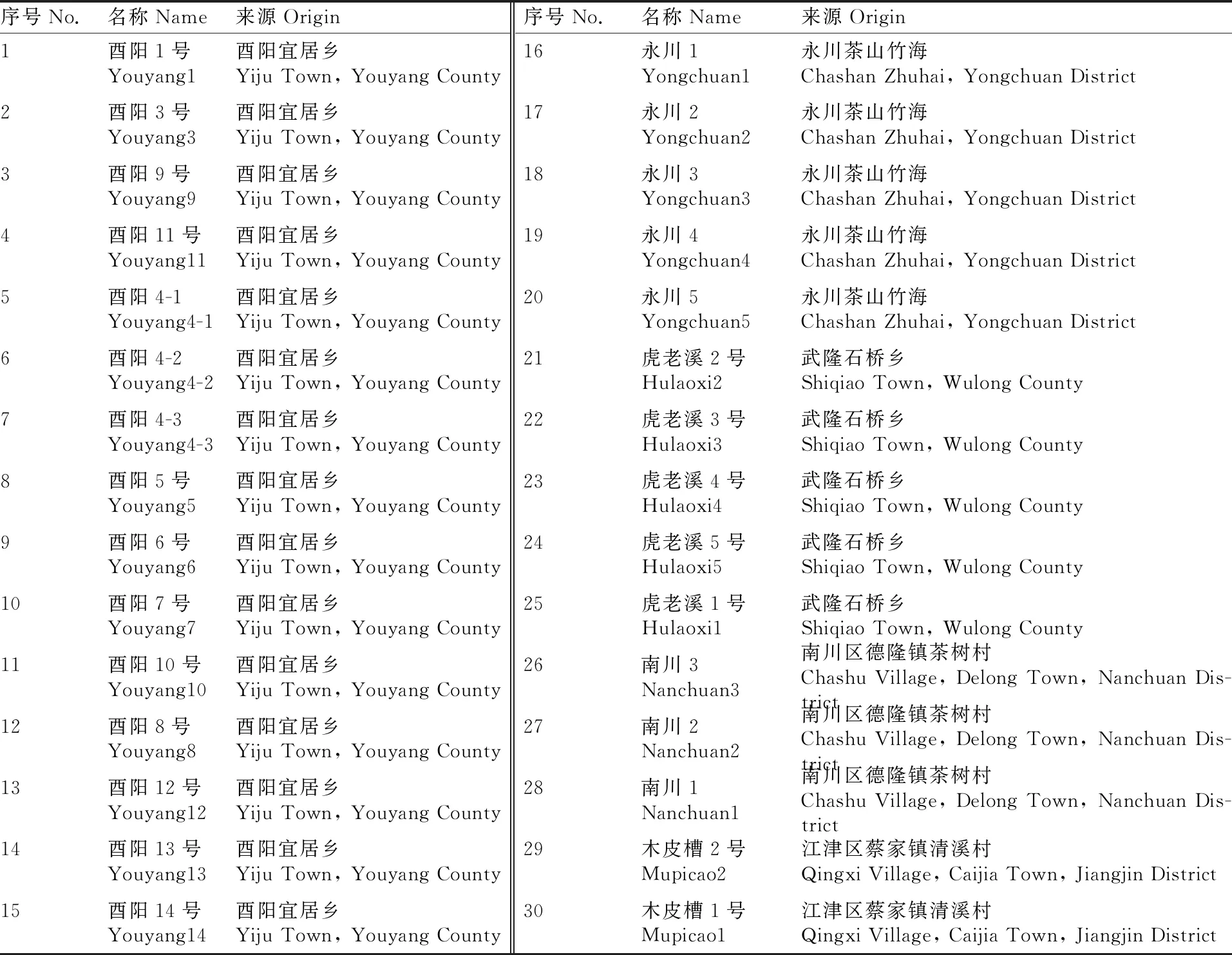
表1 30份重庆茶树资源基本情况
1.2 试验设计
2015—2016年将各地收集的茶树资源移栽至重庆市农业科学院茶叶研究所试验基地,并进行扦插扩繁。基地位于重庆市永川区茶山竹海,海拔460 m,29°75′N,105°71′E,年平均气温17.9 ℃,降雨量1 034.9 mm,属于亚热带季风性湿润气候。茶树材料按照双行双株种植,大行距1.5 m,小行距0.3 m,株距0.33 m,常规管理。
1.3 表型性状数据测定
于2018—2020年连续3 a对30份茶树材料的表型进行调查统计,包括18个描述型性状(树型、芽叶色泽、芽叶茸毛、一芽一叶期、一芽二叶期、叶片大小、叶形、叶色、叶尖、叶缘、叶面、叶身、花萼色泽、花瓣色泽、花瓣质地、子房茸毛、花柱裂位、雌雄蕊相对高度)和12个数值型性状[发芽密度、一芽二叶长、一芽二叶百芽重、叶长、叶宽、叶脉对数、萼片数、花冠直径(短)、花冠直径(长)、花瓣数、花柱长度、柱头开裂数]。性状调查参照《茶树种质资源描述规范和数据标准》进行,每份茶树材料的表型性状均重复测定10次,并进行年度重复。
1.4 主要生化成分测定
2018—2020年每年采摘第一轮一芽二叶茶鲜叶制作生化分析样,用于常规成分测定。参照GB/T 8305—2013的方法测定水浸出物含量;参照GB/T 8314—2013茚三酮比色法测定游离氨基酸总量;参照GB/T 8313—2008酒石酸铁比色法测定茶多酚含量;参照GB/T 8312—2013测定碱式乙酸铅比色法测定咖啡碱含量;可溶性糖含量采用蒽酮比色法测定。
1.5 数据处理
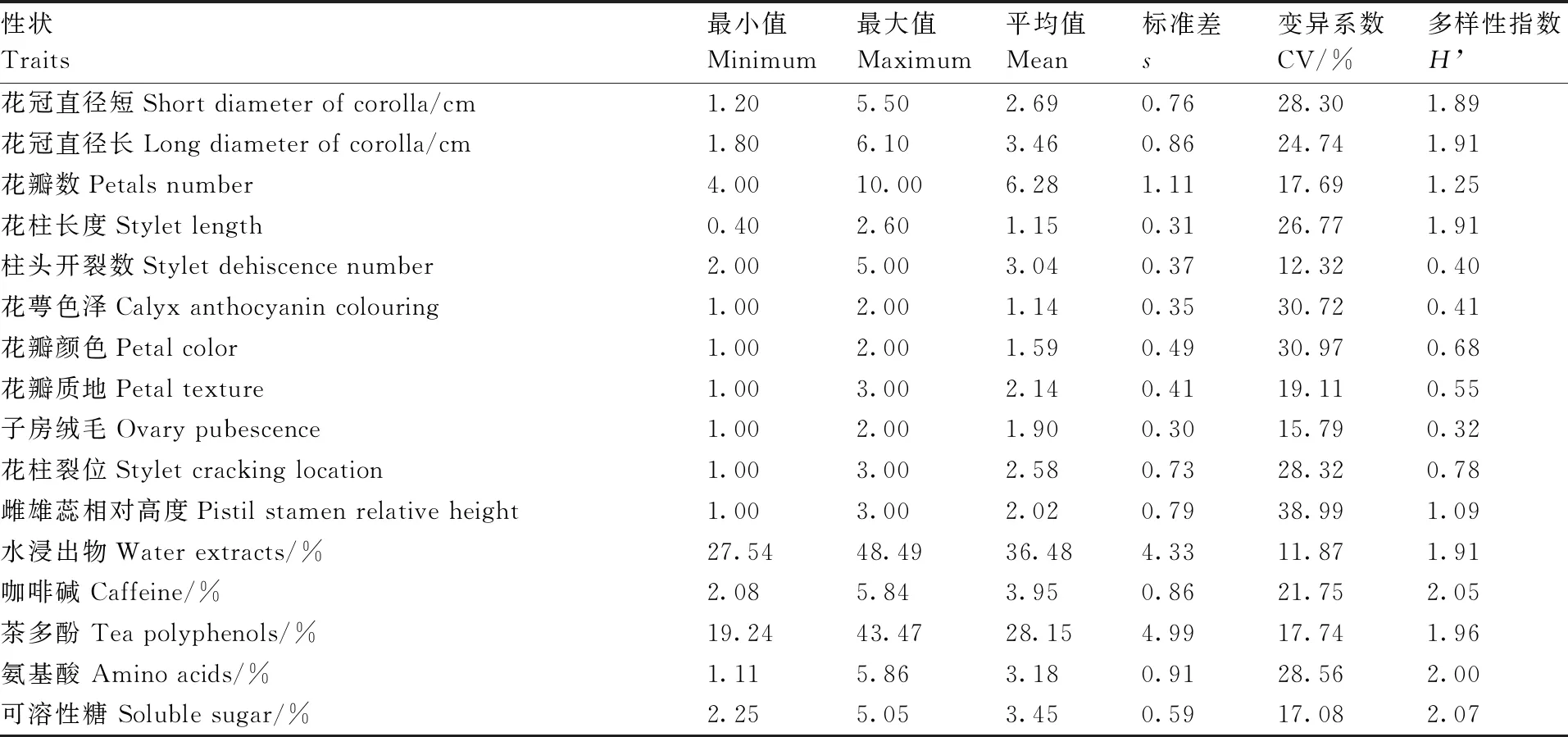
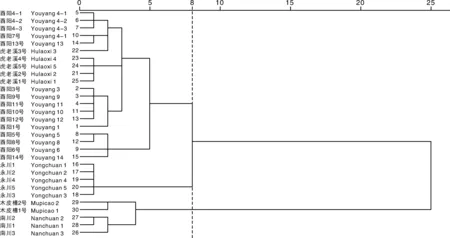
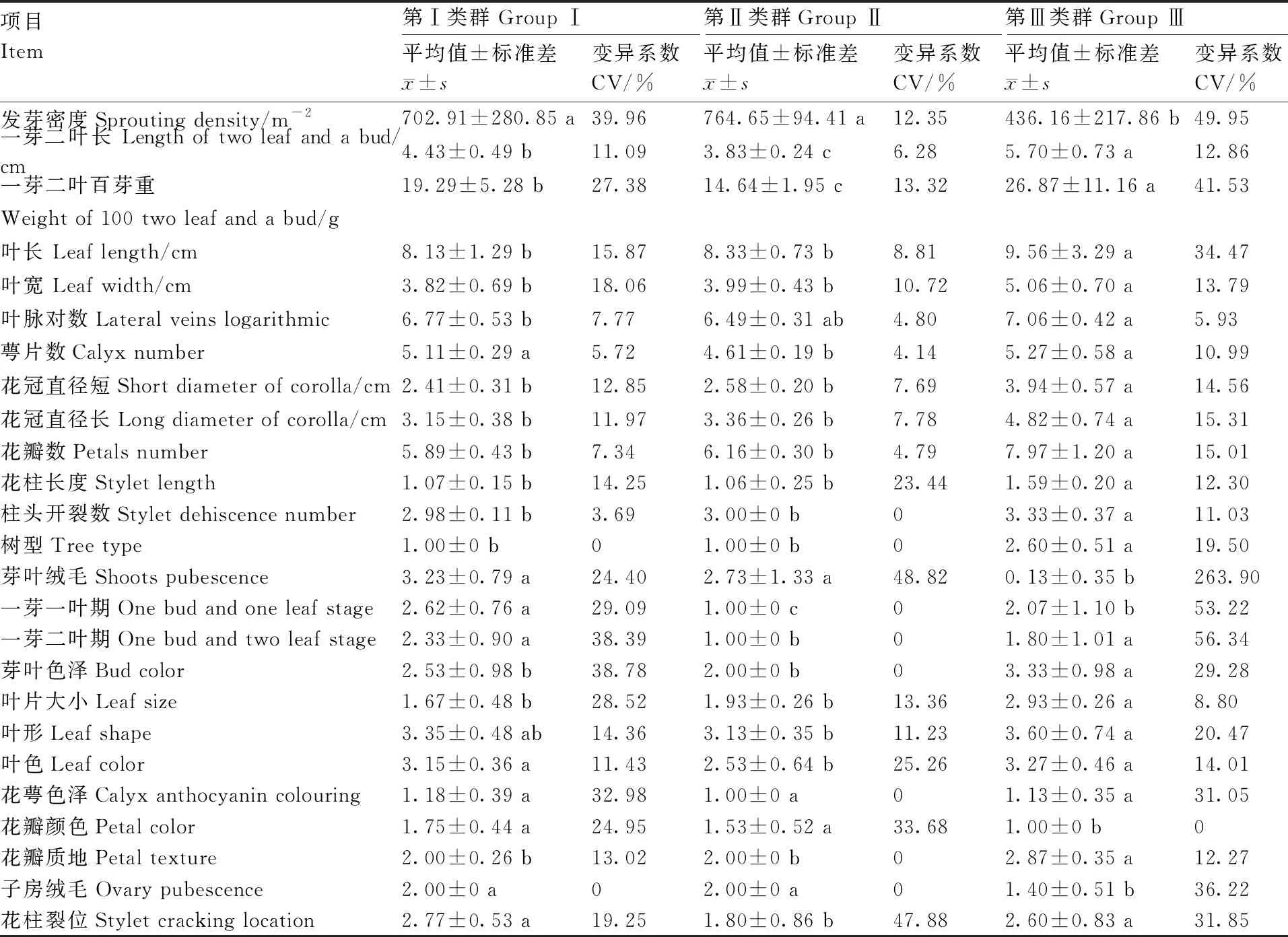
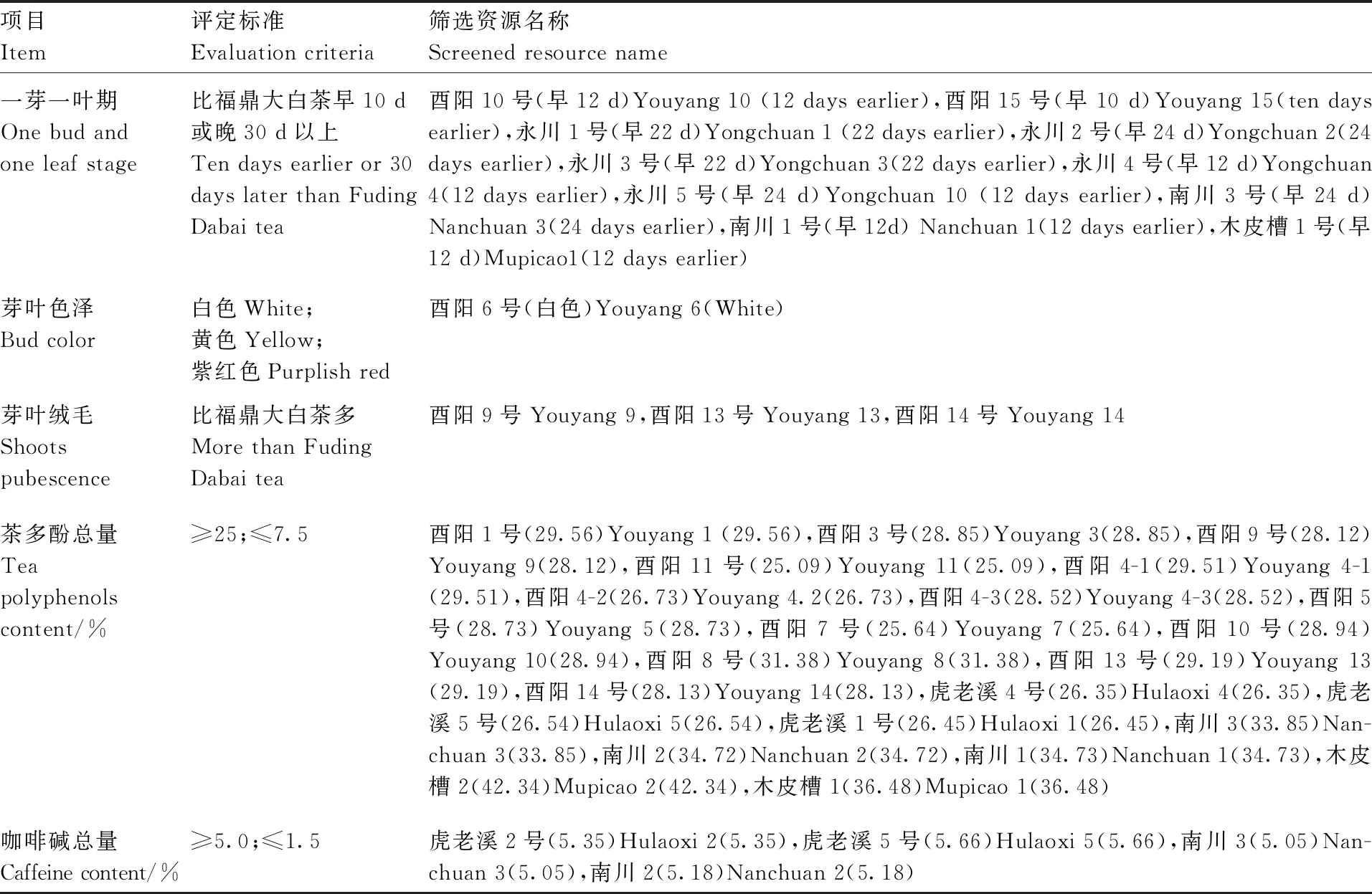
使用Microsoft Excel进行数据整理与预处理,描述型性状通过赋值法将其转化成数值型性状,采用IBM SPSS Statistics 22软件进行主成分分析、方差分析、聚类分析。聚类分析采用系统聚类中的离差平方和法,利用欧式平方距离计算种质间的遗传距离[10]。利用Shannon-Wiener指数H’进行农艺性状与品质性状的遗传多样性评价。计算H’时,先对数量性状进行质量化处理,以各性状极差的1/10为间距对每个性状进行10级分类,1级 表2 描述型性状指标与赋值标准 由表3可以看出,重庆茶树种质资源农艺性状变异系数范围较大,介于11.01%~53.54%,主要生化成分变异系数范围较小,介于11.87%~28.56%。农艺性状中芽叶绒毛的变异系数最大,为53.54%,然后是一芽二叶期、叶缘、叶面、一芽一叶期和发芽密度,变异系数分别为47.76%、46.06%、45.43%、42.61%、42.39%,均在40%以上;农艺性状中变异系数最小的为萼片数,为11.01%,柱头开裂数和叶脉对数的变异系数分别为12.32%和12.86%,均在15%以下。主要生化成分中,变异系数最大的为氨基酸(28.56%),最小的为水浸出物(11.87%),5个主要生化成分的变异程度依次为氨基酸>咖啡碱>茶多酚>可溶性糖>水浸出物。重庆茶树种质资源农艺性状的多样性指数差异较大,介于0.32~2.04;其中,叶宽、叶长的变异类型较为丰富,多样性指数分别为2.04和2.00;子房绒毛、柱头开裂数和花萼色泽的多样性指数较低,分别为0.32、0.40和0.41,多样性指数均在0.5以内。主要生化成分的变异类型较为丰富,多样性指数均在1.9以上,多样性指数最大的为可溶性糖(2.07),最小的为水浸出物(1.91)。综合各性状和生化成分的平均值、变异系数和多样性指数可以看出,重庆茶树种质资源在发芽密度、一芽一叶期、一芽二叶期、芽叶绒毛、氨基酸、咖啡碱含量等方面存在较为丰富的遗传变异类型,可为茶树品质改良提供丰富的材料基础。 表3 重庆茶树种质资源农艺性状与主要生化成分的变异统计和多样性分析 续表3 Continued Table 3 对重庆30份茶树资源的农艺性状和主要生化成分共计35个性状进行主成分分析,分析结果见表4。以特征值大于1为筛选标准,共提取出9个主成分,累计贡献率达74.85%,包含了35个性状指标的大部分信息,可以用来进行综合评价重庆茶树种质资源。贡献率第1的主成分特征值为11.638,贡献率为33.252%,主要反映了树型、花冠直径(短)、花冠直径(长)、花瓣数、花柱长度和花瓣质地等信息,为与茶树花特征相关的因子;贡献率第2的主成分特征值为3.574,贡献率为10.211%,主要解释了一芽一叶期、一芽二叶期等与茶树物候期相关的性状;贡献率第3的主成分特征值为2.038,贡献率为5.823%,其中,叶长的贡献率最大,其次为芽叶色泽、叶宽、叶面、叶身等与茶树芽叶性状相关的因子;贡献率第4的主成分特征值为2.004,贡献率为5.726%,主要反映了雌雄蕊相对高度、氨基酸含量等的影响;贡献率第5的主成分特征值为1.699,贡献率为4.853%,贡献率最大的仍然是雌雄蕊相对高度,其次为叶形、茶多酚、水浸出物等,其中茶多酚、水浸出物的影响为负;贡献率第6的主成分特征值为1.552,贡献率为4.433%,贡献率最大的为可溶性糖含量,其次为叶色、一芽一叶期、一芽二叶期和氨基酸含量,其中,叶色的影响为负;贡献率第7的主成分特征值为1.366,贡献率为3.902%,贡献率较大的因子有叶缘、花萼色泽、叶尖、可溶性糖等,叶缘影响为负;贡献率第8的主成分特征值为1.278,贡献率为3.653%,主要反映了花柱裂位、柱头开裂数、一芽二叶百芽重、叶色、一芽二叶长等的影响,其中花柱裂位、柱头开裂数贡献率为负;贡献率第9的主成分特征值为1.049,贡献率为2.997%,贡献率较大的因子有花柱裂位、叶长、氨基酸、咖啡碱等。 表4 农艺性状和主要生化成分的主成分分析 结合30个农艺性状和5个主要生化成分,在欧式平方距离8处,将30份重庆茶树种质资源分为3个类群,见图1。第Ⅰ类群包含2个亚群,共20份种质资源,第1个亚群包括酉阳4-1、酉阳4-2、酉阳4-3、酉阳7号、酉阳13号、虎老溪3号、虎老溪4号、虎老溪5号、虎老溪2号、虎老溪1号、酉阳3号、酉阳9号、酉阳11号、酉阳10号、酉阳12号、酉阳1号16份资源,第2个亚群包括酉阳5号、酉阳8号、酉阳6号、酉阳14号4份资源;第Ⅱ类群包含5份茶树资源:永川1号、永川2号、永川4号、永川5号、永川3号;第Ⅲ类群也包含2个亚群,亚群1包含木皮槽2号、木皮槽1号,亚群2包括南川2、南川1、南川3共计3份资源。 图1 重庆30份茶树资源聚类分析 对3个类群的农艺性状和主要生化成分进行统计分析,结果见表5,由表5可知,3个类群在一芽二叶长、一芽二叶百芽重、一芽一叶期、叶尖、叶缘、咖啡碱、氨基酸、茶多酚8个性状上存在显著差异。第Ⅰ类群主要表现为灌木型茶树,中小叶种,平均叶脉对数6~7对,叶尖渐尖,叶缘微波,叶面微隆,叶身较平,芽叶绒毛较多,芽叶色泽黄绿或淡绿色为主,一芽二叶长和一芽二叶百芽重中等,一芽一叶期和一芽二叶期较晚,萼片数5~6片,花瓣5~6瓣,花瓣颜色以淡绿色居多,质地中等,花冠直径较短,花柱长度较短,柱头3裂为主,裂位较高,子房有绒毛,咖啡碱、氨基酸、茶多酚含量中等,水浸出物较少;第Ⅱ类群主要表现为灌木型茶树,中小叶种,平均叶脉对数6~7对,叶尖渐尖或钝尖,叶缘波,叶面平,叶身内折,芽叶绒毛较多,芽叶色泽黄绿色,一芽二叶长和一芽二叶百芽重较小,一芽一叶期和一芽二叶期最早,萼片数4~5片,花瓣6~7瓣,花瓣颜色白色或淡绿色,质地中等,花冠直径较短,花柱长度较短,柱头3裂,裂位较低,子房有绒毛,咖啡碱、茶多酚含量较少,氨基酸含量较高,水浸出物较少;第Ⅲ类群与前2个类群差异较大,主要表现为乔木或小乔木型茶树,大、中叶种,平均叶脉对数7对,叶尖急尖,叶缘平,叶面平,叶身内折,芽叶绒毛较少,芽叶色泽淡绿色,一芽二叶长和一芽二叶百芽重较大,一芽一叶期和一芽二叶期较晚,萼片数5~6片,花瓣7~9瓣,花瓣白色,质地较厚,花冠直径较大,花柱长度较长,柱头3~4 裂,裂位较高,咖啡碱、茶多酚含量较高,氨基酸含量较少,水浸出物较多。 表5 不同类群间农艺性状与主要生化成分比较 续表5 Continued Table 5 特异性茶树种质资源是突破性茶树品种培育的基础,具有重要的科学意义和经济价值。参照《NY/T2031—2011农作物优异种质资源评价规范 茶树》中特异茶树种质资源筛选标准对30份重庆茶树资源进行筛选(表6),共筛选出特早生茶树资源(一芽一叶期比福鼎早10 d)10份,其中,有3份资源平均一芽一叶期比福鼎早24 d;筛选出白色芽变资源1份,多芽叶绒毛资源3份,高茶多酚总量(≥25%)资源21份,其中,茶多酚总量超过30%的茶树资源6份;筛选出高咖啡碱(≥5.0%)资源4份。经过连续3 a的测定,这些基于农艺性状或主要生化成分呈现出来的特异性茶树资源表现稳定,可为优异茶树品种的培育与遗传研究提供材料基础。 表6 不同类群间农艺性状及主要生化成分比较 茶树农艺性状的评价一直是茶树种质资源鉴定和评价的主要内容,近几十年,利用生物化学的方法开展茶树种质资源的研究也取得了较大进展[13]。本研究采用农艺性状结合主要生化成分评定的方法对30份重庆茶树种质资源进行鉴定评价。结果表明,重庆茶树资源在农艺性状与主要生化成分上存在不同程度的变异。对比其他地区茶树种质资源的多样性分析结果,重庆茶树种质资源农艺性状多样性较为丰富,平均多样性指数均高于武夷名丛(1.09)[9]、云南茶树种质资源(1.04)[14]、四川中小叶茶树群体种(0.98)[15]、陕西茶树种质资源(1.11)[16],证明重庆茶树种质资源农艺性状类型丰富,成分较复杂,离散程度较高,具有较强的育种潜力。重庆茶树种质资源主要生化成分的平均变异系数较低,低于贵定(22.37%)[17]、陕西(21.78%)[16]、广西(25.80%)[18]、四川(26.80)[19],高于云南地方茶树品种(16.53%)[3],但多样性指数高于贵定(1.15)、陕西(1.89)[16]、广西(1.90)[18]、四川(1.92)[19]。与其他地方相比,重庆茶树资源平均水浸出物含量较低(36.48%),低于广西(41.9%)[18]、河南(43.17%)[20]、四川(51.7%)[19];咖啡碱平均含量较高(3.95%),高于河南(2.88%)、四川(3.0%)、广西(3.6%);茶多酚平均质量分数(28.15%)处于中等水平置,低于广西(28.7%)、广东(31.09%),高于贵州(17.94%)、河南(18.47%)、四川(21.9%);氨基酸平均质量分数较高(3.18%),仅低于贵州(3.66%),高于四川(2.9%)、广西(2.9%)、河南(3.01%)、广东(3.04%)等地。以上研究结果表明,不同省份茶树资源主要生化成分含量差异较大,变异系数和多样性指数也有很大不同,与其他省份相比,重庆茶树种质资源具有较高的咖啡碱和氨基酸含量,茶多酚含量居中,但水浸出物含量相对较少,主要生化成分的平均变异系数较低,但咖啡碱和氨基酸的变异系数较高,证明重庆茶树种质资源在咖啡碱和氨基酸含量上有很大选种潜力。主要生化成分多样性指数均较高,说明这些资源的遗传多样性丰富。以上研究表明,重庆茶树种质资源具有较为明显的地方特色,具有较为丰富的农艺性状变异类型和生化成分遗传多样性,这可能与重庆地形复杂、气候多变的环境因素有关。 主成分分析可清晰地反映群体差异的主要来源,通过降维的方法分类出几个综合因子反映出众多性状指标的信息量,简化研究性状。对30份茶树资源的35个性状进行主成分分析,入选的9个主成分,累计贡献率达74.85%,特征值总和为20.469,包含了茶树的大部分特征信息,可以用来综合评价重庆茶树种质资源,且不同的性状在主成分中具有明显不同的荷载值,有些性状对主成分的影响为负值。这与田甜等[7]、王飞权等[9]、谢文钢等[15]的研究结果类似。 通过聚类分析,在欧式平方距离8处,将30份重庆茶树资源分为3个大类,这3个类群在一芽二叶长、一芽二叶百芽重、一芽一叶期、叶尖、叶缘、咖啡碱、氨基酸、茶多酚等性状上存在显著差异,尤其是第Ⅲ类群在除叶脉对数、萼片数、叶形、叶色、花萼色泽、花柱裂位、雌雄蕊相对高度、叶面、叶身、可溶性糖以外的25个性状指标上均与其他2个类群存在显著差异。第Ⅲ类群包含了南川大树茶和江津木皮槽2个小类群。南川大树茶为典型的野生大茶树类型,具体表现为:乔木型、大叶种、花瓣直径较大、花柱长度较长、柱头裂位较多、丛瓣、芽叶无绒毛、子房无绒毛,咖啡碱、茶多酚、水浸出物含量较高,氨基酸含量较低;江津木皮槽茶树资源表现为:小乔木型、中叶种、花瓣直径较大、花柱长度较长、芽叶绒毛较少或没有,茶多酚、咖啡碱含量较高,氨基酸含量高于南川大茶树,属于由野生茶树向栽培型茶树演化的中间阶段。第Ⅰ类群和第Ⅱ类群均属于栽培型茶树,第Ⅰ类群包含了酉阳宜居茶和武隆虎老溪茶2个群体种,它们最早都是由川茶群体种演化而来,与其他2个类群相比,第Ⅰ类群一芽一叶长和一芽二叶百芽重中等,萌芽期较晚,叶尖渐尖,叶缘微波,咖啡碱、氨基酸、茶多酚含量在3个类群中处于中等水平。永川茶树资源单独聚为第Ⅱ类群,由福鼎大白茶与川茶群体种杂交后代单株选育而成,突出特点为萌芽期特早,与其他2个类群相比,第Ⅱ类群一芽一叶长和一芽二叶百芽重最小,萌发期最早,叶尖渐尖或钝尖,叶缘波,咖啡碱、茶多酚含量在3个类群中最低,氨基酸含量最高。 根据陈文怀[21]对中国茶树品种演化的研究,茶树由野生种向栽培种进化的进程中,会由乔木(第Ⅲ类群南川群体)到小乔木(第Ⅲ类群木皮槽)再到灌木(第Ⅰ、Ⅱ类群),芽叶绒毛由多到少[第Ⅲ类群(无或少)<第Ⅰ类群<第Ⅱ类群],叶片由大叶(第Ⅲ类群南川群体为大叶)演化为中小叶(第Ⅰ、Ⅱ类群),花冠由大到小,花瓣由丛瓣到单瓣[第Ⅲ类群(大花冠,有丛瓣也有单瓣),第Ⅰ、Ⅱ类群(小花冠,单瓣];内含物质成分也会在茶树品种演化过程中不断变化,水浸出物由多到少(第Ⅲ类群>第Ⅰ、Ⅱ类群),茶多酚含量由高到低(第Ⅲ类群>第Ⅱ类群>第Ⅰ类群),氨基酸含量则由低到高(第Ⅲ类群<第Ⅱ类群<第Ⅰ类群)。由以上研究结果可以推测,进化程度为第Ⅱ类群>第Ⅰ类群>第Ⅲ类群木皮槽>第Ⅲ类群南川群体,重庆茶树资源既有最原始最古老的五室茶系(第Ⅲ类群南川群体)[22],又有各种中间过渡类型,以及进化程度较高、接近栽培型的茶树种质资源,资源类型丰富,可为茶树种质资源演化研究和新品种选育提供较多较好的研究材料。 重庆作为老茶区,虽然茶树栽培历史悠久,但品种单一,良种化率低,大部分产茶区县以川茶群体种为主,部分主打野生大树茶的区县也以大树茶群体种扦插繁殖,性状混杂、品质良莠不齐。重庆虽然坐拥农艺性状丰富且遗传多样性的茶树资源宝库,却没有进行充分的发掘利用。本研究聚类的第Ⅲ类群水浸出物含量丰富,茶多酚总含量均超过33%,酚氨比较高,是开发红茶的优质原料。第Ⅰ类群属于红绿兼制类型;其中的酉阳6号,春季芽叶为乳白色,经过多年观察测定,该性状遗传稳定,且与市场同类型品种相比,酉阳6号生长势旺,扦插成活率高,抗逆性强,适应性广,是特异性品种培育的优良材料;酉阳9号、酉阳13号和酉阳14号是芽叶绒毛特多类型,芽叶绒毛的多少是茶毫多少的决定因素,也是评价茶叶外形和内质的重要影响因子[23],用其开发的名优绿茶和红茶会更具有市场竞争力。第Ⅱ类群也属于红绿兼制类型,平均酚氨比5.83,氨基酸含量较高,平均氨基酸含量在4%以上,茶多酚含量也较高,平均茶多酚含量为23.35%,具有制备优质绿茶和优质红茶的物质基础;且该类群的萌芽期较早,除永川4号一芽一叶期比福鼎早12 d以外,其余4份材料一芽一叶期均比福鼎早22 d以上。除此以外,第Ⅰ类群的酉阳10号、酉阳15号,第Ⅲ类群的南川3号、南川1号、木皮槽1号也属于特早生资源,可以更早上市,抢占名优绿茶与红茶市场。30份资源中有21份材料属于高茶多酚特异资源,4份材料属于高咖啡碱特异资源,可以用于成分提取,保健型功能产品、饲料添加剂、保鲜剂、抑菌剂和新型功能茶产品的开发。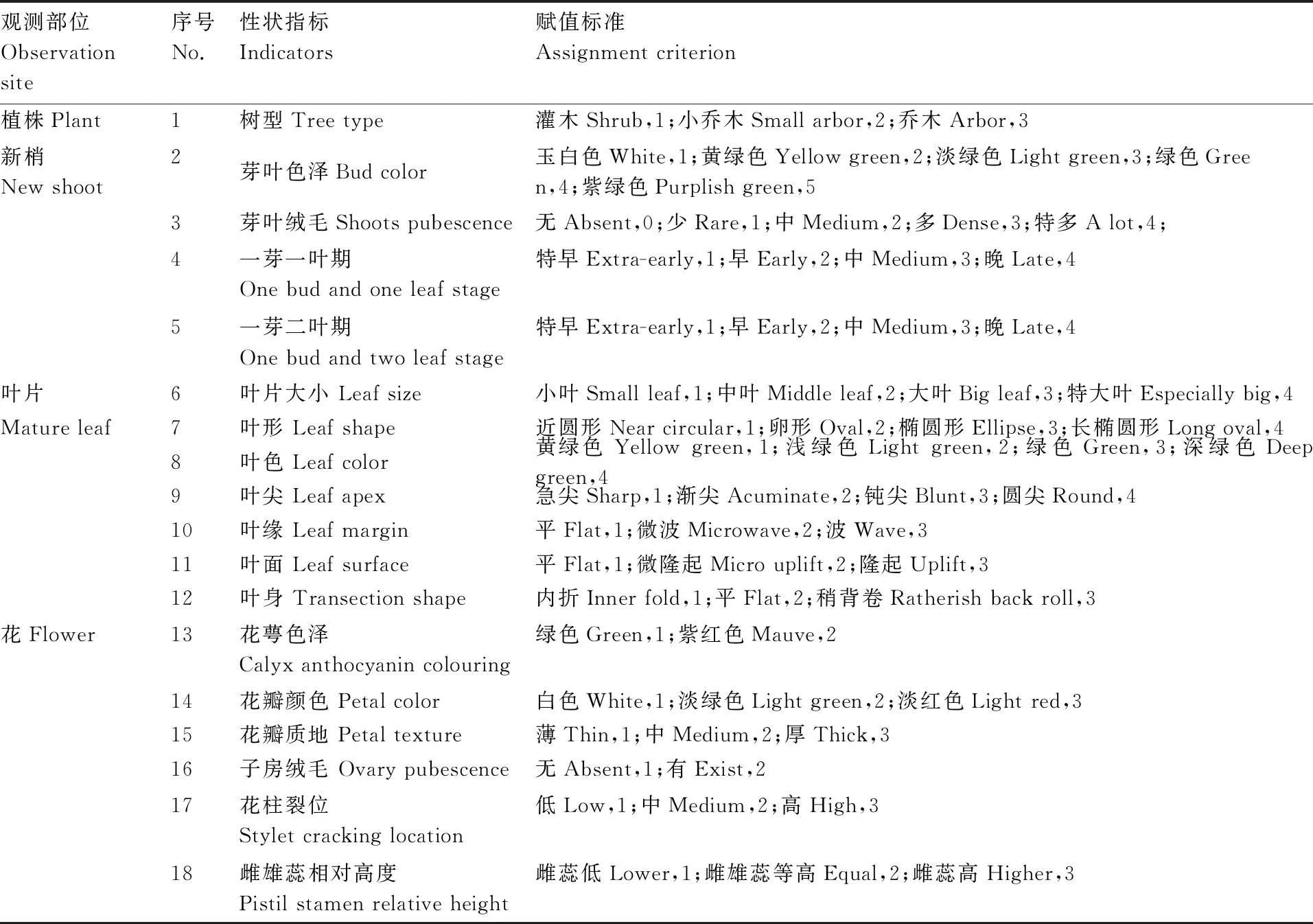
2 结果与分析
2.1 农艺性状与主要生化成分的变异和多样性

2.2 农艺性状与主要生化成分的主成分分析

2.3 基于农艺性状和主要生化成分的聚类分析

2.4 农艺性状和主要生化成分的特异茶树种质资源筛选
3 结论与讨论
