CO2加富与外源ABA对受旱黄瓜幼苗生长及叶片内源激素含量的影响
2021-07-29贺新蕊李清明
贺新蕊,李清明,
CO2加富与外源ABA对受旱黄瓜幼苗生长及叶片内源激素含量的影响
贺新蕊1,李清明1,2*
1. 山东农业大学园艺科学与工程学院, 山东 泰安 271018 2. 作物生物学国家重点实验室, 山东 泰安 271018
以‘津优35号’水培黄瓜幼苗为试材,在营养液中添加PEG-6000模拟干旱胁迫,研究了CO2加富与外源ABA对干旱胁迫下黄瓜幼苗生长及叶片内源激素含量的影响。结果表明,CO2加富可以显著促进干旱胁迫下黄瓜幼苗的生长,增加植株的株高、茎粗、叶面积以及干鲜重,提高生长素(IAA)、细胞分裂素(CTK)、赤霉素(GA)和乙烯(ETH)的含量,降低脱落酸(ABA)的含量,提高(IAA+GA+CTK)/ABA的比值;外源喷施ABA显著提高了黄瓜幼苗的茎粗及干鲜重,增加了ETH和ABA的含量而降低了IAA、CTK、GA含量和(IAA+GA+CTK)/ABA的比值。综上所述,CO2加富与外源ABA可通过对内源激素的调控来维持黄瓜幼苗在干旱胁迫条件下的生长。
黄瓜; 干旱胁迫; CO2加富; 外源激素
黄瓜(L.)是一种深受人们喜爱的蔬菜作物,但由于其浅根系的生物学特性,吸水能力差,极易受到水分亏缺的影响。干旱胁迫是自然界中制约植物生长发育最严重的非生物胁迫因子[1]。因此,开展黄瓜抗旱性研究意义重大。
自工业革命以来,全球平均气温上升了0.85 ℃,大气CO2浓度从280 μmol·mol-1上升到410 μmol·mol-1以上[2],预计到本世纪末会达到倍增[3]。大气CO2浓度的升高不仅会影响全球的气候变化,还会影响植物的生长发育。已有大量研究表明,CO2加富在植物的形态结构[4,5]、根系发育[6,7]、光合特性[8,9]、碳氮代谢[10]、气孔运动[11,12]等方面起着重要作用,还能缓解干旱[13]、高温[14]、盐害[15]等非生物胁迫对植物造成的伤害。脱落酸(ABA)是植物体内的五大激素之一,在植物应对各种胁迫因子的响应中起关键作用,又被称为应激激素[16],它可以响应非生物胁迫的信号,并在植物体内引发一系列的生理反应来应对胁迫[17]。当植物遭受到干旱胁迫时,NCED3的基因表达会被诱导,ABA合成加快,促使气孔关闭,降低蒸腾速率,减少水分的散失,提高植物水分利用效率[18]。Wang等[19]研究结果表明,外源施加ABA可以提高猕猴桃叶片中抗氧化酶的活性,增强其抗旱性,与此同时,ABA还可以提高植物体内渗透调节物质的含量,增强植物在干旱胁迫下的渗透调节能力[20]。
目前已有大量的研究表明CO2加富和外源ABA可以缓解植物的干旱胁迫,但多是针对的单一的CO2加富或外源喷施ABA。其实,CO2与ABA信号在对抗植物干旱胁迫的作用中有多种交叉,可共同作用来使植物适应不良环境[21,22]。因此,本试验选用‘津优35号’水培黄瓜幼苗为试材,并在营养液中添加PEG-6000以模拟干旱胁迫,研究了CO2加富和外源施加ABA对干旱胁迫下黄瓜幼苗生长以及内源激素水平的影响,以期从激素水平探明CO2加富和ABA缓解黄瓜幼苗的耐旱性机制。
1 材料与方法
1.1 材料
本试验于2020年9月-2020年11月在山东农业大学科技创新园进行,供试黄瓜品种为‘津优35号’,购自天津市科润黄瓜研究所,供试外源ABA购买自Solarbio科技有限公司。黄瓜种子点播在50孔的穴盘中,一穴一粒,基质配制比例为草炭:蛭石:珍珠岩=3:1:1。当幼苗生长到一叶一心时进行水培,水培幼苗要长势一致,水培容器为黑色的塑料盆(内径为长×宽×高= 37.5 cm×29 cm×12 cm),每盆定植6株。营养液采用全浓度的日本山崎黄瓜专用配方,微量元素为全剂量,PH控制在6.8~7.0之间,试验期间,营养液用气泵进行换气,每3 min通气30 s,并每天都将营养液补齐至刻度线,每3 d更换一次营养液。
1.2 方法
1.2.1 试验设计待幼苗长至两叶一心时,选取长势一致的植株。采用裂区试验设计,主区因素为CO2处理,设大气CO2浓度(≈400 μmol·mol−1)和加富CO2浓度(800±40 μmol·mol−1)2个水平;裂区因素为外源喷施ABA及其抑制剂,共4个水平:喷施去离子水(对照)、喷施20 μmol·L-1ABA、喷施2 mmol·L-1ABA抑制剂钨酸钠(Na2WO4)、喷施2 mmol·L-1ABA抑制剂钨酸钠+20 μmol·L-1ABA。每天喷施2次,分别是8:00和18:00,连续喷施2 d,随后进行干旱处理,即在营养液中添加5%的PEG-6000模拟干旱胁迫,渗透势(ψ)为−0.05 MPa。每个处理重复4次,每次重复3盆。干旱胁迫5 d开始取样测定各项指标。CO2气体由液态CO2钢瓶补充,拱棚内装有CO2浓度监测系统(北京奥托,2000),当棚内CO2浓度不在目标范围内时,系统会开启电磁阀自动补气。
1.2.2 测定项目与方法(1)生长指标测定株高用直尺测定;茎粗用游标卡尺测定;叶面积是用公式S=L计算,代表真叶的叶长[23];干鲜重采用称重法测定,先分别称取植株地上部和地下部鲜重,然后放在烘箱中烘干至恒重,称取干重。
(2)内源激素含量测定采用酶联免疫分析方法(ELISA)对叶片中生长素(IAA)、细胞分裂素(CTK)、赤霉素(GA)、脱落酸(ABA)及乙烯(ETH)含量进行测定,ELISA试剂盒由江苏酶免实业有限公司提供。
(3)数据分析采用软件Microsoft Excel 2019和DPS15.10来进行数据处理,采用Duncan检验法进行多重比较和差异显著性检验(=0.05),使用SigmaPlot12.5软件进行作图。
2 结果与分析
2.1 CO2加富与外源ABA对受旱黄瓜幼苗生长特性的影响

表1 CO2加富与外源ABA对受旱黄瓜幼苗生长的影响
备注:同列不同小写字母表示处理间差异显著(<0.05),下同。
Note: Different lower-case letters in the same column indicate a significant difference among treatments (Duncan’s multiple range test,< 0.05). The same below.
由表1可以看出,干旱胁迫下,CO2加富可以显著促进黄瓜幼苗的生长,具体表现为CO2加富可以在不同程度上增加植株的株高、茎粗、叶面积以及干鲜重,且均有显著性差异。ABA处理后植株茎粗及干鲜重明显高于H2O处理,植株的茎粗、鲜重、干重较喷施去离子水分别增加了7.05%、14.24%、22.41%,且差异均达显著水平,但对植株的株高及叶面积没有明显的促进作用。Na2WO4处理下植株的生长量均最低,Na2WO4+ABA处理的各项指标均显著高于Na2WO4处理,这也说明外源ABA可以有效缓解干旱胁迫对植株生长的抑制,而在CO2加富下喷施ABA更有利于干旱胁迫下植株的生长。
2.2 CO2加富与ABA对受旱黄瓜幼苗叶片内源激素含量的影响
由图1可知,CO2加富可以提高生长素(IAA)、细胞分裂素(CTK)、赤霉素(GA)和乙烯(ETH)的含量,降低脱落酸(ABA)的含量;同一CO2浓度下,与H2O处理相比,喷施ABA后,生长素(IAA)、细胞分裂素(CTK)赤霉素(GA)含量显著下降,但乙烯(ETH)和脱落酸(ABA)含量增加;喷施钨酸钠之后,脱落酸(ABA)、乙烯(ETH)、生长素(IAA)、细胞分裂素(CTK)含量均最低;Na2WO4+ABA处理的各激素含量较Na2WO4处理均有不同程度的提高。
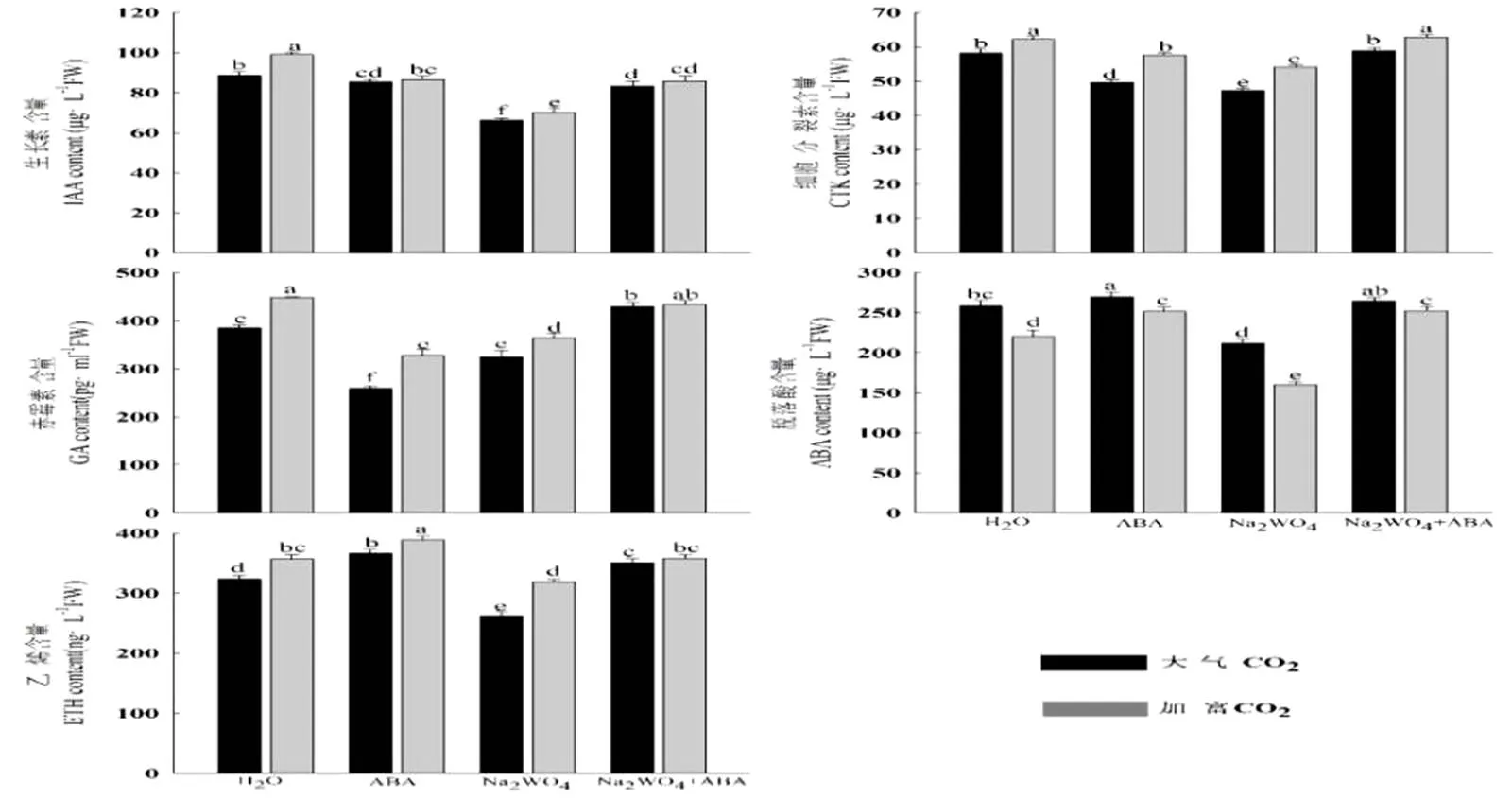
图1 CO2加富与外源ABA对受旱黄瓜幼苗内源激素含量的影响
2.3 CO2加富与ABA对受旱黄瓜幼苗叶片(IAA+GA+CTK)/ABA的影响
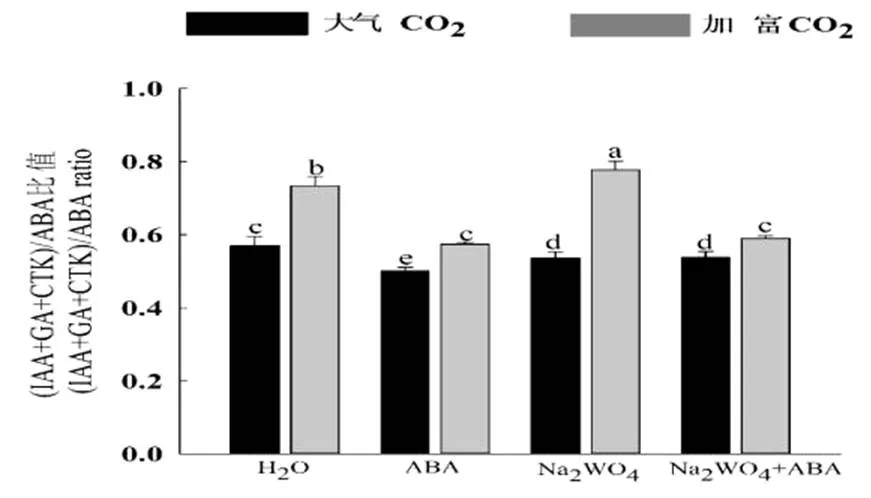
图2 CO2加富与外源ABA对受旱黄瓜幼苗(IAA+GA+CTK)/ABA比值的影响
(IAA+GA+ZR)/ABA的比值变化可以反映生长促进类激素和生长抑制类激素在植物生长发育过程中的平衡。由图2可以看出,CO2加富可以显著提高各处理下(IAA+GA+CTK)/ABA的比值,与大气CO2浓度相比,分别提高了28.55%、14.57%、44.88%、9.46%;外源喷施ABA后,由于生长类激素IAA、CTK、GA含量的下降,ABA含量的增加,所以导致(IAA+GA+CTK)/ABA的比值显著降低,与H2O处理相比,在大气CO2浓度和加富CO2浓度下分别下降了12.26%、21.8%;Na2WO4处理下(IAA+GA+CTK)/ABA的比值显著提高,但这主要是因为Na2WO4可以抑制ABA的合成,造成ABA含量的显著下降,所以导致比值的上升,并不意味着该处理下植物的生长受促进。
3 讨 论
干旱胁迫对植物的生长有显著抑制作用,最直观的表现就是生物量的下降。但有大量报道称,CO2加富可以促进干旱胁迫下植物生长,增加植株生物量的积累,尤其是对C3植物而言,在加富CO2浓度下,地上部分平均能增加20%的生物量[24,25]。这与本试验的研究结果一致,本研究发现CO2加富显著促进了干旱胁迫下黄瓜幼苗的生长,提高了株高、茎粗、叶面积以及干鲜重(表1),这主要是因为CO2加富增加了植物的碳供应,加速了植物激素促进的库器官生长,从而改变了茎的初生分生组织和次生分生组织活性[26]。同时,CO2加富可以促进叶片细胞分裂和细胞膨大,从而导致叶面积增加[27],这些变化增加了植物在高CO2浓度下的相对生长速率[28],并最终导致生物量的增加。ABA可作为一种应急胁迫激素在干旱胁迫中起信号转导的作用,从而减缓植物在干旱胁迫下受到的损伤,维持植物在干旱条件下的相对正常的生长状态[17],因此,外源施加ABA可以增加干旱胁迫下黄瓜幼苗的茎粗及干鲜重,这与李婉婷等[29]在滇润楠幼苗上的研究结果相似。且在本试验中,CO2加富与ABA互作时,黄瓜幼苗的生长状态最佳。
植物激素是一类非常微量的有机物,在调节植物生长发育和信号转导方面意义重大,参与植物对各种生物和非生物胁迫的响应[30]。植物激素的含量与外界环境的变化息息相关,依据其作用,主要被分为两类,一类是生长激素,主要包括生长素(IAA)、赤霉素(GA)、细胞分裂素(CTK)等,一类是应激激素,主要包括脱落酸(ABA)和乙烯(ETH)等[31]。干旱胁迫下植物的生长会受到抑制,生长类激素的减少就是主要原因之一[32]。目前,已有报道证明CO2加富会影响植物体内内源激素的含量,但其作用机制尚不清楚。Li等[33]发现,CO2加富可以显著增加油松在胁迫下生长素(IAA)、异戊烯基腺嘌呤核苷(iPA)、二氢玉米素核苷(DHZR)的含量,降低脱落酸(ABA)和玉米素核苷(ZR)的含量。iPA、DHZR和ZR是植物中最具活性的细胞分裂素(CTKs)。Ribeiro等[34]的试验结果表明,CO2浓度升高可提高赤霉素(GA)的含量,这与赤霉素(GA)在整合碳水化合物和氮代谢方面发挥类似的作用,是确定最佳生物量的基础。有研究表明,CO2加富可以提高ACC氧化酶的活性,增强1-氨基环丙烷-1-羧酸(ACC)转化为乙烯(ETH)的能力,从而提高乙烯(ETH)的含量[35]。本试验结果表明,CO2加富下,IAA、GA、CTKs及ETH的含量会增加,脱落ABA的含量会降低(图1),这与Teng等[36]在拟南芥上的研究结果一致,并认为引起这种变化的原因的是CO2加富影响相关基因的表达,如生长素(IAA)、细胞分裂素(CTKs)的响应因子在CO2加富下高度表达。CO2加富可以促进糖类的积累,从而加快呼吸速率,呼吸作用产生的ATP可转化为ADP从而调控生长素(IAA)的合成[37],同时ATP可作为底物合成异戊烯基腺苷-5'-三磷酸(iPTP)和异戊烯基腺苷-5'-二磷酸(iPDP),再经iPTP和iPDP羟基化合成玉米素核苷(ZR)等细胞分裂素(CTKs),从而增加细胞分裂素(CTKs)的含量[38]。也有人认为赤霉素(GA)含量的增加与被CO2诱导的Ca2+浓度的提升有关[36]。
干旱胁迫后,植物体内的激素平衡会被打破,最明显的一个特征就是脱落酸(ABA)含量的升高。研究表明,外源施加ABA可以被植物直接吸收,从而影响内源激素水平,黄杏等[39]研究结果表明,外源ABA可以提高甘蔗在低温胁迫脱落酸(ABA)的含量及ABA/GA、ABA/IAA、ABA/ZR的比值,降低生长素(IAA)、赤霉素(GA)和细胞分裂素的含量(CTKs),这是ABA提高甘蔗抗寒性的一个重要原因。在盐碱胁迫下,外源ABA对各激素的水平影响与在低温胁迫下一致,均是促进ABA的积累,降低IAA、GA、CTK的含量[40],这是因为外源ABA可作为合成内源ABA的替代物被植物直接吸收,从而提高内源ABA的含量,而ABA可与IAA、GA、CTK等生长类激素产生拮抗作用,从而抑制它们的合成[39]。本试验中,外源施加ABA可以增加干旱胁迫下黄瓜幼苗叶片中ABA和ETH的含量,降低IAA、GA和CTKs的含量(图1),这与前人在瓜尔豆[41]上的研究结果相似。这说明,干旱胁迫时,黄瓜幼苗叶片中的各激素之间可以相互促进,相互制约,通过减缓植物的生长、促进气孔关闭、降低蒸腾等使其达到新的平衡,从而适应干旱胁迫,减少损伤。
植物的生长发育过程,也是促进衰老的激素和抑制衰老的激素之间相互制约、相互平衡的过程,每种激素都有不同的效应,并以系统的方式调控植物衰老[42],而激素的比值变化能够揭示促进生长和抑制生长的激素在植物发育过程中的平衡[43],植物激素间的平衡是影响植物生长发育的重要因素,各种外源激素可通过改变内源激素的含量和平衡而发挥其作用[40]。前人研究结果表明,GA/ABA的比值与植物种子萌发及性别分化密切相关,IAA/ABA的比值与植物生长势密切相关,CTK/ABA则影响着气孔运动[44]。(IAA+GA+CTK)/ABA比值能反映植物的生长与休止状况,(IAA+GA+CTK)/ABA比值低,说明抑制型激素ABA含量占优势,植物处于休眠状态;促进型激素含量占优势时,( IAA+GA+CTK)/ABA比值高,植物生长旺盛[45]。本试验的结果在一定程度上符合这一结论,由图2可以看出,CO2加富处理显著提高了(IAA+GA+CTK)/ABA的比值,IAA能够促进细胞伸长,GA能够促进植物生长,CK能诱导细胞分裂,延缓衰老,而ABA对植物的生长势有抑制作用,所以该比值升高能够减弱抑制生长发育的作用,从而促进植物生长,因此CO2加富条件下,黄瓜幼苗的株高、茎粗、叶面积以及干鲜重都能得到增加,从而促进了生物量的积累。而外源施加ABA后,(IAA+GA+CTK)/ABA的比值会显著下降,这是因为外源施加ABA可以被植物直接吸收,ABA的含量会显著增加,而ABA与其他生长类激素之间是相互制约的关系,所以IAA、GA、CTK的含量下降,最终导致(IAA+GA+CTK)/ABA比值降低,所以,ABA对干旱胁迫下黄瓜幼苗的株高、叶面积并没有显著促进作用,此时,ABA的主要作用并不是体现在植物的生长状况上,而是体现在对植物的抗逆性上。当植物受到逆境胁迫时,ABA作为胁迫信号发挥正向调节作用[46]。所以,尽管Na2WO4处理下(IAA+GA+CTK)/ABA的比值很高,但该处理下黄瓜幼苗的生长状况最差。
4 结 论
综上所述,当黄瓜幼苗受到干旱胁迫时,CO2加富与外源喷施ABA可通过协同调控内源激素的含量及比值来平衡黄瓜幼苗的生长以适应干旱胁迫。
[1] Selda O, Melek E, Ertan Y,. Interactive effects of salinity and drought stress on photosynthetic characteristics and physiology of Tomato (L.) seedlings [J]. South African Journal of Botany, 2021,137:335-339
[2] Reitsema RE, Wolters JW, Stefan P,. Response of submerged macrophyte growth, morphology, chlorophyll content and nutrient stoichiometry to increased flow velocity and elevated CO2and dissolved organic carbon concentrations [J/OL]. Frontiers in Environmental Science, 2020.https://doi.org/10.3389/fenvs.2020.527801
[3] Kumar A, Nayak A, Sah R,. Effects of elevated CO2concentration on water productivity and antioxidant enzyme activities of rice (L.) under water deficit stress [J]. Field Crops Research, 2017,212:61-72
[4] 袁会敏,周健民,段增强,等.CO2浓度升高与增施钾肥对黄瓜生长的影响[J].土壤,2009,41(6):869-874
[5] 束秀玉.CO2加富对盐胁迫下西瓜幼苗生长及生理生化特性的影响[J].河南农业科学,2020,49(12):107-114
[6] 李仪曼,贺新蕊,李清明,等.CO2加富对干旱胁迫下黄瓜幼苗根系抗氧化系统的影响[J].植物生理学报,2019, 55(7):1011-1019
[7] Sharon BG, Orla D, Stephanie PK,. Intensifying drought eliminates the expected benefits of elevated Carbon Dioxide for Soybean [J]. Nature Plants, 2016,2(9):53-67
[8] 翟锡姣,刘婷婷,曹世宇,等.CO2加富及不同灌溉次数对菜豆生长及光合特性的影响[J].华北农学报,2020,35(2):88-97
[9] 李甜,张志鹏,侯雷平,等.CO2加富对甜瓜叶片全生命周期光合作用的影响[J].福建农业学报,2020,35(11):1198-1206
[10] 董彦红,刘彬彬,张旭,等.黄瓜幼苗非结构性碳水化合物代谢对干旱胁迫与CO2倍增的响应[J].应用生态学报,2015,26(1):53-60
[11] Murata Y, Mori IC, Munemasa S. Diverse stomatal signaling and the signal integration mechanism [J]. Annual Review of Plant Biology, 2015,66(1):369-392
[12] Engineer CB, Ghassemian M, Anderson JC,. Corrigendum: Carbonic Anhydrases, EPF2 and a novel protease mediate CO2control of stomatal development [J]. Nature, 2015,526(7573):246-250
[13] 李清明,刘彬彬,艾希珍.CO2浓度倍增对干旱胁迫下黄瓜幼苗膜脂过氧化及抗氧化系统的影响[J].生态学报,2010,(22):57-65
[14] Yu JJ, Du HM, Xu M,. Metabolic responses to heat stress under elevated atmospheric CO2concentration in a cool-season grass species [J]. Journal of the American Society for Horticultural Science, 2012,137(4):221-228
[15] 厉书豪,李曼,张文东,等.CO2加富对盐胁迫下黄瓜幼苗叶片光合特性及活性氧代谢的影响[J].生态学报, 2019,39(6):2122-2130
[16] Dodd IC, Davies WJ. Hormones and the regulation of water balance [M]. Netherlands: Springer, 2010
[17] Cutler SR, Rodriguez PL, Finkelstein RR,. Abscisic Acid: Emergence of a core signaling network [J]. Annual Review of Plant Biology, 2010,61(1):651-679
[18] Iuchi S, Kobayashi M, Taji T,. Regulation of drought tolerance by gene manipulation of 9-Cis -epoxycarotenoid dioxygenase, a key enzyme in Abscisic Acid biosynthesis in[J]. The Plant Journal, 2001,27(4):325-333
[19] Wang Y, Ma F, Li M,. Physiological responses of kiwifruit plants to exogenous ABA under drought conditions [J]. Plant Growth Regulation, 2011,64(1):63-74
[20] 陈露露,王秀峰,刘美,等.钙与脱落酸对干旱胁迫下黄瓜幼苗光合及相关酶活性的影响[J].应用生态学报,2016,27(12):3996-4002
[21] Fang L, Abdelhakim LOA, Hegelund JN,. ABA-mediated regulation of leaf and root hydraulic conductance in Tomato grown at elevated CO2is associated with altered gene expression of aquaporins [J]. Horticulture Research, 2019,6(1):657-667
[22] Li S, Li X, Wei Z,. ABA-mediated modulation of elevated CO2on stomatal response to drought [J]. Current Opinion in Plant Biology, 2020,56:174-180
[23] 龚建华,向军.黄瓜群体叶面积无破坏性速测方法研究[J].中国蔬菜,2001(4):7-9
[24] Ainsworth EA, Long SP. What have we learned from 15 years of eree-air CO2enrichment (face)? a meta-analytic review of the responses of photosynthesis, canopy properties and plant production to rising CO2[J]. New Phytologist, 2005,165(2):351-371
[25] Feihong L, Wenjie Y, Lang X,. Closing extra CO2into plants for simultaneous CO2fixation, drought stress alleviation and nutrient absorption enhancement [J]. Journal of CO2Utilization, 2020,42:1-10
[26] Pritchard SG, Rogers HH, Prior SA,. Elevated CO2and plant structure: a review [J]. Global Change Biology, 1999,5(7):807-837
[27] Gail T, J TP, Z ZF,. Spatial and temporal effects of free-air CO2enrichment (popface) on leaf growth, cell expansion, and cell production in a closed canopy of poplar [J]. Plant Physiology, 2003,131(1):175-185
[28] Makino A, Harada M, Sato T,. Growth and N allocation in rice plants under CO2enrichment [J]. Plant Physiology, 1997,115(1):199-203
[29] 李琬婷,宁朋,王菲,等.外源脱落酸对干旱胁迫下滇润楠幼苗生长及生理特性的影响[J].应用生态学报,2020,31(5):1543-1550
[30] Bari R, Jones JDG. Role of plant hormones in plant defence responses [J]. Plant Molecular Biology, 2009,69(4):473-488
[31] Kosová K, Prásil IT, Vítámvás P,. Complex phytohormone responses during the cold acclimation of two wheat cultivars differing in cold tolerance, winter samanta and spring sandra [J]. Journal of Plant Physiology, 2012,169(6):567-576
[32] 周晏起,卜庆雁.干旱胁迫下果树内源激素变化规律研究进展[J].北方果树,2011,163(3):1-4
[33] Li XM, Zhang LH, Ma LJ,. Elevated Carbon Dioxide and/or Ozone concentrations induce hormonal changes in[J]. Journal of Chemical Ecology, 2011,37(7):779-784
[34] Ribeiro GHPM, Araújo WL, Fernie AR,. Action of gibberellins on growth and metabolism ofplants associated with high concentration of Carbon Dioxide [J]. Plant Physiology, 2013,160(4):1781-1794
[35] Seneweera S, Aben SK, Basra AS,. Involvement of Ethylene in the morphological and developmental response of rice to elevated atmospheric CO2concentrations [J]. Plant Growth Regulation, 2003,39(2):143-153
[36] Teng N, Wang J, Chen T,. Elevated CO2induces physiological, biochemical and structural changes in leaves of[J]. New Phytologist, 2006,172(1):92-103
[37] 翟开恩,潘伟槐,叶晓帆,等.高等植物局部生长素合成的生物学功能及其调控机制[J].植物学报,2015,50(2):149-158
[38] 罗超,黄世文,王菡,等.细胞分裂素的生物合成、受体和信号转导的研究进展[J].特产研究,2011,33(2): 71-75
[39] 黄杏,陈明辉,杨丽涛,等.低温胁迫下外源ABA对甘蔗幼苗抗寒性及内源激素的影响[J].华中农业大学学报,2013,32(4):6-11
[40] 李红,李波,杨曌.外源ABA对苏打盐碱胁迫的紫花苜蓿内源激素含量的影响[J].黑龙江畜牧兽医,2020,594(6):103-106
[41] 周玲,魏小春,郑群,等.脱落酸与赤霉素对瓜尔豆叶片光合作用及内源激素的影响[J].作物杂志,2010,134(1):15-20
[42] Yong JW, Wong SC, Letham DS,. Effects of elevated [CO2] and Nitrogen nutrition on Cytokinins in the xylem sap and leaves of cotton [J]. Plant Physiology, 2000,124(2):767-780
[43] 梁艳,沈海龙,高美玲,等.红松种子发育过程中内源激素含量的动态变化[J].林业科学,2016,52(3):105-111
[44] 韩瑞宏,张亚光,田华,等.干旱胁迫下紫花苜蓿叶片几种内源激素的变化[J].华北农学报,2008(3):81-84
[45] 李雪梅,何兴元,陈玮,等.大气二氧化碳浓度升高对银杏叶片内源激素的影响[J].应用生态学报,2007(7):1420-1424
[46] 李玉梅,李建英,王根林,等.水分胁迫对大豆幼苗叶片内源激素的影响[J].大豆科学,2007(4):627-629
Effects of CO2Enrichment and Exogenous ABA on Growth and Contents of Endogenous Hormones of Cucumber Seedlings under Drought Stress
HE Xin-rui1, LI Qing-ming1,2*
1.271018,2.271018,
Hydroponic cucumber seedlings of 'Jinyou No.35' were used as test material, and PEG-6000 was added to the nutrient solution to simulate drought stress,the effects of CO2enrichment and exogenous ABA on growth and endogenous hormone levels of cucumber seedling under drought stress were studied.The results showed that CO2enrichment could significantly promote the growth of cucumber seedlings under drought stress, increase plant height, stem diameter, leaf area and dry and fresh weight, increase the content of auxin (IAA), cytokinin (CTK), gibberellin (GA), ethylene (ETH) and the ratio of (IAA+GA+CTK)/ABA while decrease abscisic acid (ABA) content; exogenous ABA significantly increased the stem diameter and fresh and dry weight of cucumber seedlings, increased the contents of ETH and ABA, but decreased the contents of IAA, CTK, GA and the ratio of (IAA+GA+CTK)/ABA.In conclusion, CO2enrichment and exogenous ABA can maintain the growth of cucumber seedlings under drought stress by regulating endogenous hormones.
Cucumber; drought stress; CO2enrichment; exogenous hormone
S642.2
A
1000-2324(2021)03-0352-06
2020-12-15
2021-02-14
国家自然基金(31872154;31902092)
贺新蕊(1996-),女,硕士研究生,研究方向:蔬菜生理生态. E-mail:1042888045@qq.com
Author for correspondence. E-mail:gslqm@163.com
