30份建兰种质资源的表型性状遗传多样性研究
2021-08-04王宏利曾艳华卜朝阳
王宏利 曾艳华 卜朝阳

摘 要:以30份不同來源的建兰种质资源为供试材料,测定其25个表型性状,计算其各性状的变异系数,并进行主成分分析和聚类分析,以期揭示建兰各品种间的亲缘关系及遗传差异。结果表明,30个建兰供试品种各表型性状的变异幅度为9.28%~76.17%,各表型性状间均存在较大程度的变异;表型性状聚类分析结果显示30个建兰供试品种欧式距离范围为3.612~38.796。‘汇翠粉荷和‘峨眉水仙的欧式距离最小,为3.612,表明二者的亲缘关系最近;‘素君荷和‘富山奇蝶之间的欧式距离最大,为38.796,表明二者的亲缘关系最远;其他材料之间的亲缘关系位于二者之间。对30个供试品种的表型性状使用层次聚类法进行树状图绘制,在欧氏距离为16处,30个建兰供试品种分成4个群集;主成分分析结果表明贡献值最大的花部性状有花葶高度、花长和唇瓣长等性状,均与花部质量有关,花部性状的综合评分表明广东佛山的‘红君荷花部综合特性最好,观赏价值最高,广东佛山的‘水晶皇梅综合特性最低。
关键词:建兰;表型性状;主成分分析;聚类分析
中图分类号:S682.31 文献标识码:A
Genetic Diversity of Phenotypic Traits of 30 Cymbidium ensifolium Germplasm Resources
WANG Hongli, ZENG Yanhua, BU Chaoyang
Flower Research Institute, Guangxi Academy of Agricultural Sciences, Nanning, Guangxi 530007, China
Abstract: 30 species of Cymbidium ensifolium from different resources were selected as the research materials. The phenotypic trait measuring, variation coefficients of each trait, principal component analysis and cluster analysis were conducted to reveal the genetic relationship of the germplasms to provide basis for the identification of varieties and cross breeding of C. ensifolium. The analysis of phenotypic traits showed that the variation range of each phenotypic trait was 9.28%-76.17%, and there was a large degree of variation among all phenotypic traits. The results of cluster analysis showed that the range of euclidean distance of the 30 experimental materials was 3.612-38.796. ‘Huicui Fenhe and ‘Emei Narcissus had the smallest euclidean distance of 3.612, indicating the closest genetic relationship, while ‘Sujunhe and ‘Fushan Qidie had the largest euclidean distance of 38.796, indicating the furthest genetic relationship. The hierarchical clustering method was used to draw the dendrogram of the phenotypic traits of the 30 test materials. At the Euclidean distance of 16, the 30 test varieties are divided into 4 clusters. The principal component analysis results showed that the largest contribution value included the scape height, flower length, and lip length which are all related to the quality of flowers. The comprehensive score of the flower traits showed that the comprehensive characteristics of ‘Hongjunhe from Foshan, Guangdong were the best and the ornamental value was the highest, and ‘Shuijing Huangmei from Foshan, Guangdong had the lowest comprehensive characteristics.
Keywords: Cymbidium ensifolium; phenotypic traits; principal component analysis; cluster analysis
DOI: 10.3969/j.issn.1000-2561.2021.06.007
建兰[Cymbidium ensifolium (L.) Sw.]属于国兰的一大种群[1]。其花浅黄绿色,芳香。与其他种类国兰明显区别是,其花期较长且可多次开花(1年可达6次),故又名四季兰。常见栽培品种包括:‘金丝马尾‘凤尾素‘小桃红‘龙岩素‘铁骨素‘君荷‘朝阳三星蝶等。建兰喜温暖湿润和半阴环境,耐寒性差,越冬温度不低于3 ℃,怕强光直射,不耐水涝和干旱,宜疏松肥沃和排水良好的腐叶上。建兰不仅观赏价值高,还可以入药使用,有极高的经济价值。
表型性状多样性是遗传物质多样性的具体表现,是进行遗传多样性研究最基本最重要的组成部分[2]。目前,我国兰属植物的研究主要集中在兰花的鉴别分类[3-5]、高效栽培[6-9]、组织培养[10-12]及分子调控机理[13-15]等方面。广西农业科学院花卉研究所的卜朝阳等在春兰、豆瓣兰、蕙兰等国兰资源收集繁育及育种研究方面取得了较好成果,但在兰属植物各种质的遗传背景领域等方面研究还鲜见报道。曾碧玉等[16]收集了大量各地区的兰属品种并对兰花选育种研究进展进行了综述;傅巧娟等[17]则从细节出发,调查兰属植物的株形、叶片等农艺特征在品种间的相关性,得出了兰属种质表型多样性丰富,数量性状变异程度高的结论,间接地验证了中国植物志关于兰属种质多样性丰富的相关理论。张韶伊等[18-19]分别对蕙兰及春兰的特征和生物学性状进行了调查研究;姜殿强等[20]对建兰、寒兰的花表型结构进行了分析研究。本研究通过对广西农业科学院花卉研究所资源圃内的30份建兰种质资源的25个表型性状进行测定,分析其资源的多样性和亲缘关系,以期为兰属植物的分类和资源进一步评价利用提供参考依据。
1 材料与方法
1.1 材料
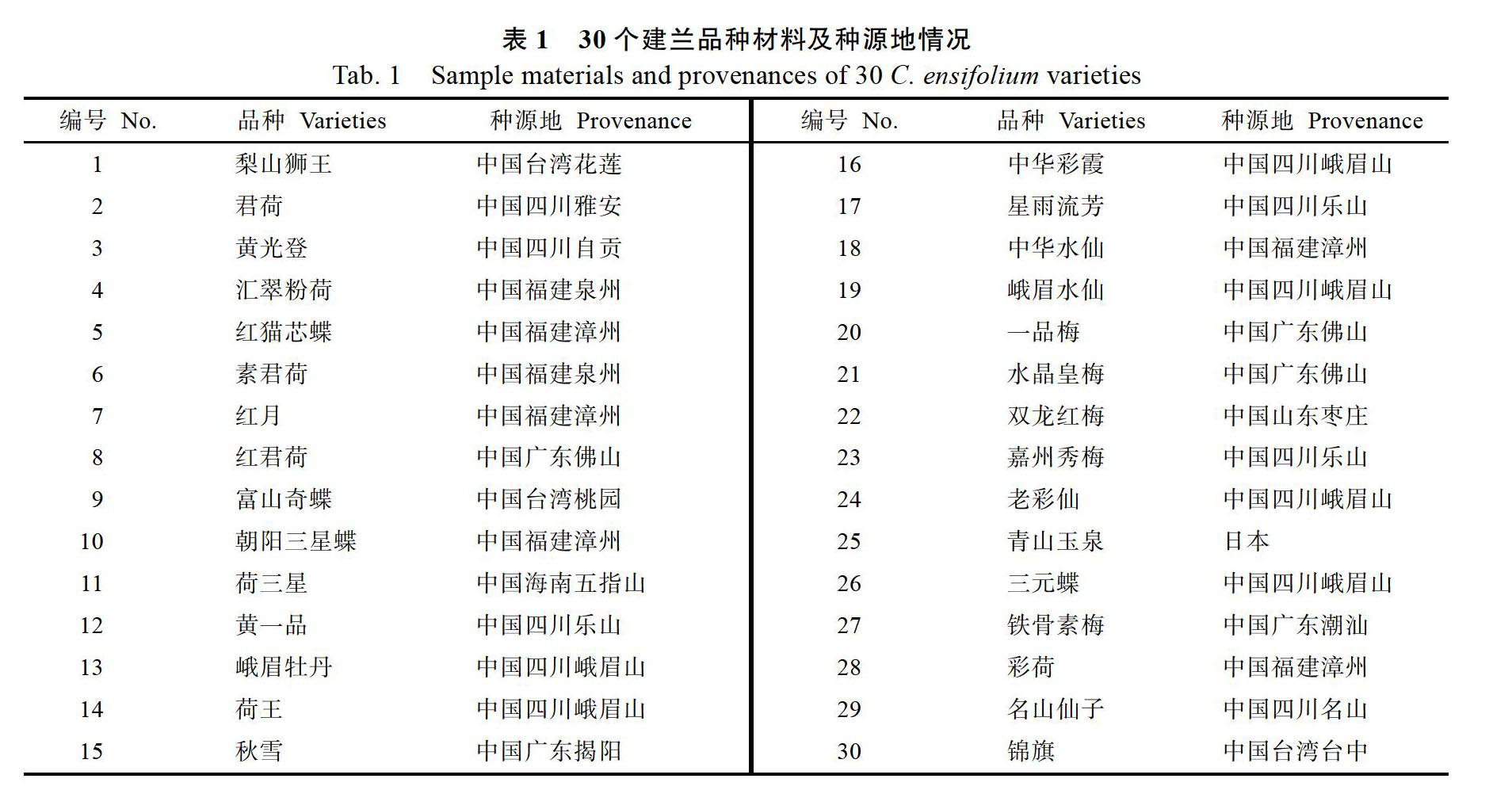
实验所用的30个建兰品种(表1)是广西农业科学院花卉研究所科研人员于2017年6月陆续从台湾、四川、福建、广东等地收集而来,种植在广西农业科学院花卉研究所资源圃内。
每个建兰品种种植8株,每份材料都种植于0.15 m2的实验区域内,间距30 cm,15 d浇水1次,每周施1次花多多氮、磷、钾稀薄液肥,用量5 g,施肥要宁淡勿浓,每天维持10 h光照等常规的栽培方式进行统一管理。
1.2 方法
1.2.1 表型性状的选择及数据采集 由于兰属植物是以观叶赏花为主,评价兰属植物的重要指标是兰花的叶姿、叶艺和花的颜色、形状等特征,根据云南省农业科学院质量标准与检测技术研究所制定的兰属植物品种特异性、一致性和稳定性测试指南,选取了株高、叶长、花瓣形状、花葶着花数等25个具有代表性且遗传相对稳定的表型性状进行测试,当待测品种在盛花期时分别选取健康单株用卷尺、游标卡尺对各个性状进行测量,然后进行统计分析。
1.2.2 性状评价标准 参考敖素燕等[21]的31个国兰品种的数量性状分析及国兰表型性状与核型分析方法,确定建兰测试性状及方法见表2。在建兰花朵开放到最佳状态时,记录表2的各项指标,各项指标的观察样本数不少于5株,以實测值求平均数对数量性状、赋值后的质量性状(表3)进行记录和分析。
1.3 数据处理
所测表型数据进行标准化处理后,使用Excel软件对质量性状出现频率和表型变异系数进行处理,用社会科学统计软件SPSSAU在线系统进行主成分分析及聚类分析。
2 结果与分析
2.1 30个建兰品种质量性状出现的频率
30个建兰品种11个质量性状的多样性统计分析见表4。不同表型性状在群体中出现的频率不同,其中主花色为粉色或者红色类型、叶艺为无类型、萼片及花瓣姿态为全内拱、唇瓣为有斑点类型、花香浓度为香味较明显类型等性状在各表型性状中出现的频率相对较高,是具有代表性的表型性状。
2.2 30个建兰品种表型性状的变异分析
对30个建兰品种各表型性状进行统计分析。从图1、表5可见,建兰试验材料遗传变异丰富,
各表型性状的变异幅度为9.28%~76.17%,说明存在较大程度的变异。变异系数从大到小依次为:唇瓣形状(76.17%)>唇瓣(74.52%)>叶艺(62.51%)>萼片及花瓣姿态(53.29%)>叶片形状(49.79%)>萼片宽(38.25%)>花瓣形状(36.96%)>花长(36.63%)>花葶着花数(34.70%)>花梗粗(32.65%)>唇瓣长(32.28%)>花宽(31.15%)>主花色(30.95%)>花葶高度(28.45%)>花瓣颜色数量(27.90%)>叶色(26.26%)>叶宽(26.25%)>唇瓣宽度(25.45%)>萼片长(23.94%)>叶片数(23.06%)>株高(22.05%)>叶长(18.72%)>花香浓度(17.70%)>株型(13.12%)>叶尖端形状(9.28%)。唇瓣形状的变异系数为76.17%,在25个表型性状中最高,叶尖端形状变异系数为9.28%,在25个表型性状中最低,其余性状的变异幅度处于唇瓣形状和叶尖端形状之间。
2.3 花部性状主成分分析
根据特征根大于1的原则提取了前6个主成分,特征根分别为4.049、2.373、1.838、1.321、1.254、1.094,贡献率分别为25.305%、14.833%、11.485%、8.255%、7.837%和6.84%,累计贡献率为74.557%,这6个主成分几乎涵盖了影响建兰花部表型性状的大部分因素。由建兰花部旋转后的成分矩阵表6可知,萼片长、花长、花宽、萼片及花瓣姿态、花瓣颜色数量、唇瓣长、唇瓣宽和唇瓣等8个性状包含在第1主成分当中,萼片及花瓣形状、花瓣颜色数量与其他各性状呈负相关,其余性状间呈正相关。其中,在第1主成分中萼片长的权重系数最大,其值为0.913,权重系数最小的是唇瓣,其值为0.472,通过查看成分得分系数矩阵表,旋转因子的得分分别为0.225、0.117。花葶着花数、花葶高度、花梗粗、花瓣形状、唇瓣宽度、唇瓣形状、花香浓度7个性状包含在第2主成分当中。第3主成分包括花葶着花数、主花色、唇瓣3个性状,三者呈正相关,主花色(0.687)比萼片宽(?0.515)的权重值要大。第4主成分包括萼片宽、唇瓣形状2个性状,与其他性状存在正相关。第5主成分包括花葶高度、花梗粗、萼片宽和唇瓣长度4个性状。第6主成分只包括花香浓度1个性状。可见,花葶高度、花长和唇瓣长等性状在花部性状中贡献值最大,这些性状都和花部质量联系紧密;其次是花葶着花数、萼片长、萼片宽和花色等性状,这些性状都与花部视觉感官有关。
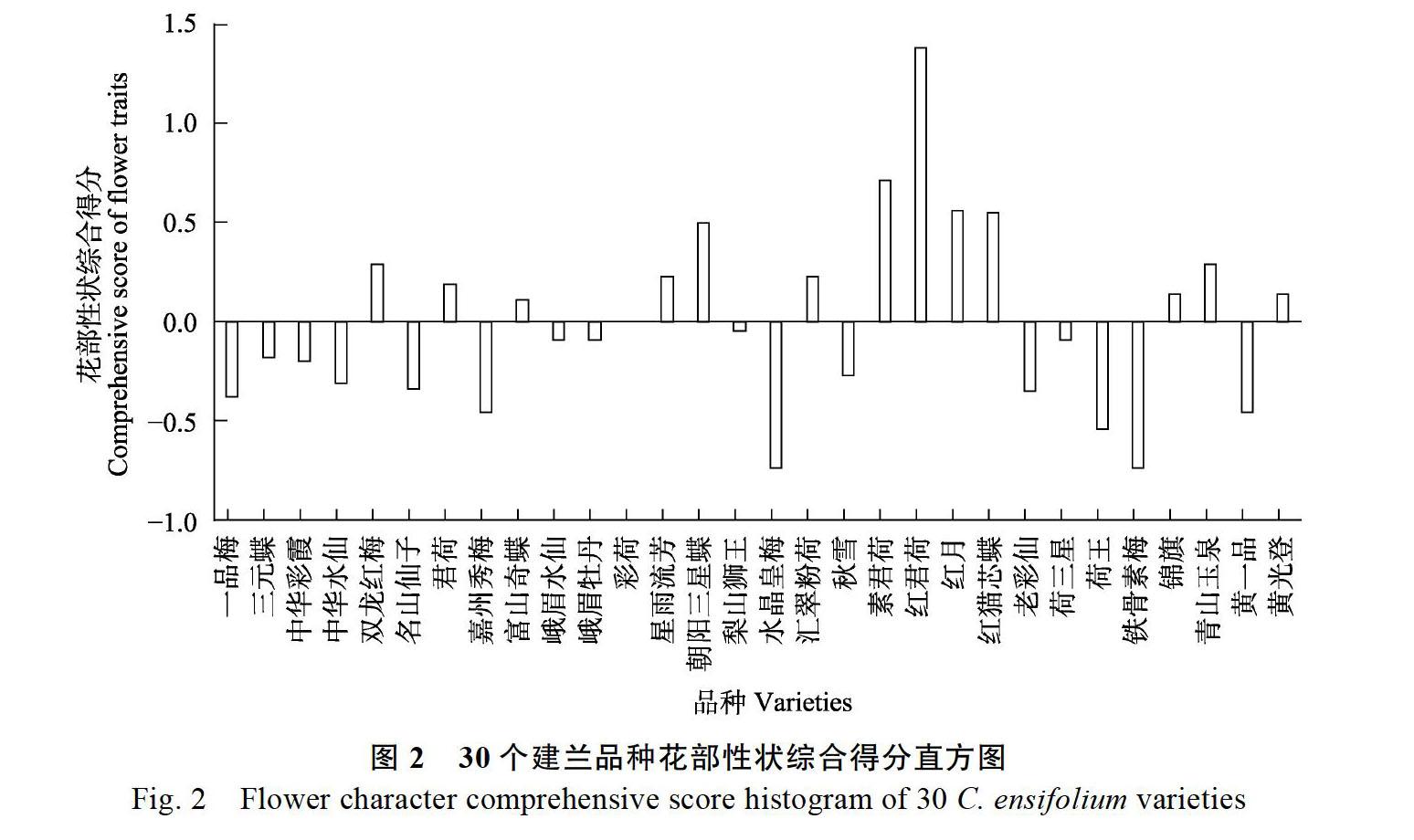
花部性状主成分因子得分表见表7。由表7可知,花部性状综合得分最高的是8号广东佛山的‘红君荷,花部性状综合得分最低的是21号广东佛山的‘水晶皇梅。
根据建兰品种花部性状的综合得分绘制柱形图。由图2可知,花部综合特性最好、观赏价值最高的是8号广东佛山的‘红君荷。6号福建泉州的‘素君荷、7号福建漳州的‘红月、10号福建漳州的‘朝阳三星蝶花部综合得分都达到了0.50以上,综合特性表现较好,21号广东佛山的‘水晶皇梅综合特性最低,其他建兰品种的花部综合特性介于二者之间。综上,花部性状综合得分高的建兰品种具有花葶着花数相对其他品种多,花型的花朵饱满,萼片长、宽尺度相对较大的特点,观赏价值比较高。
2.4 聚类分析
使用SPSSAU软件计算30份建兰种质间的欧式距离,其范围为3.612~38.796,其中,4号福建泉州的‘汇翠粉荷和19号四川峨眉山的‘峨眉水仙的欧式距离最小,为3.612,表明二者的亲缘关系最近;6号福建泉州的‘素君荷和9号台湾桃园的‘富山奇蝶之间的欧式距离最大,为38.796,表明二者的亲缘关系最远,其他材料之间的亲缘关系位于二者之间。
对30个供试材料的表型性状使用层次聚类法进行树状图绘制(图3),结果显示,在欧氏距离为16处,30个建兰供试品种分成4个群集。
类群Ⅰ有:‘汇翠粉荷‘峨眉水仙‘铁骨素梅‘峨眉牡丹‘红月‘一品梅‘老彩仙‘红君荷‘名山仙子‘中华水仙‘水晶皇梅‘三元蝶‘双龙红梅‘黄一品‘青山玉泉‘朝阳三星蝶‘星雨流芳‘彩荷;
类群Ⅱ有:‘梨山狮王‘荷三星‘秋雪‘荷王‘红猫芯蝶‘中华彩霞‘富山奇蝶‘君荷‘黄光登;
类群Ⅲ有:‘嘉州秀梅‘锦旗;
类群Ⅳ有:‘素君荷。
在欧氏距离为14处,可以将类群Ⅰ划分为2个亚类,Ⅰ-i亚类包括4号福建泉州的‘汇翠粉荷、19号四川峨眉山的‘峨眉水仙、10号福建漳州的‘朝阳三星蝶等16份材料,该类材料的特点为:除了‘红君荷外,其余材料花梗粗、唇瓣长等性状均处于中下水平;Ⅰ-ii亚类只包括17号四川乐山的‘星雨流芳、28号福建漳州的‘彩荷2份材料。该类所有材料叶片长、叶宽及叶片数均处于最低水平。类群Ⅱ中的‘汇翠粉荷瓣型规整大气,该类材料的特点为花葶着花数、花葶高度等性状均处于中上水平。类群Ⅲ中的2份建兰材料各性状参数接近,二者表型性状除叶艺和主花色外其他特征相似度高。类群Ⅳ中‘素君荷各个性状表现都较好,它在14个数量
性状的观测中表现状态佳,观赏价值高,能够满足市场需要。
不同类群主要性状指标平均值见表8。可见,类群Ⅰ的平均株高为26.19 cm,平均叶长为37.69 cm,平均叶宽为1.54 cm,平均花葶高度为18.78 cm,平均花梗粗为2.88 mm,平均花长为3.04 cm,平均花宽2.97 cm,这一类群的显著特点是株高、叶长、叶宽、花葶高度、花梗粗和花长、宽都适中。类群Ⅱ的平均株高为22.72 cm,平均叶长为28.30 cm,平均叶宽为1.48 cm,平均花葶高度为14.96 cm,平均花梗粗为2.57 mm,平均花长为3.40 cm,平均花宽2.90 cm,这一类群在4个类群中性状最差,除花长外其余性状的观测平均值都是最低的。类群Ⅲ的平均株高为38.00 cm,平均叶长为40.05 cm,平均叶宽为1.53 cm,平均花葶高度为25.40 cm,平均花梗粗为2.92 mm,平均花长为3.50 cm,平均花宽2.95 cm,这一类群在4个类群中性状较优,仅次于类群Ⅳ。类群Ⅳ的平均株高为39.30 cm,平均叶长为51.30 cm,平均叶宽为1.90 cm,平均花葶高度为34.50 cm,平均花梗粗为2.93 mm,平均花长为5.60 cm,平均花宽4.60 cm,这一类群在4个类群中性状最优,所有主要数量性状的观测值都是最高的。
3 讨论
表型性狀分析结果表明:建兰实验材料各表型性状的变异幅度为9.28%~76.17%,各表型性状间存在较大程度的变异。表明建兰植物种质材料的数量性状变异大,遗传多样性丰富,可以利用系统选育及杂交育种等方法从中选育出性状优良的建兰新品种。在本次研究中,很多数量性状集中于花部,针对花部性状进行分析可知,相对于其在种内所呈现出来的较高稳定性而言,其在种间则表现出来一定的变化,因此,此类性状能够有助于品种的分类与鉴定。花部性状综合得分高的建兰材料具有花葶着花数相对其他品种多,花
朵饱满,萼片长、宽尺度相对较大的特点,观赏价值比较高。广东佛山的‘红君荷花部综合特性最好,观赏价值最高。广东佛山的‘水晶皇梅综合特性最低,观感较差,这和实际观测兰花田间表现具有一致性。部分数量性状在品种间会表现出差异,变异程度往往会受到其品种的不同性状而影响,相对于蕊柱长度和宽度只在少数品种之间表现出极明显的差异性来说,萼片和花瓣的长度所表现出来的性状往往会在很多的品种中表现出明显的差异。在建兰种内,花葶在花数、萼片的长宽这几个性状当中差异并不大,而在种间则表现出明显的差异,出现这种情况的原因可能
与建兰一葶多花的特征有关。由于数量性状的局限性,在今后工作中可以更深入从花色、花瓣、花型、叶姿、叶艺等其他表型性状上展开系统化的研究,以期为建兰新品种的选育奠定基础。
30份建兰种质在欧氏距离为16处被分别归于4个类群。并且,当这些材料进行交叉聚集时,其结果总体上与其所表现出来的表型性状变异规律一致。表型聚类中,建兰供试材料根据株高和花葶着花数等性状的差异划分为三大类,5种建兰品种材料的唇瓣长、花葶高度等性状均处于中等偏上水平聚在1个类群中;中国四川峨眉的‘中华彩霞和日本的‘青山玉泉未聚在同一类群,可能的原因为材料来源地差异过大,二者性状由于环境的差异而出现了不同。表型性状多样性受生境条件和空间距离等多因素的影响,是在形态水平上对遗传多样性的展现,代表着物种的变异程度大小和遗传多样性[22-23]。就本研究而言,研究过程中所选用的供试品种所适应的环境复杂多样,因而在广西南宁市的同一试验地进行种植的过程中,针对其遗传相似性,会出现其表型性状无法准确地真实反映遗传关系的情况。这一结果和张冬玲等[24]的报道,当所研究的群体分布过于广泛且存在其外界生态环境过于复杂的情况时,会出现表型差异明显的结果一致。
本研究明确了广西农业科学院花卉研究所收集保存的30份建兰种质资源的表型性状和遗传变异相近程度,可以为今后兰属杂交选育及杂交种间遗传多样性研究提供基础数据;筛选获得综合特性好观赏价值高的‘红君荷,可以为居家赏兰提供参考材料。
参考文献
[194] 刘仲健, 章迪杨, 朱雅婷, 等. 中国兰属[J]. 森林与人类, 2019(11): 98-103.
[195] 杨乐琦, 仇淑芳, 唐 菲, 等. 基于主成分分析的观赏生菜品质综合评价[J]. 上海交通大学学报(农业科学版), 2015, 33(3): 53-60.
[196] 景袭俊, 胡凤荣. 兰科植物研究进展[J]. 分子植物育种, 2018, 16(15): 5080-5092.
[197] 韩 东. 国兰市场调查及国兰资源保护研究[D]. 北京: 北京林业大学, 2009.
[198] 邓银霞. 中国兰品种资源与分类研究[D]. 长春: 吉林农业大学, 2008.
[199] 黄萍萍, 刘 霖, 陈红玉. 建兰的栽培管理[J]. 现代园艺, 2014(16): 37.
[200] 陈翠云. 建兰无土栽培的基质筛选的研究[J]. 嘉应学院学报, 2009, 27(3): 88-92.
[201] 徐久生. 淺谈建兰的栽培与管理[J]. 现代园艺, 2012(12): 42.
[202] 吴俊玲. 建兰的栽培管理技术[J]. 福建农业科技, 2008(2): 26-27.
[203] 陈 春. 建兰‘岭南奇蝶种子无菌萌发与离体繁殖技术[J]. 亚热带农业研究, 2016, 12(2): 125-129.
[204] 王济红, 刘 燕, 祁 翔, 等. 建兰新品种黄金小神童组培育苗集成技术的优化[J]. 西南农业学报, 2014, 27(5): 2135-2140.
[205] 叶秀仙, 黄敏玲, 林榕燕, 等. 素心建兰无菌播种快繁技术研究[J]. 福建农业学报, 2015, 30(10): 939-943.
[206] Li X B, Jin F, Jin L, et al. Characterization and comparative profiling of the small RNA transcriptomes in two phases of flowering in Cymbidium ensifolium[J]. BMC Genetics, 2015, 16(1): 622-638.
[207] Li X B, Luo J, Yan T L, et al. Deep sequencing-based analysis of the Cymbidium ensifolium floral transcriptome[J]. PLoS One, 2013, 8(12): 1-13.
[208] Yang F X, Zhu G F, Wang Z, et al. A putative miR172-targeted CeAPETALA2-like gene is involved in floral patterning regulation of the orchid Cymbidium ensifolium[J]. Genetics and Molecular Research, 2015, 14(4): 12049-12061.
[209] 曾碧玉, 朱根发, 刘海涛. 兰花选育种研究进展[J]. 中国农学通报, 2005(12): 272-276.
[210] 傅巧娟, 李春楠, 赵福康, 等. 7种兰属种质表型性状遗传多样性分析[J]. 分子植物育种, 2018, 16(10): 3381-3394.
[211] 张韶伊. 大花蕙兰品种DUS测试及性状间的相关性分析[D]. 杭州: 浙江农林大学, 2013.
[212] 张韶伊, 范义荣, 褚 怡, 等. 春兰属植物表型性状的多样性研究[J]. 北方园艺, 2013(12): 68-72.
[213] 姜殿强, 李良珍, 杨 珺, 等. 建兰、寒兰花表型分析[J]. 现代园艺, 2017(21): 3-5.
[214] 敖素燕, 范义荣, 段艳皊, 等. 31个国兰品种的数量性状分析[J]. 江苏农业科学, 2014, 42(9): 131-134.
[215] 张冰冰, 宋洪伟, 刘慧涛, 等. 寒地梨种质资源表型多样性研究[J]. 果树学报, 2009, 26(3): 287-293.
[216] 王力荣, 朱更瑞, 方伟超. 桃种质资源若干植物学数量性状描述指标探讨[J]. 中国农业科学, 2005(4): 770-776.
[217] 张冬玲, 张洪亮, 魏兴华, 等. 贵州栽培稻的遗传结构及其遗传多样性[J]. 科学通报, 2006(23): 2747-2754.
责任编辑:沈德发
