Leaf Photosynthetic Characteristics of High-Yield Soybean [Glycine max (L.) Merr.] Under Different Planting Densities, Different Fertilization Rates, and Single/Mixed Planting Patterns
2021-08-02ZhigangLIXueyanBAOXingdongYAO
Zhigang LI Xueyan BAO Xingdong YAO


Abstract Understanding the photosynthetic characteristics of high-yield soybean [Glycine max (L.) Merr] cultivar (HYC) would aid research aiming at investigating the soybean high yield formation mechanism and optimization of cultivation system. To assess the photosynthesis of HYC, a pot experiment was conducted to quantify the differences in photosynthetic characteristics between HYC and common-yield soybean cultivar (CC) under different planting densities, fertilization rates, and single/mixed planting patterns. The leaf greenness (Lg), net photosynthetic rate (Ph), stomatal conductance (St) and transpiration rate (Tr) were significantly higher in HYC than CC mainly in seed-filling stages. HYC was more tolerant to dense and mixed planting because the decreases of Ph and St under high planting density and those of Ph, St, and Tr under mixed planting were lower in HYC than CC. The Lg and Ph in HYC were more superior to those in CC at high fertilization rate. Thus, the HYC has a superior performance in photosynthetic characteristics under the varied cultivation practices, which may contribute to the greater seed yield in HYC than CC.
Key words High-yield soybean; Photosynthesis; Planting density; Fertilizing; Planting pattern
Received: January 29, 2021 Accepted: April 15, 2021
Supported by National Natural Science Foundation of China (31260310); Science and Technology Reserve Project of Inner Mongolia Autonomous Region (2018MDCB02).
Zhigang LI (1970-), male, P. R. China, professor, devoted to research about crop cultivation and breeding, agricultural ecology.
*Corresponding author. E-mail: baoxueyan@imun.edu.cn.
Positive correlation is commonly found between the leaf or community photosynthetic capacity and crop production[1-2] because approximately 90%-95% of the organic substrate in the dry matter that forms the crop yield comes from leaf photosynthesis[3]. In the soybean growing region of northeast China, the target soybean yield for breeders is 4 875 kg/hm2 and soybean cultivars with seed yields that exceed this goal are known as high-yield cultivars (HYC)[4]. This goal has often been achieved by using high yield potential soybean and the most efficient cultivation practices, e.g., fertilization, irrigation, and planting patterns[5]. However, the same high-yield record might not be achieved again when the same cultivar and practices are adopted in other regions[6], partly due to the regional differences in environmental conditions, e.g., light, water, and temperature, but mainly because of a poor understanding of the physiological photosynthetic parameters of HYC soybean and their differences from common soybean cultivars (CC)[4]. Therefore, analyzing the photosynthetic characteristics of HYC is important for exploring the high-yield mechanism and further enhancing the seed yield in soybean.
Several studies have investigated the photosynthetic characteristics of HYC. In particular, Zhang et al.[7] and Wei et al.[6] concluded that the leaf greenness (Lg), net photosynthesis, duration of the maximum leaf area, and photosynthetic potential of HYC were superior to a common cultivar (CC) during the late growing stage. However, these studies were all conducted under a single cultivation practice, thereby failing to improve our understanding of the photosynthetic characteristics of HYC.
Manipulating the planting density and fertilization rate are the two most important and common cultivation practices applied in soybean fields[8-10]. The planting density can strongly affect the leaf area, light interception, and canopy photosynthesis in soybean[11-13]. An appropriate planting density can enhance the canopy light interception to increase photosynthesis in the canopy and the seed yield[14-15]. Supplying nitrogen can have positive effects on plant growth, mainly by stimulating photosynthesis[16]. About 70% of the nitrogen in plant leaves is present in the chloroplasts and most is used to synthesize the photosynthetic apparatus[17-18]. Therefore, fertilization has important effects on leaf photosynthesis[19-21]. Previous studies of HYC under different planting density levels and fertilization rates investigated their characteristics in terms of nutrient uptake[4], nutrient accumulation[22], root morphology or activity[23], and root nodules[24]. The results suggested that certain advantages of these traits are beneficial for high yield formation compared with CC, but the photosynthetic characteristics of HYC cultivars and their differences compared with CC under cultivation practices at different levels are still unknown.
An efficient planting pattern is important for improving photosynthesis and the seed yield in soybean[25-26]. Previous studies have investigated the effects of different row configurations for the same soybean cultivar[27] or intercropping with two or more crops in narrow to wide rows[28-30] on the physiology of photosynthesis, but the photosynthetic characteristics under mixed planting patterns have not yet been studied in soybean. In the present study, we determined the photosynthetic characteristics of a HYC Liaodou 14, and a CC Liaodou 11 under different planting densities, fertilization rates, and planting patterns. Our objective was to explore the differences in the photosynthetic characteristics of soybean cultivars with two yield levels under different cultivation practices. Our findings will help to understand the photosynthetic traits of HYC soybean and they provide a useful reference to facilitate efficient breeding and optimizing soybean cultivation practices.
Materials and Methods
Plant materials and experimental design
This study used a HYC Liaodou 14 with high yield records in soybean production, and a CC Liaodou 11. These soybean cultivars were provided by Agricultural Collage, Shenyang Agricultural University.
The experiment was conducted in pots at the experiment farmland of Shenyang Agricultural University during 2012 and 2013. The soil used in the experiment was brown loam containing 70.2 mg/kg alkali-hydrolyzed nitrogen, 21.5 mg/kg available phosphorus, and 138 mg/kg available potassium. Each pot measured 25 cm×20 cm×30 cm (height bottom diameter top diameter) in size and contained 12.5 kg of soil. Three planting densities (D1, two plants per pot, D2, four plants per pot, and D3 six plants per pot with no fertilizer and single planting), three fertilization rates (F1, 0 mg diammonium phosphate per kg of soil, F2, 100 mg diammonium phosphate per kg of soil, and F3, 200 mg diammonium phosphate per kg of soil at a planting density of four plants per pot and single planting), and two planting patterns (single planting, S: each pot contained a single cultivar; and mixed planting, M: four cultivars planted at a ratio of 1∶1 and with a planting density of four plants per pot and no fertilizer) were tested to assess the leaf photosynthesis characteristics of HYC and CC. The experiments were arranged in a completely randomized block design with three replicates, i.e., three pots per each treatment.
Environmental information
Mean air temperature was 7.8 and 8.9 ℃ in 2012 and 2013, respectively. The annual rainfall was 445 and 475 mm, respectively. Additional meteorological information is given in Table 1, which was obtained from China Meteorological Administration.
Relevant measurements
The soybeans were sown in May and harvested during October in 2012 and 2013. A chlorophyll meter (SPAD-502) was used to measure the Lg of the third fully expanded leaves from the top of each sample soybean plant per each pot during the flowering stage (FL), mid-seed filling stage (MS), and late seed filling stage (LS). Moreover, the net photosynthetic rate (Ph), stomatal conductance (St), and transpiration rate (Tr) were measured for the same leaf used to obtain Lg measurements with a Li-6400XT (Li-Cor, United States). Since the leaf photosynthetic rate usually present remarkable daily variation, in order to minimize the error caused by the change of light condition, all the measurement conducted within 1 h (10:00-11:00 am) on a clear day. At full maturity stage (R8), plants of three pots per each treatment were taken for seed yield measurement, and the yields were adjusted to 132 g/kg moisture content.
Data analysis
The variance of soybean photosynthetic traits between the two years may result from the combined effects of environment and genotype. A method of best linear unbiased predictors (BLUP) was used to predict the true genotypic effect of each soybean cultivar regardless of environmental effect. It can provide a sufficient estimate of cultivar effect across different environments. Best linear unbiased predictors for each cultivar were obtained using estimate statements in a model that included environment, cultivar, and cultivar×environment interaction as a random effect. For each trait, variance components were determined by fitting a linear mixed-effects model using the lmer function in the lme4 package in R (R Development Core Team, 2014). Parameter estimates for the model were obtained using the restricted maximum-likelihood method. The analysis was implemented in Graphpad Prism version 5.00 for Windows. The differences between cultivars and treatments were analyzed by variance in the general linear model using SPSS 19.0 (SPSS, Inc., Chicago. USA). Probability values of less than 0.05 were considered significant. The least significant difference test was used to separate means when treatments were significant.
Results
Analysis of environmental effects
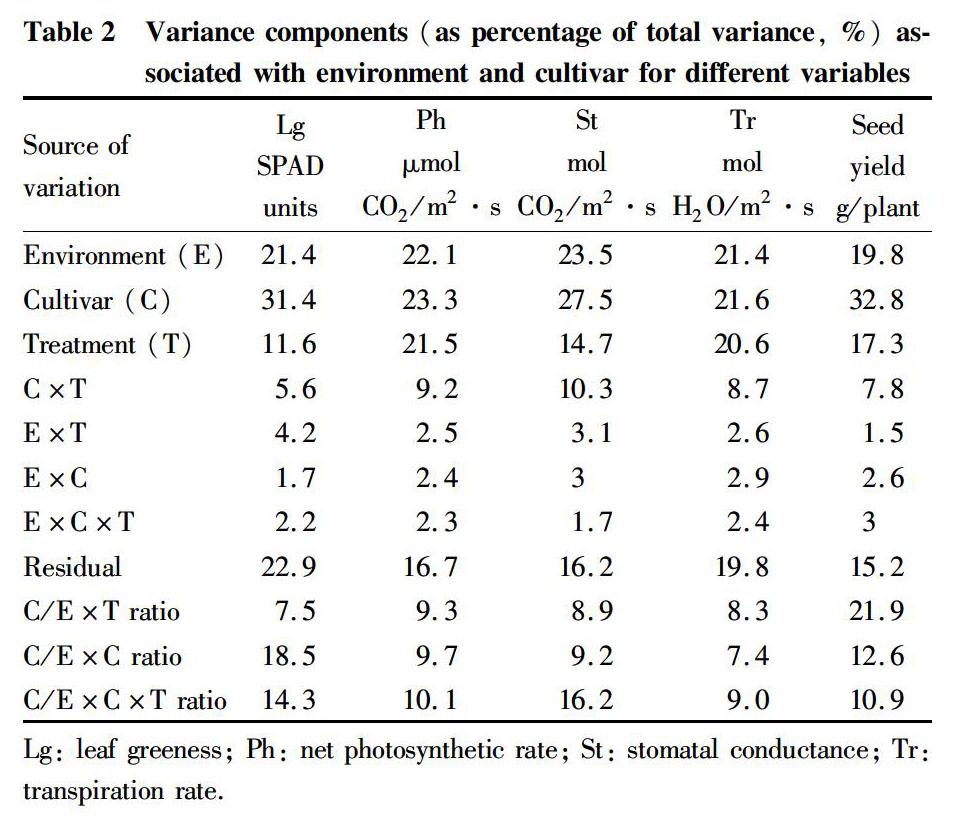
Table 2 shows that environment accounted for 19.8%-23.5% of the total variation in photosynthetic traits and seed yield, while the variance due to cultivar ranged from 21.6% to 32.8%. Cultivar explained 7.5-21.9 times more variation than the environment×cultivar, environment×treatment, and environment×cultivar×treatment interaction, indicating high consistency of cultivar behavior across environments. Therefore, we used mean values for photosynthetic traits and seed yield from the two years for the following discussions.
Lg in HYC and CC
Lg of HYC was significantly higher than that of CC at D1-MS (i.e. at D1 for MS stage, the same below), D2-LS, D3-MS,F1-LS, F2-LS, F3- FL, MS, and LS, S-LS, M- FL, MS, and LS, with the change magnitudes ranged from 3.1% to 14.9% (Fig. 1). Table 3 shows that as the planting density increased, the Lg in CC exhibited an obvious decreasing trend, whereas the Lg in HYC did not change significantly. Mixed planting reduced the Lg of CC, but did not affect the Lg of HYC (Table 3).
Ph in HYC and CC
Ph in HYC was significantly higher than that of CC at D1-LS, D2-LS, D3-MS and LS, F1-LS, F2-LS, F3-MS and LS, S-LS, M-LS, with the change magnitudes ranged from 10.5%-47.7% (Fig. 2). Table 3 shows that as the planting density increased, Ph in HYC at D3 was lower than that at D2, which was lower than the decrease in CC. Increasing the fertilization rate did not significantly change Ph in CC but improved Ph in HYC. Compared with F1, Ph in HYC increased by 12.73% and 15.88% at F2 and F3, respectively. Treatment M reduced the Ph values in CC and HYC, but the decrease in HYC was lower than that in CC.
The values in the table are averages based on the three measurement stages and two observation years (2012 and 2013). Lg, Ph, St and Tr indicate leaf greennesss, net photosynthetic rate, stomata conductance and transpiration rate, respectively. D1, D2, and D3 indicate two, four, and six (plants per pot), respectively, under the conditions with no fertilization and single planting. F1, F2, and F3 indicate 0, 100, and 200 (mg/kg) fertilizer, respectively, under the conditions with four (plants per pot) and single planting. S and M indicate single and mixed planting, respectively, under the conditions with four (plants per pot) and no fertilization. X (%) indicates the percentage change in different characteristics for D2, D3, versus D1, or F2, F3, versus F1, or M versus S. The uppercase and lowercase letters outside the brackets denote significant differences between different treatments at P<0.01 and 0.05, respectively. The uppercase and lowercase letters within brackets denote significant differences within the same treatment at P<0.01 and 0.05, respectively.
St in HYC and CC
St in HYC was significantly higher than that in CC at D1-LS, D2-MS, D3-LS, F1-MS, F1-LS, F2-MS, F3-MS, F3-LS, S-MS, S-LS, M-MS, M-LS, with the change magnitudes ranged from 25.11% to 83.48% (Fig. 3). As the planting density increased, St exhibited a decreasing trend in both cultivar types (Table 3), i.e. St in CC were 6.19% and 23.38% lower at treatments F2 and F3, respectively, compared with treatment F1. St in HYC was 20.43% lower at treatment F3 compared with that at treatment F1. St values in CC and HYC were significantly lower at treatment M than at treatment S.
Tr in HYC and CC
Ph in HYC was significantly higher than that of CC at D1-MS and LS, D2-LS, D3-MS and LS, F1-LS, F2-MS, F3-FL and LS, S-LS, M-MS, with the change magnitudes ranged from 14.62%-80.18% (Fig. 4). Table 3 shows that as the planting density increased, Tr in CC was significantly lower at D3 treatment than at D1 treatment, but Tr in HYC did not change significantly. Treatment M decreased Tr in both cultivars compared with S treatment, but the decrease in HYC was lower than that in CC.
D1, D2, and D3 indicate two, four, and six plants per pot, respectively, under the conditions with no fertilization and single planting. F1, F2, and F3 indicate 0, 100, and 200 mg/kg of fertilizer, respectively, under the conditions with four plants per pot and single planting. S and M indicate single and mixed planting, respectively, under the conditions with four plants per pot and no fertilization. * and ** indicate significant differences at P<0.05 and 0.01, respectively.
Fig. 1 Differences in leaf greenness (Lg) in high-yield soybean cultivars (HYC) and common soybean cultivars (CC) in the flowering stage (FL), mid-seed filling (MS), and late-seed filling (LS) under different planting densities (a), fertilization rates (b), and planting patterns (c)
D1, D2, and D3 indicate two, four, and six plants per pot, respectively, under the conditions with no fertilization and single planting. F1, F2, and F3 indicate 0, 100, and 200 mg/kg of fertilizer, respectively, under the conditions with four plants per pot and single planting. S and M indicate single and mixed planting, respectively, under the conditions with four plants per pot and no fertilization. * and ** indicate significant differences at P<0.05 and 0.01, respectively.
Fig. 2 Differences in the net photosynthetic rate (Ph) in high-yield soybean cultivars (HYC) and common soybean cultivars (CC) in the flowering stage (FL), mid-seed filling (MS), and late-seed filling (LS) under different planting densities (a), fertilization rates (b), and planting patterns (c)
D1, D2, and D3 indicate two, four, and six plants per pot, respectively, under the conditions with no fertilization and single planting. F1, F2, and F3 indicate 0, 100, and 200 mg/kg of fertilizer, respectively, under the conditions with four plants per pot and single planting. S and M indicate single and mixed planting, respectively, under the conditions with four plants per pot and no fertilization. * and ** indicate significant differences at P<0.05 and 0.01, respectively.
Fig. 3 Differences in stomatal conductance (St) in high-yield soybean cultivars (HYC) and common soybean cultivars (CC) in the flowering stage (FL), mid-seed filling (MS), and late-seed filling (LS) under different planting densities (a), fertilization rates (b), and planting patterns (c)
D1, D2, and D3 indicate two, four, and six plants per pot, respectively, under the conditions with no fertilization and single planting. F1, F2, and F3 indicate 0, 100, and 200 mg/kg of fertilizer, respectively, under the conditions with four plants per pot and single planting. S and M indicate single and mixed planting, respectively, under the conditions with four plants per pot and no fertilization. * and ** indicate significant differences at P<0.05 and 0.01, respectively.
Fig. 4 Differences in transpiration rate (Tr) in high-yield soybean cultivars (HYC) and common soybean cultivars (CC) in the flowering stage (FL), mid-seed filling (MS), and late-seed filling (LS) under different planting densities (a), fertilization rates (b), and planting patterns (c)
Relationships between seed yield and photosynthetic characteristics
The seed yield in CC was significantly positively correlated with Lg during MS (R2=0.68, P=0.049) and LS (R2=0.91, P=0.002 9), with Ph during LS (R2=0.69, P=0.041), with St during MS (R2=0.68, P=0.043) and LS (R2=0.76, P=0.024), and with Tr during MS (R2=0.66, P=0.048). The seed yield in HYC had significant positive relationships with Lg during LS (R2=0.73, P=0.029), with Ph during FL (R2=0.97, P=0.000 3), MS (R2=0.83, P=0.011), and LS (R2=0.67, P=0.047), with St during LS (R2=0.68, P=0.045), and with Tr during MS (R2=0.71, P=0.035) (Fig. 5 and Table 4).
CC indicates common yield soybean cultivar. HYC indicates high-yield soybean cultivar. Lg, Ph, St and Tr indicate leaf greennesss, net photosynthetic rate, stomata conductance and transpiration rate, respectively. FL, MS, and LS denote the flowering, mid-seed filling, and late-seed filling growth stages, respectively. R2 is the correlation coefficient.
Discussion
Differences in photosynthetic characteristics of HYC and CC
This study quantified the Lg, which is a good indicator of leaf chlorophyll concentration[31-32]. Leaf chlorophyll is the main photosynthetic pigment in green plants[31], where it is essential for the absorption, transfer, and conversion of light energy[33], and it is closely related to the leaf photosynthetic ability[34]. Leaf stomata are the main channels that allow leaves to exchange gases with the environment via transpiration[35-36]. The St value measures the extent of stomatal opening and it is inversely related to stomatal resistance[37]. A high St value indicates that a leaf can assimilate more CO2 molecules, and thus leaf photosynthesis will be improved[38]. Therefore, plant transpiration and photosynthesis are jointly controlled by the behavior of the stomata on leaves [33]. Our results showed that Lg, Ph, St, and Tr in HYC were significantly higher than CC at MS and/or LS, indicating photosynthetic abilities in HYC were superior to CC. Compared with other growing stages, seed filling stage is the most complex metabolic stage for carbon and nitrogen assimilation and transportation[34]. Therefore, we speculated that HYC has more developed abilities that are responsible for dry matter formation, accumulation and transfer, which could provide substrate, nutrients and energy for the synthesis of matters participating in photosynthesis process. More relevant measurements and analysis are needed to explore why photosynthesis traits in HYC were superior to CC mainly at seed filling rather than other stages.
Many studies have suggested that the Lg, Ph, St, and Tr values in soybean can be significantly affected by environmental conditions[11,39-40]. Our findings suggested that increasing the planting density had negative effects on Lg, Ph, St, and Tr in CC, and on Ph and St in HYC, but it had no effects on Lg and Tr in HYC. Increasing the planting density intensifies the competition among plants for water, light, and nutrients, thereby reducing the availability of photoassimilates to meet the demands of the filling seeds and for maintaining other plant structures[41-42]. Meanwhile, as the planting density increased, the decreases in Ph and St in HYC were lower than those in CC, thereby indicating that HYC was more tolerant to dense planting and it exhibited superior photosynthetic characteristics. We did not collect the leaf area data in this study, but we inferred that the tolerance to dense planting for HYC may relate to its lower leaf area index. Many studies have suggested that high-yielding soybean cultivars usually have small leaves[43-45]. For example, during generations of genetic improvement process aimed at high-yielding soybean cultivars breeding, Jin et al.[43] found that the change in leaf area index decreased with year of cultivar release. Cultivars with smaller leaf area were usually selected because after canopy closure, there was less shading of the lower leaves than upper leaves, with subsequent higher canopy photosynthetic rate and consequent high seed yield[11].
Increasing the fertilization level improved Lg but it had no effects on Ph, St, and Tr in CC, whereas it significantly improved Lg and Ph in HYC. Moreover, the increase in Lg was higher in HYC than that in CC (Table 3), thereby indicating the superior photosynthetic characteristics of HYC compared with CC. This superior performance was more obvious at the fertilization levels of D2 and D3. This result may due to the stronger ability to absorb soil nutrients for HYC roots than CC roots.
The mixed planting pattern significantly decreased Lg, Ph, St, and Tr in both cultivar types compared with single planting (Table 3), thereby indicating that mixed planting did not enhance the photosynthetic capability of soybean in pot cultivation conditions. We consider that HYC may have developed a better root system than CC and this more developed root system may have facilitated the greater absorption and utilization of limited soil nutrients and water compared with CC. Thus, the availability of nutrients and water was more restricted for CC. This restriction may have affected the production of Lg or other photosynthetic structures, as well as reducing the water content of the plant leaves, thereby leading to stomatal closure and decreases in the Ph and Tr values in CC[46-47]. The decreases in Ph, St, and Tr in HYC under the mixed planting conditions may have been related to the greater plant height in CC compared with HYC (according to measurements during the seed filling stages), and thus CC may have shaded HYC to inhibit its growth. The St value in HYC may have been decreased due to stomatal closure under low light conditions [48-49]. However, further studies are needed to understand the underlying mechanism responsible for decreasing the traits in soybean under mixed planting. The photosynthetic characteristics decreased in soybean under mixed planting but we found that the reductions were lower in HYC than those in CC, and thus HYC was more tolerant of mixed planting than CC and its photosynthetic capacity was superior.
The underlying mechanism of high yield formation of HYC
In general, the soybean seed yield is closely related to the leaf photosynthetic characteristics. Cui and Yu[50] determined a significant correlation between the seed yield and Lg in soybean during the late growth stages. Other studies also showed that the Ph values of the upper leaves are significantly correlated with the seed yield in soybean, particularly during the seed-filling stages [51-53]. Our results are consistent with those obtained in previous studies. We found that the seed yields from both HYC and CC were closely and positively correlated with Lg during MS and/or LS, and with Ph, St, and Tr during MS and/or LS (Fig. 5, Table 4), suggesting that MS and LS are crucial stages for the accumulation of photoassimilates in soybean. In other words, during these important growth periods, Lg, Ph, St, and Tr were higher in HYC than those in CC, indicating that the high yield formation in HYC was closely related to its superior leaf photosynthetic characteristics at late growing stages.
Outlook
In order to define the HYC photosynthetic advantage, this study preliminarily explored the differences of photosynthetic characteristics between HYC and CC through pot experiment under different cultivation practices. During the critical period for soybean seed yield formation, the photosynthetic characteristics of HYC were significantly higher than those of CC, and HYC was more resistance to dense and mixed planting than CC. Those superior performances of HYC in photosynthetic characteristics under different cultivation practices may be an important reason for its higher seed yield than CC. However, although the pot soil was taken from the field, the aggregate structure of the topsoil layer, soil permeability, aeration and fertilizer utilization rate have changed because of manual digging, mixing and no loss of nitrogen fertilizer. Therefore, the results from traits measured on pot-grown plants may be different from those in field. Thus, field comparative experiments should be conducted in future to verify the current results and deepen our understanding of the photosynthetic characteristics of HYC.
Author contributions
Conceived and designed the experiment: ZL and XB. Performed the experiments: ZL, XB and XY. Analyzed the data: XB and ZL. Contributed reagents/materials/analysis tools: XB and ZL. Wrote paper: ZL and XB.
References
[1] HARRISON SA, BOERMA HR, ASHLEY DA. Heritability of canopy-apparent photosynthesis and its relationship to seed yield in soybeans[J]. Crop Science, 1980(21): 222-226.
[2] LIU T, WANG Z, CAI T. Canopy apparent photosynthetic characteristics and yield of two spike-type wheat cultivars in response to row spacing under high plant density[J]. Plos One, 2016(11): e0148582.
[3] PENG S, KRIEG DR, GIRMA FS. Leaf photosynthetic rate is correlated with biomass and grain production in grain sorghum lines[J]. Photosynthesis Research, 1991(28): 1-7.
[4] ZHAO Y, ZHANG H, AO X, et al. Effect of different diammonium phosphate levels on nutrient uptake of super-High yielding soybean[J]. Scientia Agricultura Sinica, 2014(47): 2326-2334.
[5] ZHANG X, ZHAO C, BAI H, et al. Study on physiological indexes of summer soybean variety Youchu 4 yielding 4 500kg/ha[J]. Scienta Agricultura Sinica,1996(29): 46-54.
[6] WEI J, LUO G, ZHANG L, et al. Physiological parameters of super-high yielding soybean cultivar Zhonghuang 35[J]. Acta Agronomica Sinica, 2009(35): 506-511.
[7] ZHANG W, XIE F, ZHANG H, et al. Canopy and yield characteristics of super-high-yielding soybean cv. Liaodou No. 14[J]. Scientia Agricultura Sinica, 2007(40): 2460-2467.
[8] BUTTERY B. Effects of plant population and fertilizer on the growth and yield of soybeans[J]. Canadian Journal of Plant Science, 1969(49): 659-673.
[9] EGLI D. Plant density and soybean yield[J]. Crop Science, 1988(28): 977-981.
[10] SHARMA U, DATTA M, SHARMA V. Effect of applied phosphorus on the yield and nutrient uptake by soybean cultivars on acidic hill soil[J]. Open Journal of Soil Science, 2011(1): 45.
[11] WELLS R. Soybean growth response to plant density: Relationships among canopy photosynthesis, leaf area, and light interception[J]. Crop Science, 1991(31): 755-761.
[12] BOARD J. Light interception efficiency and light quality affect yield compensation of soybean at low plant populations[J]. Crop Science, 2000(40): 1285-1294.
[13] SINGER JW. Soybean light interception and yield response to row spacing and biomass removal[J].. Crop science, 2001(41): 424-429.
[14] BOARD J, KAMAL M, HARVILLE B. Temporal importance of greater light interception to increased yield in narrow-row soybean[J]. Agronomy Journal, 1992(84): 575-579.
[15] YUNUSA IAM, IKAWELLE MC. Yield response of soybean (Glycine max [L.] Merr.) to planting density and row spacing in a semi-arid tropical environment[J]. Journal of Agronomy and Crop Science, 2010(164): 282-288.
[16] ZHOU XJ, LIANG Y, CHEN H, et al. Effects of rhizobia inoculation and nitrogen fertilization on photosynthetic physiology of soybean[J]. Photosynthetica, 2006(44): 530-535.
[17] HUBER S, SUGIYAMA T, ALBERTE R. Photosynthetic determinants of growth in maize plants: effects of nitrogen nutrition on growth, carbon fixation and photochemical features[J]. Plant and Cell Physiology, 1989(30): 1063-1072.
[18] UPRETY D, MAHALAXMI V. Effect of elevated CO2 and nitrogen nutrition on photosynthesis, growth and carbon-nitrogen balance in Brassica juncea[J]. Journal of Agronomy and Crop Science, 2000(184): 271-276.
[19] BONDADA BR, SYVERTSEN JP. Leaf chlorophyll, net gas exchange and chloroplast ultrastructure in citrus leaves of different nitrogen status[J]. Tree Physiology, 2003(23): 553-559.
[20] HUANG ZA, JIANG DA, YANG Y, et al. Effects of nitrogen deficiency on gas exchange, chlorophyll fluorescence, and antioxidant enzymes in leaves of rice plants[J]. Photosynthetica, 2004(42): 357-364.
[21] MARCHESE J, KATZ I, SOUSA A, et al. Gas exchange in lisianthus plants (Eustoma grandiflorum) submitted to different doses of nitrogen[J]. Photosynthetica, 2005(43): 303-305.
[22] XIAO W, XIE F, ZHANG H, et al. Accumulation and utilization of nitrogen in super-high-yielding soybean cv. Liaodou 14[J]. Soybean Science, 2008(6): 960-965.
[23] MA Z, CHE R, WANG H, et al. Effect of different seeding rates and planting patterns on root morphological traits and root vigor of super-high-yield soybean cultivars[J]. Scientia Agricultura Sinica, 2015(48): 1084-1094.
[24] ZHANG X, ZHANG H, SONG S, et al. Comparison on root activity and nodulation characteristics of super-high-yielding soybeans[J]. Soybean Science, 2013(32): 496-500.
[25] ECHARTE L, MAGGIORA D, CERRUDO D, et al. Yield response to plant density of maize and sunflower intercropped with soybean[J]. Field Crops Research, 2011(121): 423-429.
[26] LIU T, SONG F. Maize photosynthesis and microclimate within the canopies at grain-filling stage in response to narrow-wide row planting patterns[J]. Photosynthetica, 2012(50): 215-222.
[27] LIU Y, ZHOU X, CHEN Y, et al. Effects of pre-sowing soil moisture and planting patterns on photosynthetic characteristics and yield of summer soybean[J]. Acta Ecologica Sinica, 2011(52): 216-227
[28] YANG F, LIAO D, WU X, et al. Effect of aboveground and belowground interactions on the intercrop yields in maize-soybean relay intercropping systems[J]. Field Crops Research, 2017(203): 16-23.
[29] FAN Y, CHEN J, WANG ZH, et al. Soybean (Glycine max L. Merr.) seedlings response to shading: leaf structure, photosynthesis and proteomic analysis[J]. BMC plant biology, 2019(19): 34.
[30] RAZA MA, LING Y, WOPKE W, et al. Narrow-wide-row planting pattern increases the radiation use efficiency and seed yield of intercrop species in relay‐intercropping system[J]. Food and Energy Security, 2019, 3.
[31] MA BL, MORRISON MJ, VOLDENG HD. Leaf Greenness and Photosynthetic Rates in Soybean[J]. Crop Science, 1995(35): 1411-1414.
[32] MONJE OA, BUGBEE B. Inherent limitations of nondestructive chlorophyll meters: a comparison of two types of meters Hort[J]. Science, 1992(27): 69-71.
[33] MATSUMOTO K, OHTA T, TANAKA T. Dependence of stomatal conductance on leaf chlorophyll concentration and meteorological variables[J]. Agricultural and forest meteorology 2005(132): 44-57.
[34] BUTTERY B, BUZZELL R, FINDLAY W. Relationships among photosynthetic rate, bean yield and other characters in field-grown cultivars of soybean[J]. Canadian Journal of Plant Science, 1981(61): 190-197.
[34] ZOU J, JIN X, ZHANG Y, et al. Effects of melatonin on photosynthesis and soybean seed growth during grain filling under drought stress[J]. Photosynthetica, 2019(57): 512-520.
[35] WOODWARD FI. Do plants really need stomata? [J]. Journal of Experimental Botany, 1998(49): 471-480.
[36] HETHERINGTON AM, F IAN W. The role of stomata in sensing and driving environmental change[J]. Nature, 2003(424): 901-908.
[37] COLLATZ GJ, BALL JT, GRIVET C, et al. Physiological and environmental regulation of stomatal conductance, photosynthesis and transpiration: a model that includes a laminar boundary layer[J]. Agricultural and Forest Meteorology, 1991(54): 107-136.
[38] YU GR, NAKAYAMA K, MATSUOKA N, et al. A combination model for estimating stomatal conductance of maize (Zea mays L.) leaves over a long term[J]. Agricultural and Forest Meteorology, 1998(92): 9-28.
[39] GAN Y, STULEN I, KEULEN HV, et al. Physiological response of soybean genotypes to plant density[J]. Field Crops Research, 2002(74): 231-241.
[40] ZHAO Y, XIAO X, BI D, et al. Effects of sulfur fertilization on soybean root and leaf traits, and soil microbial activity[J]. Journal of Plant Nutrition, 2008, 31:473-483. doi:10.1080/01904160801895001.
[41] TOLLENAAR M, DIBO A, AGUILARA A, et al. Effect of crop density on weed interference in maize[J]. Agronomy Journal, 1994(86): 591-595.
[42] RAMBO L, COSTA JA, PIRES JLF, et al. Soybean yield response to plant arrangement[J]. Ciência Rural, 2003(33): 405-411.
[43] JIN J, LIU X, WANG G, et al. Agronomic and physiological contributions to the yield improvement of soybean cultivars released from 1950 to 2006 in Northeast China[J]. Field Crops Research, 2010(115): 116-123.
[44] LIU G, YANG C, XU K, et al. Development of yield and some photosynthetic characteristics during 82 years of genetic improvement of soybean genotypes in northeast China Australian[J]. Journal of Crop Science, 2012(6): 1416.
[45] MORRISON MJ, VOLDENG HD, COBER ER. Physiological changes from 58 years of genetic improvement of short-season soybean cultivars in Canada[J]. Agronomy Journal, 1999(91): 685-689.
[46] WANG S, YANG Y, TRISHCHENKO AP, et al. Modeling the response of canopy stomatal conductance to humidity[J]. Journal of Hydrometeorology, 2009(10): 521-532.
[47] LEUNING R. A Critical appraisal of a combined stomatal-photosynthesis model for C3 plants [J]. Plant Cell & Environment, 2010(18): 339-355.
[48] SHARKEY TD, RASCHKE K. Effect of light quality on stomatal opening in leaves of Xanthium strumarium L[J]. Plant Physiology, 1981(68): 1170-1174
[49] YU Q, ZHANG Y, LIU Y, et al. Simulation of the stomatal conductance of winter wheat in response to light, temperature and CO2 changes[J]. Annals of Botany, 2004(93): 435-441.
[50] CUI S, YU D. QTL mapping of chlorophyll content at various growing stages and its relationship with yield in soybean (Glycine max L. Merr.)[J]. Acta Agriculturae Sinica, 2007(35): 744-750.
[51] SECOR J, MCCARTY DR, SHIBLES R, et al. Variability and selection for leaf photosynthesis in advanced generations of soybeans[J]. Crop Science, 1982(22): 255-259. doi:10.2135/cropsci1982.0011183X002200020013x.
[52] FORD DM, SHIBLES R, GREEN DE. Growth and yield of soybean lines selected for divergent leaf photosynthetic ability[J]. Crop Science, 1983(23): 517-520. doi:10.2135/cropsci1983.0011183X002300030018x.
[53] BOERMA HR, ASHLEY DA. Canopy photosynthesis and seed-fill duration in recently developed soybean cultivars and selected plant introductions[J]. Crop Science, 1988(28): 137-140. doi:10.2135/cropsci1988.0011183X002800010029x.
杂志排行

农业生物技术(英文版)的其它文章
- Anti-inflammatory Activity and Mechanism of Total Flavonoids from the Phloem of Paulownia elongate S.Y. Hu in LPS-stimulated RAW264.7 Macrophages
- Comparative Genomic Analysis of Boron Transport Gene Family in Arabidopsis and Five Crops
- Effects of Different Water-saving Irrigation Methods on Fruit Quality and Yield of Snow Melon
- Field Control Effects and Crop Safety Assessment of Triazole Fungicides on Apple Rust
- Effects of Acetylacetone Solution Soaking on Agrobacterium-transformed Maize Seed Buds
- Effects of Meteorological Factors on Overwintering Ability, Yield and Quality of Forage Rape