南海珊瑚礁海域黑缘尾九棘鲈生物学特征初步研究*
2021-08-04洪小帆陈作志江艳娥张俊王欢欢李媛洁李纲
洪小帆, 陈作志, 江艳娥, 张俊, 王欢欢, 李媛洁, 李纲
1. 上海海洋大学海洋科学学院, 上海 201306;
2. 中国水产科学研究院南海水产研究所, 农业农村部外海渔业开发重点实验室, 广东 广州 510300;
3. 南方海洋科学与工程广东省实验室(广州), 广东 广州 511458
中国南海珊瑚礁海域的分布面积约有37200km2(赵焕庭 等, 2016), 其中热带珊瑚礁数量约占世界热带珊瑚礁总数量的7.274%(王耕 等, 2019)。南海海域具有相对丰富的珊瑚礁鱼类资源, 当前所记录的珊瑚礁鱼类物种超过900种(李永振 等, 2007; 邵广昭等, 2011; 高永利 等, 2014)。近年来, 在人类活动及全球气候变化的影响下, 南海珊瑚礁渔业资源已出现严重衰退的现象(傅秀梅 等, 2009; 张俊 等,2020)。
黑缘尾九棘鲈(Cephalopholis spiloparaea)是南海珊瑚礁渔业捕捞活动的主要目标物种之一, 隶属于鲈形目(Perciformes)鮨科(Serranidae)九棘鲈属(Cephalopholis), 为小型珊瑚礁鱼类(体长<30cm)。在西起东非沿岸、东至法属波利尼西亚、北达日本琉球群岛、南至澳大利亚大堡礁(Great Barrier Reef)南端的海域中广泛分布(Randall, 1987), 普遍栖息于水深15~30m的珊瑚礁外缘礁坡之上(Myers, 1991;Donaldson, 1995a, 2002), 主要以底栖甲壳类动物为食(Donaldson, 1995a)。此前已有国外学者对黑缘尾九棘鲈的生境特征、繁殖特性、觅食活动和社交行为等进行了研究(Shpigel et al, 1991; Donaldson,1995a, b, 2002), 但对于其地理纬度的群体基础生物学比较研究仍相对缺乏。本文利用2018年4月、9月和2019年5月、6月于南海南沙群岛和西沙群岛两个珊瑚礁海域采集的黑缘尾九棘鲈样本, 对两海域群体的体长、体质量、性比与性成熟率、性腺成熟度、摄食等级、肥满度、肝体比, 以及年龄和生长等生物学特征进行研究和分析, 比较两海域间黑缘尾九棘鲈群体生物学特征的差异, 并探讨其生物学特征变化与地理环境差异的潜在联系, 旨在为南海小型珊瑚礁鱼类资源的保护和可持续利用提供基础资料和科学依据。
1 材料与方法
1.1 样本采集
2018年4月、9月(简称2018年样本)和2019年5月、6月(简称2019年样本)在南海南沙和西沙群岛海域采集黑缘尾九棘鲈样本共362尾(表1)。调查船为“南锋”号(满载排水量1980t, 主机总功率1920kW)和船载工作艇(总吨位 1t, 主机功率14.7kW)。样本采集方式为手钓和地笼网, 钓具的钓钩尺寸为25mm×8mm, 钓线直径为0.33mm; 地笼网的长、宽、高分别为100cm、30cm、30cm; 作业深度为10~30m; 采样时间段分别为6:00—12:00、12:00—18:00和18:00—24:00。

表1 南海南沙群岛和西沙群岛海域黑缘尾九棘鲈样本采集信息Tab. 1 Sampling information of C. spiloparaea specimens from Nansha Islands and Xisha Islands waters in the South China Sea
1.2 生物学测定
现场采集的黑缘尾九棘鲈冷冻保存, 实验室解冻后进行生物学实验, 按照《海洋调查规范》(GB/T 12763.6-2007)测定体长(Standard length, SL)、体质量(Body weight, BW)、性别(Sex)、性腺成熟度(Maturity stage)和摄食等级(Feeding stage)等。其中, 体长和体质量分别精确到1mm和0.01g, 性腺成熟度的鉴定采用通用的Ⅰ~Ⅵ期的6期划分标准, 摄食等级的鉴定采用0~4级的5级划分标准(费鸿年 等, 1990)。
1.3 体长和体质量关系
根据频率分布法分别以10mm和15.00g作为黑缘尾九棘鲈体长和体质量的组距进行分组, 统计并分别绘制南沙群岛和西沙群岛海域样本的体长组和体质量组的频次分布图, 定义超过样本总尾数20%以上的体长组和体质量组作为优势体长组和优势体质量组。
利用幂函数关系式表示黑缘尾九棘鲈的体长-体质量关系:

式中:W表示体质量(单位: g);L表示体长(单位:mm); 参数a、b作为常数分别表示条件因子和异速生长因子。当b=3时, 表示鱼类等速生长; 当b<3时, 表示鱼类处于负异速生长状态; 当b>3时, 表示鱼类处于正异速生长状态。
利用对数转换的体长-体质量关系方程(殷名称,1995)及最小二乘法(Least square method, LSM)求解lna和参数b:

利用lna对数变换得到的lga与b进行线性回归拟合并绘制相应散点图, 以检验参数a、b是否为异常值, 若点偏离回归直线超过2倍标准差即可认为该点所对应的体长-体质量关系异常(张俊 等,2020)。
1.4 性比与性腺成熟度
分析南沙和西沙海域黑缘尾九棘鲈的性比和性腺成熟度在不同体长组间的差异, 并利用如下公式计算各群体的性成熟率:

1.5 摄食等级、肥满度与肝体比
分析2018—2019年南沙和西沙海域黑缘尾九棘鲈摄食等级在各体长组间的差异, 并利用如下公式计算相关群体的空胃率(Percentage of empty stomach, PES):

式中:Ne和Nt分别为空胃样本数和样本总数。
肥满度(Badiani et al, 1997)作为一种表征动物的生理或营养状态的形态生理指数(戴强 等, 2006),在鱼类生物学特征研究中被广泛应用, 以反映鱼类的生长状态。利用Le Cren相对状态指数Ka(Relative condition factor)评估黑缘尾九棘鲈的肥满度(Froese,2006), 其计算公式为:

式中:W、L同式(1);am为黑缘尾九棘鲈各群体非异常a值的几何平均值;bm为各群体非异常b值的算术平均值。
数据应用SPSS18.0进行分析,其中计数进行χ2(%)检验,计量进行t检测(±s)检验,P<0.05提示有显著差异。
肝体比(Hepatopancreas index, HSI)可以反映鱼类的生存环境和生长状况, 其计算公式(Hempson et al, 2018)为:

式中:Wl和Wg分别为肝脏质量(单位: g)和净体质量(单位: g)。
1.6 年龄和生长
由于鱼类的耳石与躯体的其他硬组织相比, 具有记录鱼类生活史的连续性以及易保存性等特征,通常被用作鱼类年龄鉴定的优质材料(刘飞 等,2019)。在鱼类生物学研究中, 耳石日轮对于揭示鱼类自身生长发育和外界环境的关系具有重要的应用价值(陈新军, 2014)。本研究于2019年5—6月在南沙群岛的永暑礁和西沙群岛的琛航岛海域分别采集了黑缘尾九棘鲈耳石样本各30对。耳石样本通过固化剂和冷埋树脂包埋于模具中, 待其硬化之后利用切割机沿耳石纵切面方向切割, 并使用热熔胶将切片固定于载玻片之上。在研磨机上依次利用多种规格的防水耐磨砂纸沿耳石纵切面研磨至核心, 利用0.3μm氧化铝水绒布抛光。对耳石切片进行双面研磨和抛光, 以便精确计数耳石日轮(刘必林 等, 2016;朱庆环 等, 2020)。
使用Olympus双筒光学显微镜(CX21FS1)观察研磨成型的耳石切片, 在40倍物镜下采用DP71(显微数码相机)进行拍照, 并利用Adobe Photoshop CS6软件对图像进行叠加和拼接处理(李建华 等,2018)。耳石日轮计数时, 每一样本独立计数5次, 去除其中的最大值与最小值, 若剩余3次的观测值与均值之差均不超过均值的10%时, 将其均值作为最终读数(刘必林 等, 2009; 张国政 等, 2010)。
根据黑缘尾九棘鲈耳石样本的日轮数目和相应的体长数据, 采用von Bertalanffy生长方程拟合黑缘尾九棘鲈的体长-年龄关系(詹秉义, 2005):
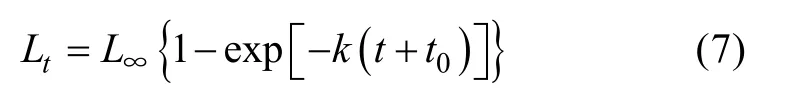
式中:tL代表年龄为t时的体长;L∞为渐近体长;k为生长系数;t0为理论上鱼类体长等于零时的年龄。
1.7 数据分析
2 结果与分析
2.1 群体结构
2018—2019年于南海南沙和西沙珊瑚礁海域采集的黑缘尾九棘鲈样本的体长和体质量分布均呈明显的单峰分布(图 1, 表 2); 单样本的Kolmogorov-Smirnov检验结果显示, 有且仅有西沙海域群体的体质量分布不符合正态分布(P<0.05)。通过非参数检验分析, 结果显示南沙海域和西沙海域间的黑缘尾九棘鲈在体长和体质量上均具有极显著的差异(P<0.01); 单因素方差分析结果表明, 两海域黑缘尾九棘鲈雌性和雄性间样本的体长均值具有显著性差异(P<0.05), 西沙海域群体雌、雄间的体质量均值也具有显著性差异(P<0.05)。

表2 2018—2019年南沙和西沙海域黑缘尾九棘鲈的体长、体质量信息Tab. 2 Information of body length and body weight of C. spiloparaea in Nansha and Xisha waters in 2018 and 2019
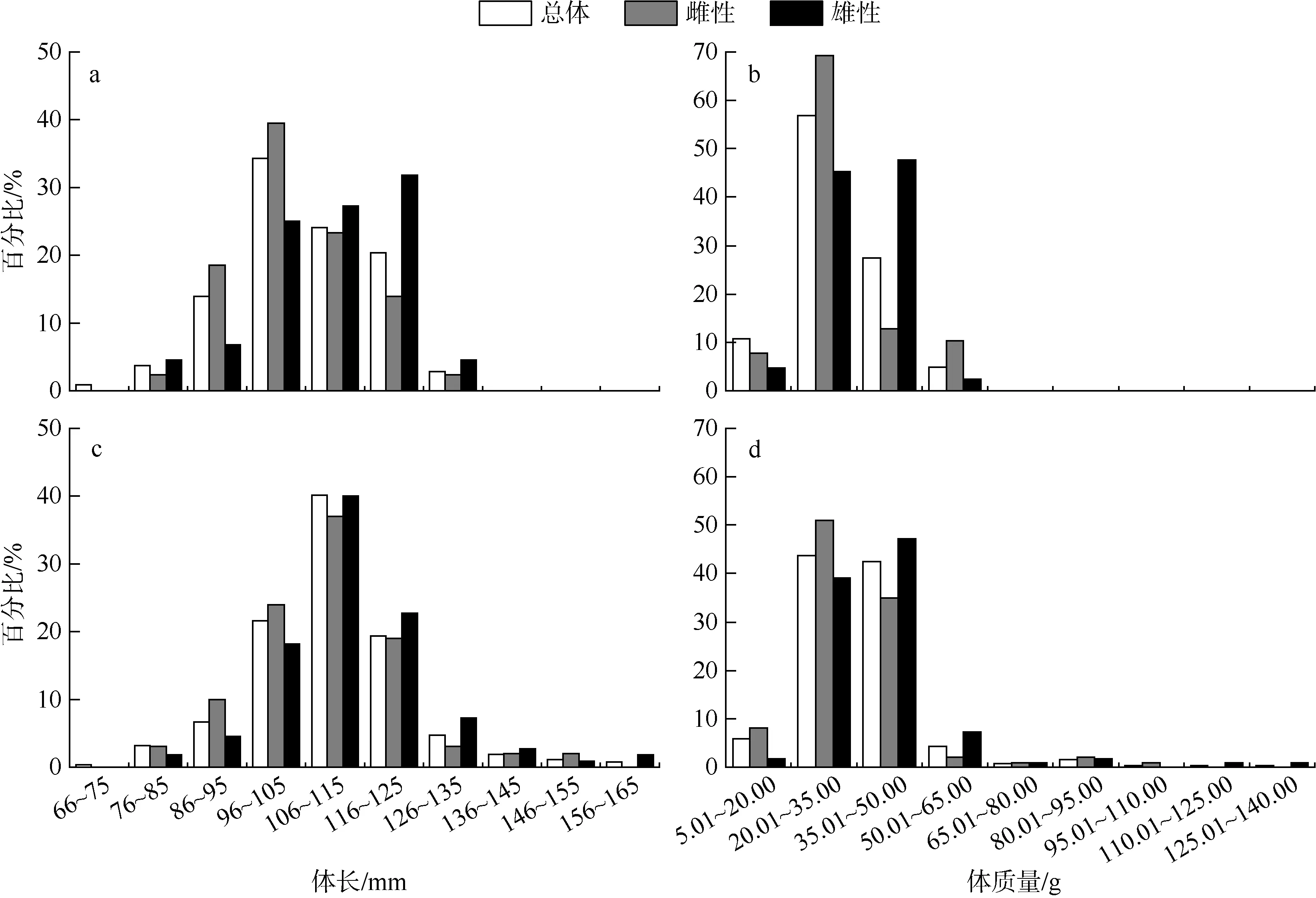
图1 黑缘尾九棘鲈的体长、体质量分布a、b: 南沙海域; c、d: 西沙海域Fig.1 Distributions of body length and body weight of C. spiloparaea
2.2 体长与体质量关系
协方差分析检验结果表明黑缘尾九棘鲈样本的体长-体质量关系在雌、雄之间差异不显著(P>0.05),其体长-体质量关系参数如表3所示。在南沙和西沙海域中, 各黑缘尾九棘鲈群体均为负异速生长(b<3);雌性群体b值范围为2.789~2.851, 雄性群体b值范围为2.735~2.801。通过对黑缘尾九棘鲈的体长-体质量关系参数lga与b进行线性关系检验, 结果显示lga与b值呈线性负相关关系(斜率为-2.002), 且各群体的参数a、b均不存在异常值。

表3 黑缘尾九棘鲈的体长-体质量关系参数Tab. 3 Estimated parameters of body length vs body weight of C. spiloparaea in Nansha and Xisha Islands waters
2.3 性比与性腺成熟度
在本研究所采集的黑缘尾九棘鲈样本中, 共有359尾进行了性别和性腺成熟度检测, 其中雌性和雄性尾数分别为143尾和154尾, 未能分辨性别的样本为62尾。南沙海域和西沙海域黑缘尾九棘鲈的雌雄性比分别为0.91和0.97, 卡方检验分析显示两海域黑缘尾九棘鲈的雌雄性比均符合1:1(P>0.05)。非参数检验分析显示南沙海域群体在各体长组间的性比分布差异极显著(P<0.01), 而西沙海域群体则无显著性差异(P>0.05)(图2)。南沙和西沙海域黑缘尾九棘鲈雄性个体在各体长组的占比均基本随体长的增长而增加。
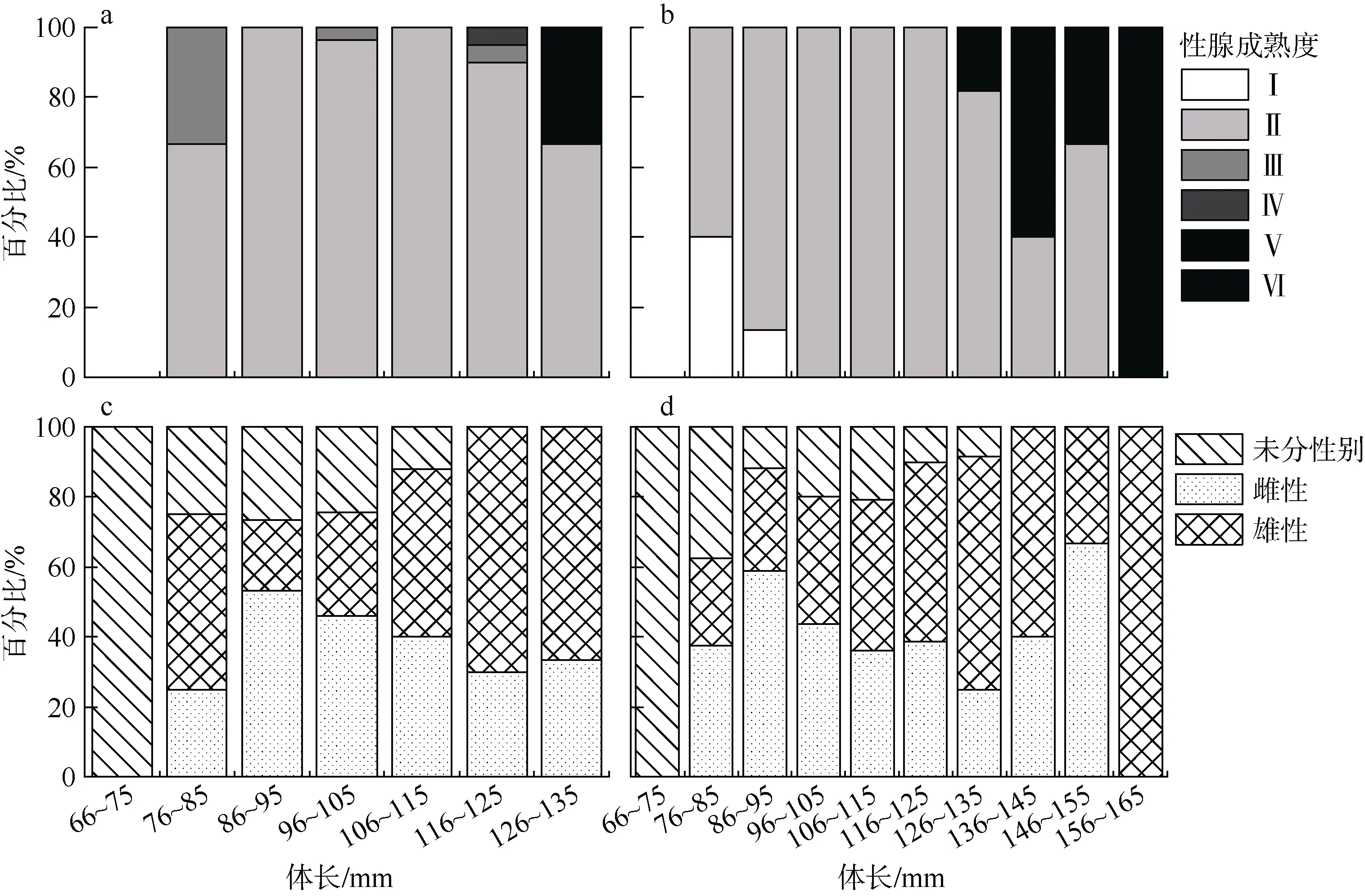
图2 黑缘尾九棘鲈不同体长组的性腺成熟度和性别分布a、c: 南沙海域; b、d: 西沙海域Fig.2 Distributions of gonad maturity stages and sexes of C. spiloparaea with different body length classes
根据性腺成熟度的鉴定分析结果, 黑缘尾九棘鲈的性腺成熟度包括Ⅰ期、Ⅱ期、Ⅲ期、Ⅳ期和Ⅵ期, 占比分别为1.35%、94.28%、1.01%、0.34%和3.03%, 调查期间未发现有Ⅴ期性腺成熟度的样本。南沙和西沙海域间黑缘尾九棘鲈的性腺成熟度分布无显著性差异(P>0.05), 且均以Ⅱ期为主。南沙海域和西沙海域黑缘尾九棘鲈群体的性成熟率分别为1.91%和3.15%。
2.4 摄食等级
2018—2019年南沙和西沙海域的黑缘尾九棘鲈摄食等级包括0~4级, 其中南沙海域群体各体长组中的平均摄食等级整体上随体长的增大而上升(图3),西沙海域的黑缘尾九棘鲈群体则例外。南沙和西沙群体的摄食等级均以1级为主, 分别占49.00%和51.90%, 两海域间各摄食等级占比的分布无显著性差异(P>0.05); 南沙和西沙黑缘尾九棘鲈的空胃(摄食等级为0级)率分别为39.00%和37.98%, 两海域样本的平均摄食等级分别为0.82和0.75。南沙和西沙海域各群体的摄食等级在体长组间的分布均无显著性差异(P>0.05)。

图3 黑缘尾九棘鲈不同体长组的摄食等级百分比a. 南沙海域; b. 西沙海域Fig.3 Percentage of different feeding stages of C. spiloparaea with different body length classes
黑缘尾九棘鲈的平均摄食等级随采样时间的不同而具有一定的差异(图4)。在上午(6:00~12:00)、下午(12:00~18:00)和夜晚(18:00~24:00)三个时间段中, 南沙海域群体的平均摄食等级分别在上午和夜晚出现极小值(0.49)和极大值(1.13), 西沙海域群体的平均摄食等级则分别在下午和上午出现极小值(0.40)和极大值(0.82) 。根据非参数检验和单因素方差检验分析, 南沙海域黑缘尾九棘鲈在各时间段中体长的分布与体长均值均无显著性差异(P>0.05),而西沙海域群体在各时间段中体长的分布和体长均值则均具有极显著的差异(P<0.01) (图5)。
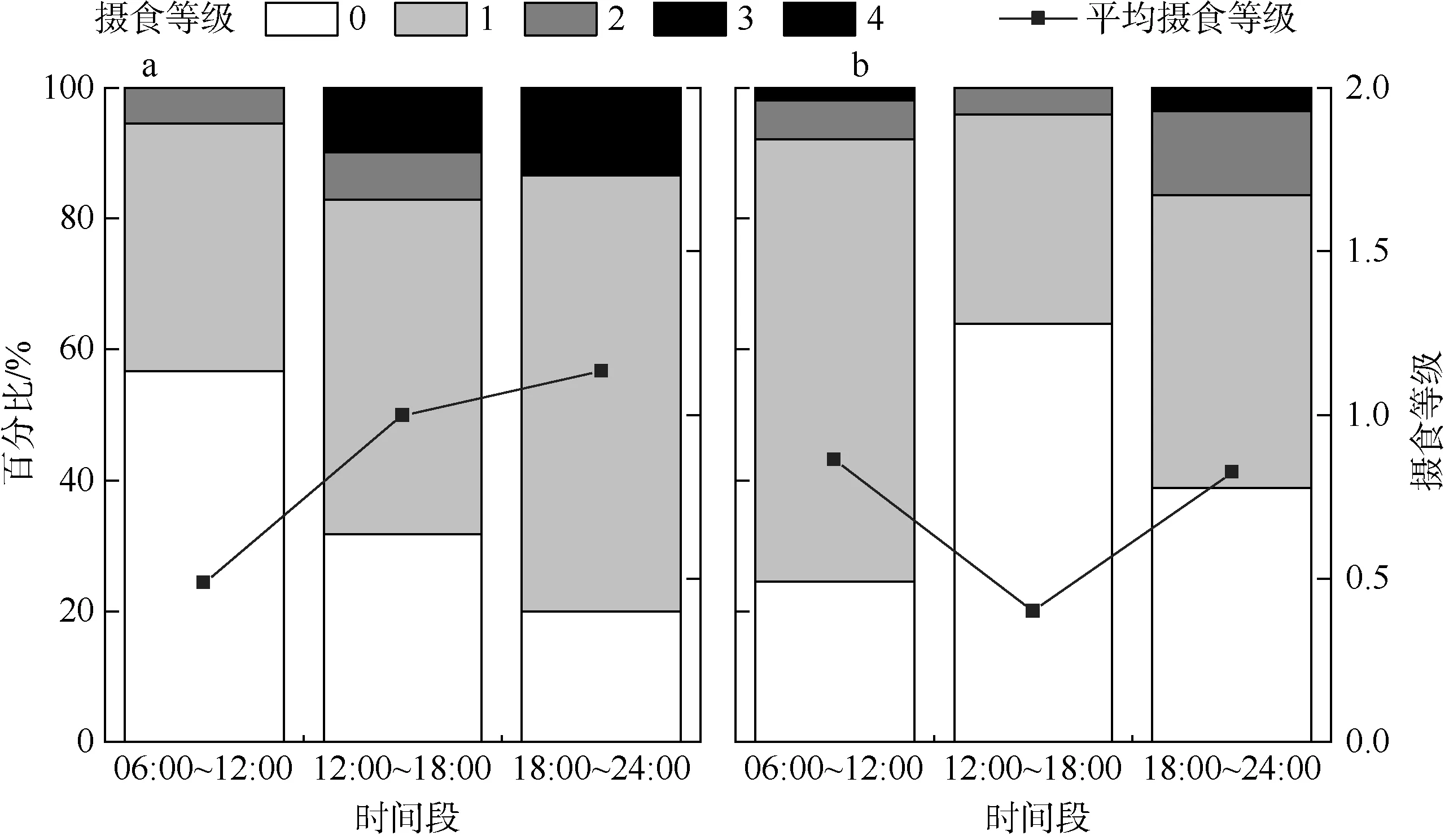
图4 黑缘尾九棘鲈不同时间段的摄食等级百分比a. 南沙海域; b. 西沙海域Fig.4 Percentage of different feeding stages of C. spiloparaeaat different times: a) samples from Nansha waters; b) samples from Xisha waters
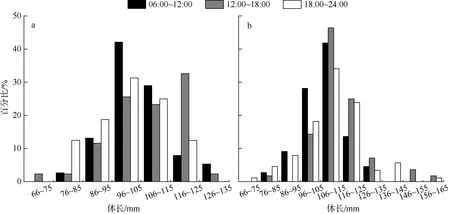
图5 黑缘尾九棘鲈不同时间段的体长分布a. 南沙海域; b. 西沙海域Fig.5 Distribution of body length of C. spiloparaea at different times: a) samples from Nansha waters; b) samples from Xisha waters
2.5 肥满度和肝体比
根据上文计算得出的各群体的参数a、b值, 可推算得到其相应的几何平均值am和bm分别为6.514×10-5和2.8040, 最终获得各群体的肥满度分布状态如图6a所示。南沙海域和西沙海域黑缘尾九棘鲈的平均肥满度分别为100.85%和99.71%, 两海域间的肥满度分布无显著性差异(P>0.05)。两海域间雌性黑缘尾九棘鲈的肥满度分布与均值均具有极显著的差异(P<0.01),而雄性的则均无显著性差异(P>0.05)。
南沙海域和西沙海域黑缘尾九棘鲈的平均肝体比分别为0.54%和0.78%(图6b), 两海域间的肝体比分布存在极显著性差异(P<0.01)。西沙海域黑缘尾九棘鲈的平均肝体比高于南沙海域, 且西沙海域雌性群体的平均肝体比低于雄性。

图6 南沙和西沙海域黑缘尾九棘鲈的肥满度和肝体比分布箱线图a. 肥满度分布; b. 肝体比分布Fig.6 Boxplot of condition factor and hepatosomatic index of C. spiloparaea in each reef areas: a) distribution of condition factor, and b) distribution of hepatosomatic index
2.6 年龄与生长
本研究选取60对黑缘尾九棘鲈耳石样本, 其中南沙海域(永暑礁)和西沙海域(琛航岛)的样本分别30对。南沙海域礁区黑缘尾九棘鲈的年龄范围为0.66~1.44a, 平均年龄为0.93±0.19a, 最小年龄与最大年龄所对应的体长分别为95mm和134mm; 西沙海域礁区黑缘尾九棘鲈的年龄范围为0.58~1.39a,平均年龄为1.01±0.24a, 最小年龄与最大年龄所对应的体长分别为87mm和123mm。两海域样本的平均年龄为0.97±0.22a, 不同海域间样本的年龄分布并无显著性差异(P>0.05)。黑缘尾九棘鲈的体长-年龄关系适合von Bertalanffy模型(图7)(费鸿年 等,1990)。von Bertalanffy方程的拟合结果显示, 南沙海域礁区和西沙海域礁区黑缘尾九棘鲈的渐近体长(L∞)分别为185.24mm和195.44mm(表4), 生长系数(k)分别为0.69a-1和0.42a-1, 满足 e-k( 0.458 ) < 1, 表明k值估算准确。

图7 黑缘尾九棘鲈的体长-年龄关系a. 永暑礁(2019年5月); b. 琛航岛(2019年6月); c. 总体Fig.7 Relationship between body length and age of C. spiloparaea

表4 南沙和西沙海域黑缘尾九棘鲈的生长参数Tab. 4 Growth parameters of C. spiloparaea in reef areas of the South China Sea
3 讨论
3.1 南海海域黑缘尾九棘鲈的生长特征
鱼类的体长-体质量关系式中的异速生长因子b值可用于判断鱼类种群的生长发育速度是否均匀,其数值也会受到食物组成、摄食强度、性腺成熟度等因素的影响(李忠炉 等, 2011)。从南沙和西沙海域各群体黑缘尾九棘鲈的体长-体质量关系式可以看出,b值范围为2.744~2.881, 均小于3, 为负异速生长, 这与陈国宝等(2005)研究发现南海海域的主要鮨 科鱼类异速生长因子b值范围为2.7~3.0的结果相一致。雌性黑缘尾九棘鲈在南沙海域与西沙海域中的b值均略高于雄性, 而雄性的环境因子a值则明显高于雌性(江艳娥 等, 2019), 这可能反映了相同环境压力下雌性相较于雄性会投入更多资源用于自身的生长和发育(Trippel, 1995)。当前多个珊瑚礁海域均有过度捕捞现象的出现(Previero et al, 2018),捕捞压力的增加会使鱼类为适应捕捞而改变其生长和发育策略(Silva et al, 2013)。因此, 造成南海岛礁海域黑缘尾九棘鲈a值和b值在雌、雄之间具有相同差异特征的原因, 可能与鱼类为适应捕捞而投入更多能量用于性发育以及繁殖活动有关(Conover et al, 2002)。
3.2 南海海域黑缘尾九棘鲈的性成熟发育
黑缘尾九棘鲈的性腺成熟度以Ⅱ期为绝对优势,这与南海海域中其他礁栖鱼类如金带齿颌鲷(Gnathodentex aurolineatus)(张俊 等, 2015)、尾纹九棘鲈(Cephalopholis urodelus)和六角石斑鱼(Epinephe lus hexagonatus)(陈国宝 等, 2005)等的调查结果相似。另外, 黑缘尾九棘鲈性成熟个体在各体长组中的比例基本随体长组等级的增大而增加,即成正比例关系, 符合鱼类正常的生长发育形式(郜星晨 等, 2019)。但南沙海域中黑缘尾九棘鲈雌性性成熟个体的最小体长(124mm)显著小于西沙海域雌性性成熟个体的最小体长(134mm), 这可能反映了南沙海域黑缘尾九棘鲈群体较西沙海域群体具有更快的性腺发育速度。
调查发现, 南海海域黑缘尾九棘鲈的雄性个体数略高于雌性, 雌性与雄性在体型上出现明显的二态性, 雄性的平均体长与平均体质量均高于雌性;各海域间的雌、雄性比不存在差异, 各体长组间的雌、雄性比大致随体长组等级的增大而减小,Donaldson (1995b)认为黑缘尾九棘鲈具有雌雄同体的发育状态, 且可能雌性性腺的成熟较早发生。因此, 根据这个现象, 本研究推测黑缘尾九棘鲈的性腺发育速度在雌性和雄性之间存在差异。此外, 根据“一雄多雌”制的种群社会结构, 不排除在某些特定情况下黑缘尾九棘鲈可能也具有其他鮨科鱼类所表现的性逆转现象(陈国宝 等, 2005)。根据本文研究所知黑缘尾九棘鲈雌雄性比随体长的增大而减小,推测其部分雌性个体体型生长达到一定尺寸之后通过性逆转转化为雄性个体, 这种雌性先熟的发育特征在大部分珊瑚礁鱼类中均有出现(Warner, 1970)。
3.3 生长发育
根据von Bertalanffy方程估算出南沙海域和西沙海域黑缘尾九棘鲈的生长系数(k)分别为0.69a-1和 0.42a-1, 均表现为快速生长类型(k>0.2a-1)(Branstetter, 1987), 这可能表明黑缘尾九棘鲈在南海珊瑚礁海域普遍具有较高的生长速度。此外, 南沙海域黑缘尾九棘鲈的渐近体长明显小于西沙海域,结合上述两者k值的大小比较, 推测南沙海域的黑缘尾九棘鲈比西沙海域体型相对较小、生长速度相对较快。其中的影响因素可能包括不同纬度下海表温度(Sea surface temperature, SST)的差异(贾丹丹,2018), 较高的水温会促使鱼类新陈代谢加速, 从而导致鱼类生长速度加快(庞旭, 2014)。此外, 南沙海域黑缘尾九棘鲈体型的小型化趋势也可能是由于人类活动所造成的环境胁迫的结果, 过度捕捞所导致的鱼类小型化现象已在多数近海经济鱼类种群中出现(张魁 等, 2016; 王继隆 等, 2020), 张俊等(2020)在南海礁栖鱼类四带笛鲷(Lutjanus kasmira)的生物学研究中也发现了这一现象。
研究显示, 捕捞会诱使鱼类的生物学特征产生非自然变化, 捕捞诱导进化(Fishing-induced evolution,FIE)(Jørgensen et al, 2007)促使鱼类个体变小且性成熟提前(单秀娟 等, 2020), 而环境变化因素也会对鱼类生长产生关键性的影响(Pérez-Rodríguez et al,2013)。本文研究结果显示, 南海黑缘尾九棘鲈的生长发育存在明显的地区差异, 且南沙海域群体的性腺发育较早成熟, 这可能与其所栖息海域纬度更低有关, 也可能是该鱼类为适应捕捞压力增大和礁栖环境恶化(龙丽娟 等, 2019)等外界因素而提前性成熟的结果(Heino et al, 2015)。
3.4 摄食等级肥满度与肝体比
摄食活动作为珊瑚礁鱼类群落构建的重要组成部分, 是维持种群和生态平衡的基础之一(Hixon,1991)。本研究发现, 南海各海域黑缘九棘鲈的摄食等级分布在相同年份出现高度的一致性, 南沙和西沙海域群体均以1级为主, 两海域中各体长组间的平均摄食等级差异并不显著, 这些现象可能表明黑缘尾九棘鲈在南沙和西沙礁栖环境中的摄食条件是相似的。Donaldson(2002)发现黑缘尾九棘鲈的分布主要与火焰滨珊瑚(Porites rus)的覆盖和增殖密切相关, 因而具有特殊的生境偏好性。但人类活动所导致的环境变化也可能会促使鱼类的觅食等行为产生可塑性变化, 以应对其生活史中的环境变化(Karkarey et al, 2017)。虽然过去记录显示黑缘尾九棘鲈具有以甲壳类为主的食性, 但在大多数鱼类体内, 鱼类被消化的速度约为甲壳类的 4倍(Beukers-Stewart et al, 2004 )。因此, 南海海域黑缘尾九棘鲈的索饵行为是否随珊瑚礁生态环境变化(李淑 等, 2011; 贾丹丹, 2018; 李娇 等, 2020)而发生了某种程度的转化, 从而导致其平均摄食等级偏低, 还有待于通过对其食性开展进一步研究来验证。
本研究中, 南沙和西沙海域黑缘尾九棘鲈群体平均摄食等级的极小值均出现在白昼期间, 且在夜晚时间段中高胃充塞度(摄食等级≥3)的群体占比均高于其他时间段, 这印证了夜间可能是黑缘尾九棘鲈的主要觅食时间之一(Donaldson, 1995b)。此外,鱼类的摄食不仅受外界环境条件影响, 同时也受自身内源性的影响(李忠义 等, 2018; 许莉莉 等,2018)。本研究中各采集时间段间黑缘尾九棘鲈的体长分布, 有且仅有西沙海域群体具有显著性差异,夜晚和下午时间段的平均体长均高于上午时间段的平均体长; 西沙海域群体在上午时间段所出现的优势体长组为96~115mm, 小于下午和夜晚时间段出现的优势体长组(106~125mm)范围。这显示黑缘尾九棘鲈的觅食模式与其生活史阶段的变化密切相关,即大体型个体偏好在夜晚进行觅食活动, 中小体型个体为满足自身生长发育所需要的更多物质和能量而进行全天候觅食。
在体长一致的前提下, 鱼类个体体质量相对越大则肥满度越高, 显示个体的生理和营养状态越好(Jones et al, 1999; 曹希全 等, 2019)。而鱼类体内的物质和能量在个体性成熟之前主要用于鱼体的生长,但在性成熟之后还须用以支持性腺发育(杨骏 等,2020)。本研究中, 西沙海域黑缘尾九棘鲈群体的平均肥满度整体略低于南沙海域群体, 这表明黑缘尾九棘鲈的性腺发育会消耗自身储存的物质和能量,群体的性成熟率是造成不同海域群体间肥满度差异的主要原因(陈新军, 2014)。
肝体比在鱼类的生长发育研究中同样可以作为反映生存环境和生长状况的指标(区又君 等,2006)。本研究发现2018年西沙海域黑缘尾九棘鲈的平均肝体比高于南沙海域群体, 相同时期中前者的平均肥满度也高于后者, 表明本研究中肝体比和肥满度对于黑缘尾九棘鲈各群体生长状态的描述具有相对统一性, 这也印证了黑缘尾九棘鲈雌性群体在不同地区中采取的生存策略可能具有一定的差异性, 生境退化(Hempson et al, 2018)和食物短缺(王婷等, 2015)等原因均会导致鱼类通过消化肝脏内储存的物质以维持正常的生理活动(谢小军 等, 1998;张辉 等, 2020)。此外, 肝体比也受性腺发育的影响,鱼类在肝脏中合成卵巢发育后期所需的各类营养物质将导致肝体比的下降(韦众 等, 1999; 朱玲 等,2002)。此外,南海黑缘尾九棘鲈的雌性群体在南沙的平均肝体比则明显低于其在西沙的肝体比, 这表明肝体比与性腺的提前发育具有高度相关的关系;在环境变化和捕捞等因素所导致的生存压力增大的情况下, 南沙海域的雌性黑缘尾九棘鲈通过将更多的能量和物质投入到性腺发育中, 有助于缩减繁殖周期, 从而增加种群补充量以维持种群规模。
4 结论
本文通过对2018年4月—2019年6月在南海南沙海域和西沙海域采集的362尾黑缘尾九棘鲈进行生物学特征研究, 发现南沙海域群体相较于西沙海域群体具有体型小以及性成熟早等特征。两海域群体的异速生长因子存在雌雄性别差异, 但均在2.7~2.9的范围内。黑缘尾九棘鲈的摄食状态与生活史密切相关, 夜晚时间段是其主要的觅食活动时间之一。其肥满度和肝体比的空间差异可能是由于南沙海域雌性黑缘尾九棘鲈的生长和发育速度快于西沙海域所导致。
