桃树花朵对低温胁迫的生理响应
2021-07-29刘兆宇张晓煜南学军胡宏远李芳红
刘兆宇,张晓煜,南学军,胡宏远,李芳红
桃树花朵对低温胁迫的生理响应
刘兆宇1,张晓煜2,3*,南学军2,3,胡宏远2,3,李芳红4
1. 曲阜师范大学计算机学院, 山东 日照 276800 2. 中国气象局旱区特色农业气象灾害监测预警与风险管理重点实验室, 宁夏 银川 750002 3. 宁夏气象科学研究所, 宁夏 银川 750002 4. 宁夏大学农学院, 宁夏 银川 750021
为揭示桃花受霜冻低温胁迫的生理响应过程,鉴定桃花抵御霜冻胁迫能力。选择5~6 a生“麦香”桃花,采用人工霜冻模拟试验箱模拟自然降温过程,对桃花进行-2.5 ℃、-3.0 ℃、-3.5 ℃、-4.0 ℃、-4.5 ℃以及-5.0 ℃的模拟低温处理,以室温(15 ℃)为对照。测定桃花抗寒生理指标和电导率,分析桃花受低温胁迫的生理响应过程。研究结果表明:“麦香”桃花半致死温度为-3.99 ℃;随着低温强度增强,桃树花朵中保护酶(SOD、POD、CAT)、渗透调节物质(可溶性蛋白质和可溶性糖)含量均呈现先上升后下降趋势,其峰值主要集中在-4.0~-3.5 ℃;脯氨酸呈现增加趋势且在-4.0 ℃时增长幅度最大;丙二醛含量增加呈慢—快—慢趋势,其含量在-4.0~3.5 ℃增长速度最快。SOD、POD、CAT、可溶性糖、可溶性蛋白、脯氨酸和MDA对低温响应过程表现出较好的一致性。通过对“麦香”桃花低温生理响应过程和一致性分析,结合各生理指标变化可确定5~6 a生“麦香”桃花受霜冻冻害的临界温度为-4.0~-3.5 ℃。
低温胁迫; 桃花; 生理响应
桃树L.)是蔷薇科(Rosaceae)李属()植物,在世界各地广泛种植。中国是桃的重要起源中心,经过4000多年的演化、栽培,形成以无锡水蜜、奉化玉露、深州蜜桃、肥城桃为代表的“四大名桃”。在1993年,中国桃树栽培面积和桃产量全面超过美国,成为桃产业第一大国[1]。截止2016年底,我国桃树种植面积约76.35万hm2,产量达到1383.31万t左右,国内市场总供给量达到1375.98万t[2],成为仅次于苹果、梨的第三大落叶果树。桃主要划分7个生态区,宁夏属于西北高旱生态区[3],光照充足、干燥少雨,气候条件十分适宜桃树生长[4],经过几十年发展,桃产业已成为宁夏自治区农民脱贫致富的重要支柱产业,在促进当地经济发展,增加农民收入方面发挥着重要作用。
近年来,随着全球气候变暖,果树越冬冻害风险有所降低,但春季温度回升过早使果树提前萌发,受霜冻灾害危害的风险增加[5,6]。频繁发生的晚霜冻对桃以及其他林果产业带来难以估量的损失[7-8]。宁夏平原处于东亚季风气候的边缘地带,春季冷暖交替频繁、气温极不稳定[9],早熟桃品种始花期在每年4月10日左右,但宁夏晚霜终止日在5月下旬[10]。桃开花期、幼果期与霜冻高频期高度重叠。春霜冻已成为制约宁夏桃产业品质提升、稳产高产的主要限制性因素之一。
关于桃花霜冻研究多集中在霜冻防御、桃花期风险区划[11-13]以及通过桃花表型反映受冻程度[14]等方面。本试验从低温下桃花生理响应温度与生理指标一致性方面开展。以5~6 a生“麦香”桃(mai xiang)花朵为试材,通过模拟自然霜冻连续降温过程,研究持续降温过程中桃花半致死温度、保护酶系统、渗透调节物质及丙二醛等[15-19]生理指标变化,分析桃花在低温下不同生理指标敏感性和一致性,从生理角度鉴定桃花抗低温能力,明确桃花在低温下各项生理指标的响应温度,为提高桃产业预防霜冻和低温冻害监测预警水平提供科学理论依据。
1 材料与方法
1.1 试验地概况
试验于2019年4月在宁夏银川市永宁县望洪镇进行。试验地段海拔1100 m,年均气温9.6 ℃,≥10 ℃年积温3427.1 ℃·d,年降水量177.2 mm,年均日照时数2893 h,年均相对湿度65%,无霜期167 d。试验土壤类型为灌淤土,占地1 hm2,株行距3 m×4 m,树体整形方式为Y型,常规管理。
1.2 试验材料
试验材料为树势、营养状况一致5~6 a生早熟桃 “麦香”大蕾期花枝。于树冠中部东、西、南、北四个方位采桃树大蕾期花枝,每个方向采4枝带花枝条,每棵树取15枝带花枝条混合样,将其插入水培缸中备用。
1.3 试验材料的低温处理
桃花低温处理与生理检测在中国气象局旱区特色农业气象灾害监测预警与风险管理重点实验室理化分析室进行。霜冻试验仪器采用中国农业科学院开发的以PID调节方式控温的MSX-2F型模拟霜箱系统。
将带花朵的花枝连同水培缸置于霜冻模拟试验箱,按照图1所示设置降温曲线,温度梯度设定为-2.5 ℃,-3.0 ℃,-3.5 ℃,-4.0 ℃,-4.5 ℃和-5.0 ℃,以室温(15 ℃)为对照。在连续降温过程中在达到每个处理温度并持续0.5 h之后取出一批样品。对不同低温处理和对照的样本进行随机抽取,进行生理指标测定。
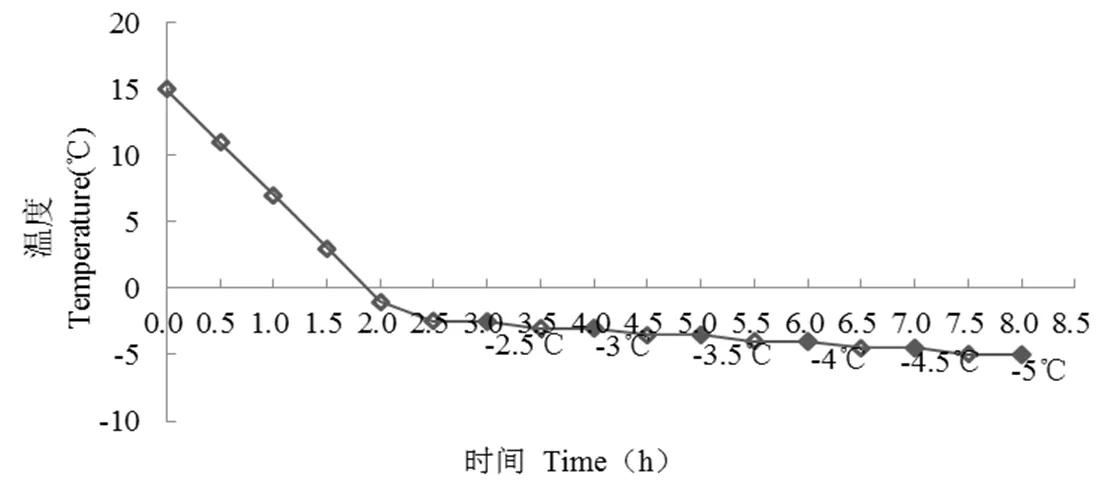
图1 桃树花朵人工模拟降温曲线
1.4 测定指标与方法
电导率采用电导法测定[20];
SOD测定采用南京建成试剂盒方法(货号:A001-1);
POD测定采用南京建成试剂盒方法(货号:A084-3-1);
CAT测定采用南京建成试剂盒方法(货号:A007-1);
MDA测定采用硫代巴比妥酸法[20];
游离脯氨酸测定采用磺基水杨酸浸提-酸性茚三酮显色法[20];
可溶性蛋白测定采用考马斯亮蓝染色法[20];
可溶性糖测定采用蒽酮比色法[20]。
1.5 数据处理与分析
半致死率采用电导法配以Logistic方程进行拟合,根据(1)计算“麦香”桃花样品的半致死温度。
=/(1+-bx) (1)
式中:为该温度下相对电导率,为处理温度,为电导率的饱和容量,、为系数。
采用SPSS 19.0对数据进行处理分析。
2 结果分析
2.1 不同低温处理后桃花的电导率
电解质渗透率反应生物膜透性破坏的程度。经过低温的处理,桃花相对电导率表现不同,随低温强度增强而增加。从室温下降至-3.5 ℃前,桃花电解质渗透率稳步上升,但不具有显著性变化(>0.05),温度下降至-3.5 ℃时,“麦香”桃花的电解质渗透率基本维持在30%左右;当低温强度到达-4.0 ℃时,桃花电导率显著升高(<0.05)至52%(图2),说明当温度下降至-4 ℃时,细胞膜严重受损电解质大量外渗。后随低温强度增强,桃花电导率增长速率有所下降。
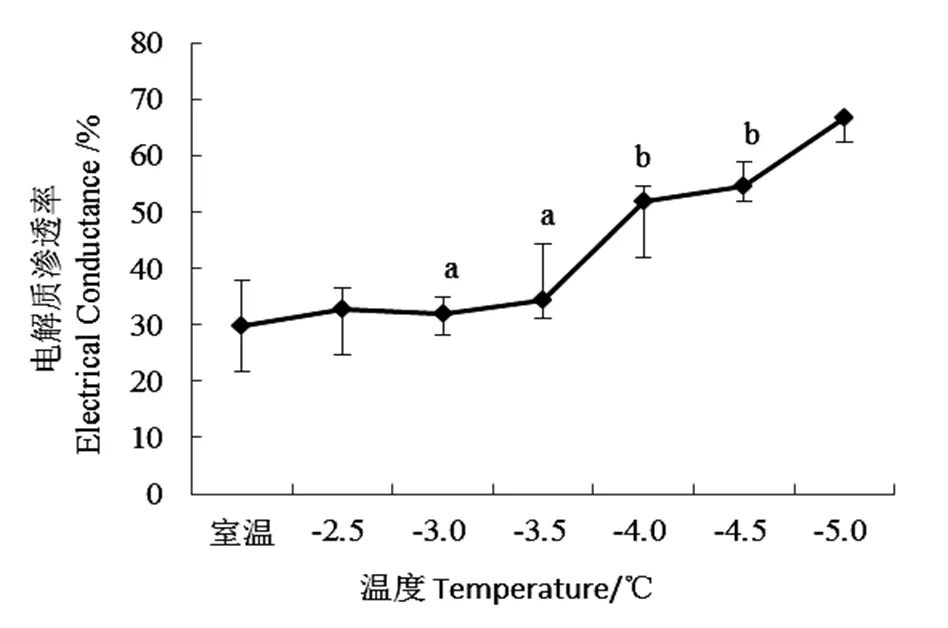
图2 不同低温胁迫下“麦香”桃花电解质渗透率
2.2 桃花半致死温度
半致死温度是指在某一温度条件下,植物达到半致死状态,如温度继续低于这一温度时,植物将遭受不可逆伤害乃至死亡。在-3.0 ℃、-4.0 ℃、-5.0 ℃低温作用下,运用Logistic方程拟合相对电导率与低温的关系,求解“麦香”桃花半致死温度为-3.99 ℃(表1),此时桃花处于半致死状态。其决定系数2达到0.9922,拟合度较好。

表1 不同低温胁迫下“麦香”桃花电导率与半致死温度
2.3 桃花抗逆酶对不同低温处理的响应
SOD酶催化产生歧化反应,是重要的逆境指标。由图3可得,随低温强度增强桃花保护酶活性均整体呈现先升后降趋势。从室温下降至-3.5 ℃前,SOD活性持续上升,当温度下降至-3.5 ℃时,SOD活性达到最高值,后随温度强度SOD活性开始呈现下降趋势。在-4~-3.5 ℃时SOD活性显著下降(<0.05),可知-3.5 ℃是低温胁迫对桃花SOD的抑制温度。POD是防止膜脂过氧化的主要酶,从室温下降至-3.5 ℃前,桃花POD活性呈上升趋势。温度下降至-3.5 ℃时POD活性达到峰值且与-3.0 ℃、-4.0 ℃处理下POD活性存在显著性差异(<0.05),后随低温强度增强POD活性开始呈现下降趋势。从室温下降至-3.5 ℃前,桃花CAT活性整体呈上升趋势,当温度降至-3.5 ℃时达到峰值。随低温强度增强,温度在-4.0~-3.5 ℃CAT活性显著下降(<0.05)。温度低于-4.0 ℃后CAT活性不具有显著性变化,可知桃花CAT活性低温敏感温度为-3.5 ℃左右。
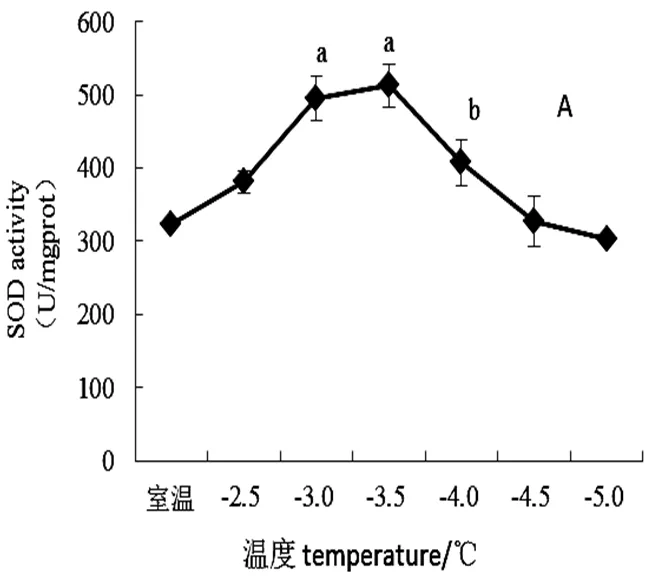
图3 不同低温胁迫下“麦香”桃花抗逆酶含量变化
2.4 桃花主要渗透调节物质对不同低温的响应
由图4A可知,随低温强度增强,桃花可溶性蛋白含量整体呈先升后降趋势。室温时桃花可溶性蛋白质含量为2.31±0.14 mg/gFW。从室温下降至-4.0 ℃前,可溶性蛋白含量呈上升趋势。当温度下降至4.0 ℃时,可溶性蛋白含量达到最高(3.03±0.01 mg/gFW)较对照增加0.72 mg/gFW,具有显著变化。室温至-4.0 ℃阶段这可能是桃花对轻度低温适应的结果。温度低于-4.0 ℃后可溶性蛋白含量开始呈现下降趋势。
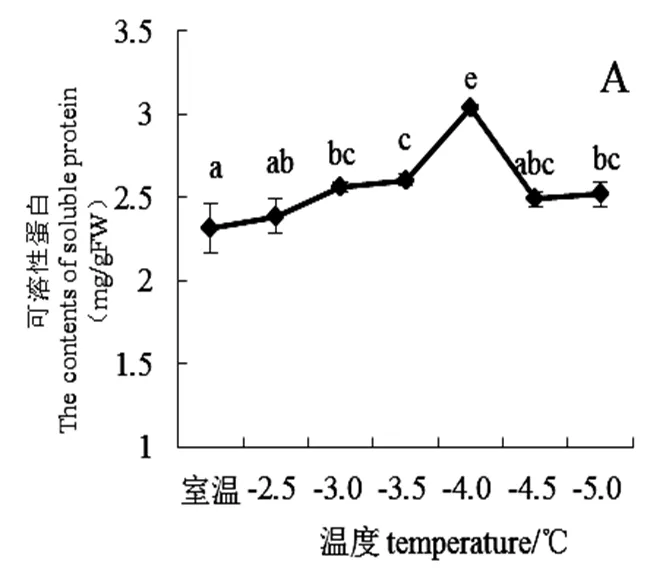
图4 不同低温胁迫下“麦香”桃花渗透调节物质含量
由图4B可知,桃花可溶性糖含量随低温强度增强呈先升高后下降趋势。从室温下降至-3.5 ℃前,桃花可溶性糖含量变化不具有显著性差异。温度下降至-4.0 ℃时可溶性糖含量达到峰值(29.13±1.25 mg/gFW),较对照增加4.76 mg/gFW具有显著性变化。随低温强度增强,可溶性糖含量出现显著性下降(<0.05),说明此时桃花已不能通过增加可溶性糖含量来抵御低温胁迫。
由图4C可知,脯氨酸含量随温度下降整体呈上升趋势,温度下降至-3.5 ℃时较室温脯氨酸含量显著(<0.05)上升。低温强度在-3.5~-4.0 ℃阶段桃花体内脯氨酸含量呈小幅度下降,低温低于-4 ℃后脯氨酸含量升高但无显著性变化,这可能是低温强度到达-4.0 ℃时,超出了桃花对低温的最低耐受能力。
2.5 桃花MDA对低温的响应
MDA是膜脂过氧化作用分解的最终产物。由图5可知,随着低温强度增强MDA含量整体呈上升趋势。从室温下降至-3.5 ℃前,丙二醛含量无显著性变化。当温度降至-4.0 ℃时MDA含量急剧升高,较对照增加1.15 mmol/gFW变化显著。低温低于-4 ℃后,MDA含量无显著性变化。
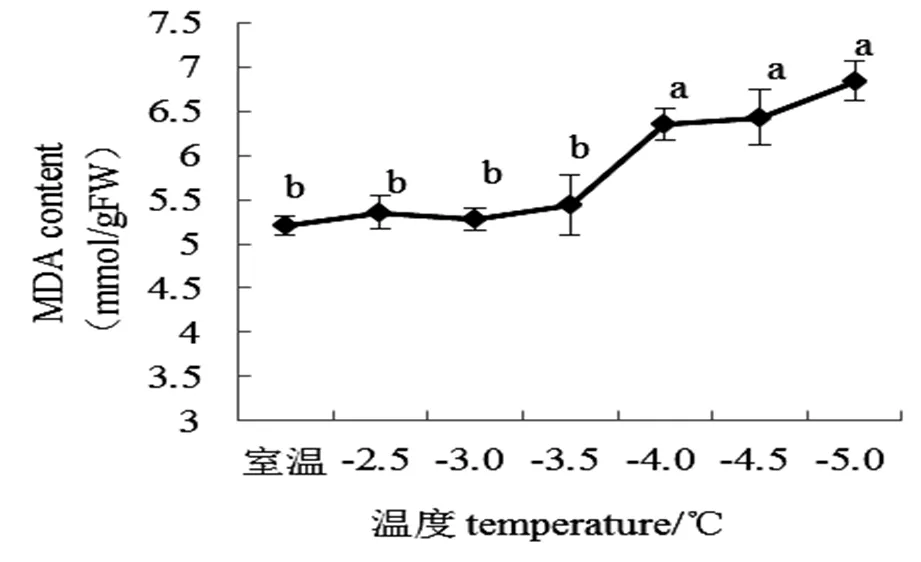
图5 不同低温胁迫下“麦香”桃花MDA含量
3 讨论
细胞膜是细胞内外物质交换的重要媒介,是细胞感受外界环境最敏感部位。电解质渗透率增加是低温胁迫对生物膜透性破坏的效应[21]。降温前期电导率上升缓慢是桃花对于低温胁迫适应阶段,后电解质渗出率迅速升高超过50%最后趋于缓慢,说明细胞膜此时已经被破坏,大量电解质外渗。运用Logistic方程拟合相对电导率与低温的关系,求得桃花半致死温度是-3.99 ℃。
SOD、POD、CAT是植物中重要的抗逆酶[22]。正常条件下,植物体内活性氧积累与清除处于动态平衡,遭受逆境时活性氧积累。SOD是清理活性氧的重要酶类,POD通过转化植物体内碳水化合物成为木质素来增强抵御逆境的能力[23],CAT是一种关键的生物防御酶。轻度低温胁迫可诱导并激发保护酶活性,增强植物抗逆性。温度下降至-3.5 ℃时桃花SOD、POD、CAT活性达到最高然后出现显著性下降,表明桃花SOD、POD和CAT低温敏感温度为-3.5 ℃,此时桃花体内自由基平衡受到破坏,活性氧积累。后随低温强度增强桃花不能通过提高保护酶活性途径抵御低温逆境。本试验中SOD、POD和CAT对低温的响应温度相对于其他指标较为敏感。
可溶性蛋白、可溶性糖、游离脯氨酸是植物体内广泛分布的主要渗透调节物质。一般认为可溶性蛋白质、可溶性糖的含量与植物的抗寒性相关,一定逆境强度下两者含量升高,但超过自身极限逆境其合成水平会逐步下降[24-26]。脯氨酸是植物在逆境下的产物,随温逆境增强,游离脯氨酸迅速积累,是植物对低温的适应,但也有研究认为这两者没有必然联系[27]。
渗透调节物质有助于提高细胞液浓度,维持细胞膨压,减缓原生质脱水。本研究表明,随低温强度增强,桃花体内可溶性糖、可溶性蛋白呈现先上升后下降趋势。这表明,在一定轻度低温胁迫下桃花体内积累大量渗透调节物质抵御胁迫。温度下降至-3.5 ℃时可溶性糖含量达到最高,表明温度下降至-3.5 ℃前桃花可以通过提高可溶性糖含量,通过增加保水能力、降低水势等途径避免低温下原生质因结冰而受伤害致死。温度下降至-4.0 ℃时桃花已不能通过提高可溶性糖含量这一途径抵御低温逆境,但仍可通过提高可溶性蛋白含量抵御低温逆境。当低温下降至-4.0 ℃后,可溶性蛋白合成受阻含量显著下降,证明此时不能通过提高可溶性蛋白含量这一途径抵御低温逆境,同时可溶性蛋白分解产生脯氨酸,导致逆境后期脯氨酸含量有小幅度升高。在温度下降至-4.0 ℃时脯氨酸含量出现小幅下降,证明在-4 ℃时桃花已经不能通过自身合成脯氨酸一提高含量,但由于脯氨酸属于氨基酸,可由可溶性蛋白在低温逆境下分解产生,导致脯氨酸含量有小幅度增加。其中可溶性蛋白变化规律与杨春祥[28]基本一致。由此可得,桃花渗透调节物质对低温敏感温度为-3.5~-4.0 ℃左右。
研究表明,MDA作为膜脂过氧化最终产物之一,属于植物膜系统对胁迫环境响应的重要指标[29]。本实验中,降温前期MDA含量上升缓慢但不具有显著性是因为桃花通过系列途径抵御逆境免受伤害的结果。温度下降至-4.0 ℃时MDA含量显著(<0.05)上升。低温低于-4.0 ℃后,MDA含量随逆境增强增加但不具有显著性差异,表明温度在下降至-4.0 ℃时活性氧已完全破坏了膜脂,使细胞功能完全受损。
实验中,保护酶、可溶性糖和脯氨酸表现出对低温响应较好的一致性,且对低温敏感性较强,在低温逆境初期起主要抗逆作用。可溶性蛋白、MDA表现了较好一致性,对低温的敏感性较弱。可能是因为不同植物体内各物质含量以及物质形成机制不同,从而抵御逆境的途径有所差别,不同生理物质在不同阶段启动响应机制。低温逆境前期,低温敏感性较强的指标起主要抗逆作用,温度低于-3.5 ℃后对低温敏感性较强的指标抗逆性减弱,到达半致死温度-3.99 ℃时,敏感性较弱的指标达到峰值。不同物质在低温胁迫下响应强度和时间上有一定差别,反映桃花低温抗逆的生理过程。限于试验条件,本文没有进行多品种试验异,也未区分桃树营养和健康状况。因此,如果要全面阐述桃树花期对低温胁迫的生理响应,需要进一步研究桃树品种、花期、组织以及低温持续时间等方面。
4 结论
本试验结果表明,桃树花生理指标对低温响应有较好的一致性。与对照相比,桃花保护酶、脯氨酸和可溶性糖等生理指标均在温度下降至-3.5 ℃时有显著变化,这些生理过程响应低温胁迫的敏感性较强,是桃花适应逆境初期信号。可溶性蛋白和MDA响应温度为-4.0 ℃,这两个生理指标对低温的响应较迟缓,是在低温强度达到一定量值后才有所响应。实验中桃花半致死温度为-3.99 ℃。因此,初步确定5~6年生“麦香”桃花受冻临界温度为-4.0~-3.5 ℃。
[1] 俞明亮,王力荣,王志强,等.新中国果树科学研究70年——桃[J].果树学报,2019,36(10):1283-1291
[2] 邓秀新,束怀瑞,郝玉金,等.果树学科百年发展回顾[J].农学学报,2018,8(1):24-34
[3] 汪祖华,庄恩及.中国果树志桃卷[M].北京;中国林业出版社,2001
[4] 陈艳玲,喻菊芳,吕国华,等.宁夏桃产业发展中存在的问题及对策[J].北方园艺,2012(13):196-198
[5] 肖芳,段晓凤,李红英,等.宁夏枸杞春季霜冻指标研究[J].经济林研究,2019(3):193-197
[6] Bennie J, Kubin E, Wiltshire A,. Predicting spatial and temporal patterns of budburstand spring frost risk in north-west Europe: the implicationsof local adaptation to climate [J]. Global Change Biol, 2010,16:1503-1514
[7] Augspurger CK. Reconstructing patterns of temperature ,phenology and frost damage over 124 years: spring damage risk is increasing [J]. Ecology, 2013,94:41-50
[8] Crimp S, Zheng B, Khimashia N,Recent changes in Southern Australian frost occurrence: implications for wheat production risk [J]. Crop and Pasture Science, 2016,67(8):801-811
[9] 杨洋,张磊,陈豫英,等.宁夏贺兰山东麓酿酒葡萄种植区春霜冻发生时间规律研究[J].自然灾害学报,2017,26(4):84-90
[10] 张磊,王静,张晓煜,等.近50 a宁夏初、终霜日基本特征及变化趋势[J].干旱区研究,2014,31(6):1039-1045
[11] 王俊杰.复合植物生长调节剂延迟李树和桃树花期试验[J].北方园艺,2012(6):20-22
[12] 姚晓红,万信,贾建英,等.天水蜜桃花果期霜冻气候风险区划研究[J].西南师范大学学报(自然科学版),2016,41(7):144-150
[13] 许彦平,姚晓红,万信,等.天水蜜桃开花坐果期霜冻灾害气象风险评估[J].中国农业气象,2013,34(5):606-610
[14] 许彦平,万信,贾建英,等.北京7号桃树()盛花期霜冻害指标试验研究[J].干旱区资源与环境,
2019,33(11):195-200
[15] Caradonna PJ, Bain JA. Frost sensitivity of leaves and flowers of subalpine plants is related to tissue type and phenology [J]. Journal of Ecology, 2016,104(1):55-64
[16] 刘玉祥,马艳芝,张胜珍,等.桃花芽不同受冻情况其抗寒性生理指标的变化规律[J].贵州农业科学,2009,37(9):200-202
[17] 赵海亮,赵文东,高东升.冰冻低温处理下桃芽水分存在状态和膜透性与休眠解除的关系[J].北方园艺,2008(4):1-4
[18] 郜怀峰,张亚飞,王国栋,等.钼在桃树干旱胁迫响应中的作用解析[J].植物学报,2019,54(2):227-236
[19] 曹建东,陈佰鸿,王利军,等.葡萄抗寒性生理指标筛选及其评价[J].西北植物学报,2010,20(11):2232-2239
[20] 赵世杰,苍晶.植物生理学实验指导[M].北京:中国农业出版社,2015
[21] 简令成.植物抗寒机理研究的新进展[J].植物学通报,1992(3):17-22,16
[22] Mkersite BD. Manipulating freezing tolerance in transgenic plant [J].Acta Physiol Planta, 1997,19(4):17-19
[23] 李惠,熊忠飞,徐梦君,等.龙薯九号甘薯冷害胁迫与适温贮藏品质调控[J].食品科技,2018,43(12):37-42
[24] 王丽雪,李荣富.葡萄枝条中淀粉,还原糖及脂类物质变化与抗寒性的关系[J].内蒙古农业大学学报(自然科学版),1994(4):1-7
[25] 姚远,闵义,胡新文,等.低温胁迫对木薯幼苗叶片转化酶及可溶性糖含量的影响[J].热带作物学报,2010,31(4):556-560
[26] Couée I, Sulmon C, Gouesbet G,Involvement of soluble sugars in reactive oxygen species balance and responses to oxidative stress in plants [J].Journal of Experimental Botany, 2006,57(3):449-459
[27] 汤章城.逆境条件下植物脯氨酸的累积及其可能的意义[J].植物生理学通讯,1984(1):15-21
[28] 杨春祥.早熟油桃抗寒性的研究[D].泰安:山东农业大学,2005
[29] 何琴飞,曹艳云,彭玉华,等.不同相思树种单株抗寒性综合评价[J].生态学杂志,2019,38(5):1339-1345
Physiological Response of Peach Blossom to Low Temperature Stress
LIU Zhao-yu1, ZHANG Xiao-yu2,3*, NAN Xue-jun2,3, HU Hong-yuan2,3, LI Fang-hong4
1.276800,2.750002,3.750002,4.750021,
To reveal the physiological response of peach flowers to low temperature and frost stress.Identification of peach blossoms' ability to resist frost stress.Select 5~6a raw “maixiang” peach flowers, and use artificial frost simulation test box to simulate natural cooling process.Peach blossoms were subjected to simulated low temperature treatments of -2.5, -3.0 , -3.5, -4.0, -4.5, and -5.0 ℃ with room temperature (15 ℃) as a control.The physiological indexes and electrical conductivity of cold resistance of peach blossoms were measured, and the physiological response of peach blossoms to low temperature stress was analyzed.The results show that the "lethal" temperature of peach blossom is -3.99 ℃; As the low temperature strength increases,with the increase of low-temperature intensity, the contents of protective enzymes and osmoregulation substances in peach flowers increased first and then decreased.The peak is mainly concentrated at -4.0~-3.5 ℃. Proline showed an increasing trend with the largest increase at -4.0 ℃; malondialdehyde content showed a slow-fast-slow trend, and its content increased fastest at -4.0~3.5 ℃.SOD, POD, CAT, soluble sugar, soluble protein, proline and MDA showed good consistency to the low temperature response process.Through the low temperature physiological response process and consistency analysis of "mai xiang" peach flowers,Combining the changes of various physiological indicators, the critical temperature of 5~6 a raw “mai xiang” peach flower to be frost-frozen was determined to be -4.0~-3.5 ℃.
Low temperature stress; peach blossom; physiological response
S661.101
A
1000-2324(2021)03-0402-06
2020-10-12
2020-12-08
国家自然科学基金(41675114)
刘兆宇(1995-),男,助教,研究方向为果树生理生态. E-mail:zhyliu@foxmail.com
Author for correspondence. E-mail:zhang_xynet@163.com
