白刺属植物灌丛沙包生物量分布及预测
2021-07-28张正国辛智鸣张景波王学全菅凯敏马海峰黄雅茹李永华
张正国,董 雪,辛智鸣,张景波,王学全,菅凯敏,马海峰,黄雅茹,,李永华,5,6
(1. 中国林业科学研究院荒漠化研究所,北京 100091;2. 中国林业科学研究院沙漠林业实验中心,内蒙古 磴口 015200;3. 巴彦淖尔市造林技术推广站, 内蒙古 巴彦淖尔 015000;4. 巴彦淖尔市野生动植物和湿地保护中心, 内蒙古 巴彦淖尔 015000;5. 库姆塔格荒漠生态系统国家定位观测研究站, 甘肃 敦煌 736200;6. 甘肃敦煌荒漠生态系统国家定位观测研究站, 甘肃 敦煌 736200)
生物量不仅与植物生存适应能力及生产能力密切相关[1-4],而且是计算生物碳存储的重要参数[5]。生物量可以反映群落生态系统的生产力,从而可以间接说明生态系统的功能[6-7]。植物的生长、发育和繁殖对生物量有巨大的作用[8],植物对各营养器官生物量的分配策略是其对环境长期适应的结果,同时受到个体植株大小,以及环境和群落物种组成等因素的影响[9]。随着植物生物量研究的深入,植物个体生物量及其分配模式逐渐成为研究全球生态系统碳循环的重点内容之一[10-11]。
灌丛沙包广泛分布于西北干旱区,是植物与风沙相互作用的共同结果。植物在生长过程中阻拦沙子,被埋枝条能够生长不定根,从而在沙包内部和下部形成两层根系分布层[12]。风沙和植物的相互作用可能改变植物生长及生物量分布,使已有生物量预测模型难以适用于灌丛沙包的生物量预测。由于植物生物量采集破坏性强、工作量大、采集测定困难,尤其是根系生物量,一直是困扰植物生物量研究的难题。在此背景下,为更好地预测植物生物量,基于易测指标的植物生物量模型不断涌现[13-14]。目前,基于个体形态指标的生物量模型,已能够很好地预测乔木和灌木等单个物种或具有相似形态结构的多个物种的单株生物量及其器官生物量[13-15]。但是,有关灌丛沙包整株生物量及其器官生物量的预测模型报道却较少。
本研究分别在乌兰布和沙漠东北缘设置唐古特白刺灌丛沙包样地,在敦煌设置泡泡刺灌丛沙包样地,进行群落调查,在此基础上采用整体挖掘法,分类收获灌丛沙包上部枝叶、沙包内部枝与不定根及沙包下部根系,通过清洗、烘干、称重,获取对应生物量。与此同时,结合文献记录的数据,研究分析了白刺属植物灌丛沙包生物量分布特征并构建了预测模型。通过回归模型研究灌丛沙包生物量分布规律,探讨白刺属植物灌丛沙包各营养器官生物量分布策略,旨在为荒漠生态系统的灌丛沙包生物量及碳储量估算提供科学有效的方法,同时为定量评估荒漠生态系统固碳效益提供科学的数据支撑。
1 研究方法
1.1 数据获取及整理
本研究共收集72 个白刺属植物灌丛沙包生物量数据,数据收集区包括内蒙古磴口(中国林业科学研究院沙漠林业实验中心二场),甘肃敦煌(敦煌东湖边缘)、民勤和临泽4 个区域,灌丛沙包种群包括唐古特白刺和泡泡刺(表1)。其中,个人采集数据包括磴口唐古特白刺沙包27 个,敦煌泡泡刺沙包9 个[12,16]。生物量采集前,先调查记录灌丛沙包的长度、宽度和高度,沙包上部植株高度(沙包上部枝条高度,选择6 个枝条测量其长度,取平均值)及植株高度(沙包高 + 沙包上植株高度),而后采用推土机与人工相接的途径,整体挖掘、分拣、清洗、烘干获取植物生物量。生物量指标包括沙包上部、内部、下部生物量和总生物量;同时计算枝叶生物量(沙埋枝 + 沙包上部枝叶)和根系生物量(沙埋根系 + 沙包下部根系)。
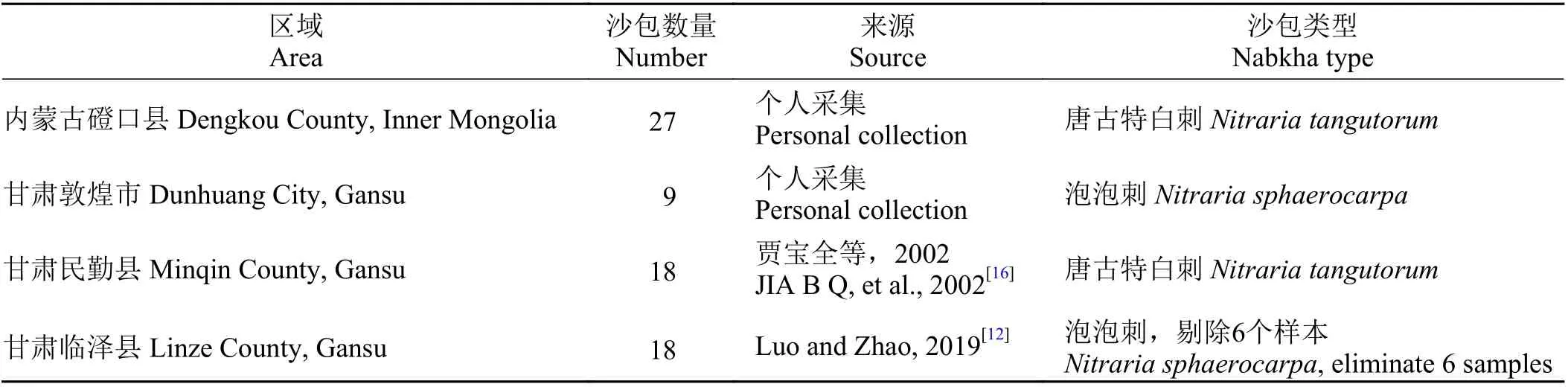
表1 采样区域、样本量及来源Table 1 Data acquisition location, sample source, and size
文献数据收集时,中文以“灌丛沙包”或“灌丛沙丘”与“生物量”为关键词,英文以“shrub sand dune”或“nabkha”或“vegetated dunes”与“biomass”为关键词,搜索文献。而后,分拣出包含唐古特白刺或泡泡刺及包含灌丛沙包上部、内部、下部生物量及沙埋根系、沙包下部根系生物量数据的文献。同时,根据数据采集方法,仅保留整理采用整体挖掘法获取的灌丛沙包生物量数据。最后民勤唐古特白刺沙包18 个,临泽泡泡刺沙包18 个。经过数据比对并与原作者交流,临泽6 个较大的灌丛沙包生物数据通过部分挖掘与推算获取,与予剔除。因此,本研究最后保留使用66 个数据。从收集数据可以看出,灌丛沙包的长与宽的变化超过20 倍,植株高度变化超过15 倍,灌丛沙包内部各组件生物量变化超过800 倍,最大达到了8 329 倍(表2)。

表2 灌丛沙包形态和各组件生物量Table 2 Shape of shrub nabkhas and the biomass of each component
1.2 数据分析
采用Excel 2016 和SPSS 17.0 软件对数据进行统计分析,本研究通过一元线性回归方程,分析了各组件生物量与总生物量的相关关系,以及各组件生物量与总生物量的比随沙包高度的变化特征;按沙包高度将数据分为5 个分级(沙包高度 < 0.5 m,0.5 m ≤ 沙包高度 < 1 m,1 m ≤ 沙包高度 < 1.5 m,1.5 m ≤ 沙包高度 < 2 m,沙包高度 ≥ 2 m;每个级别样本量分别为28、19、9、6 和4 个),运用方差分析比较了5 个级别的灌丛沙包各组件生物量与总生物量比的差异。一元线性回归分析结果显示,在水平维度上,白刺属植物灌丛沙包长、宽之间存在密切的线性正相关关系(沙包宽 = 0.93 × 沙包长,R2=0.96,P< 0.01)。因此,本研究选取代表水平维度的沙包长及代表垂直维度的植株高度(沙包高 + 沙包上枝条平均高度)为输入因子,通过自然对数处理,应用一元线性回归模拟,分析了沙包长度、植株高度与各组件生物量的关系;应用二元线性回归模型,以沙包长度、植株高度为输入变量,预测了灌丛沙包各组件生物量。
2 结果与分析
2.1 白刺属植物灌丛沙包生物量分布特征
一元线性回归分析显示,总生物量与沙包上部生物量、沙包内部生物量、沙包下部生物量、枝叶生物量及根生物量均具有密切的相关关系,总生物量对各组件生物量的解释度在72%~97% (图1)。单株总生物量每增加1 kg,它们分别对应增加0.31、0.57、0.12、0.44 和0.56 kg。
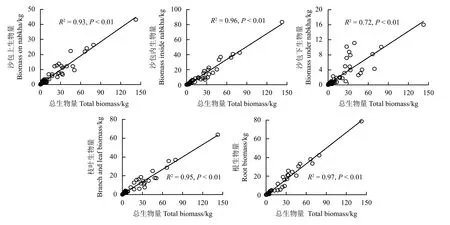
图1 灌丛沙包各组件生物量与单株总生物量的关系Figure 1 Relationship between the biomass of each component and the nabkha total biomass
随灌丛沙包高度的增加,沙包上部生物量、沙包内部生物量、沙包下部生物量、根生物量及沙包内根生物量与总生物量的比值也发生改变(图2)。总体来看,随灌丛沙包高度增加,沙包上部生物量占比呈现减少趋势,沙包内部生物量占比呈现增加趋势,而沙包下生物量占比没有明显变化趋势。高度< 0.5 m 至≥ 2 m 的沙包,沙包上生物量占总生物量的比例从大于40%下降至25%左右,沙包内生物量占总生物量的比例从40%左右增加至60%以上,而沙包下生物量占比在8.5%~19.2%之间波动。另外,随沙包高度增加,沙包内根与总体根系生物量占沙包灌丛总生物量的比例呈增加趋势。高度<0.5 m 至≥ 2 m 的沙包,沙包内根系生物量占沙包灌丛总生物量的比例从15%左右增加至50%左右,总体根系生物量占沙包灌丛总生物量的比例从35%左右增加至超过60%。
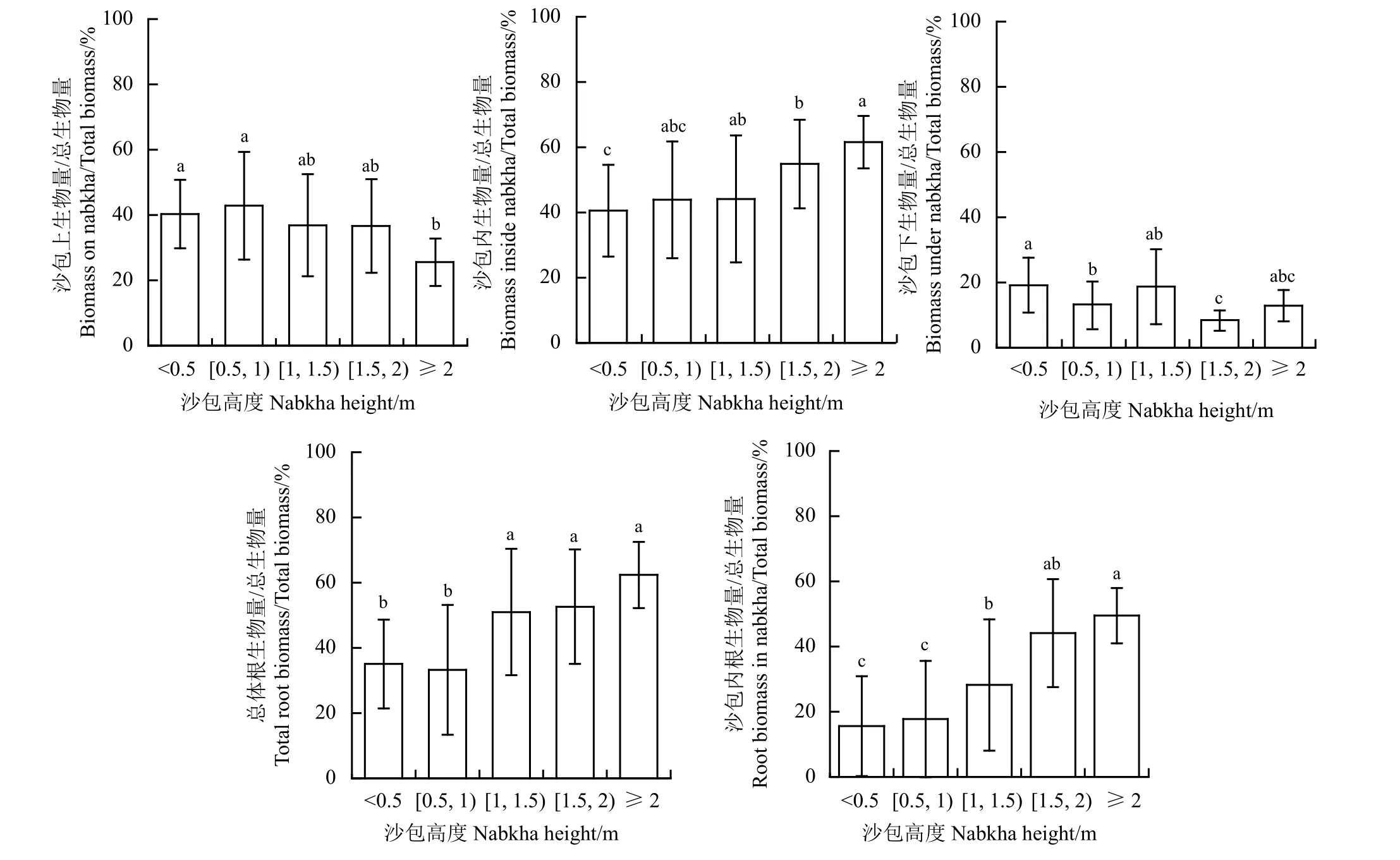
图2 不同大小灌丛沙包各组件生物量分布特征Figure 2 Biomass allocation characteristics of different components of nabkhas of different sizes
2.2 白刺属植物灌丛沙包生物量预测
自然对数处理后,沙包上部生物量、沙包内部生物量、沙包下部生物量、枝叶生物量、根系生物量、总生物量与植株高度、沙包长度呈现密切的线性相关关系(图3、图4)。植株高度变化能够解释各组件生物量及总生物量变化的76%~88% (图3),沙包长度变化能够解释各组件生物量及总生物量变化的76%~92% (图4)。联合植株高度和沙包长度,通过二元一次线性回归模型能够进一步提高灌丛沙包各组件生物量及总生物量的预测能力。拟合结果显示,植株高度和沙包长度对高灌丛沙包各组件生物量及总生物量的解释度达到84%~94%(表3)。
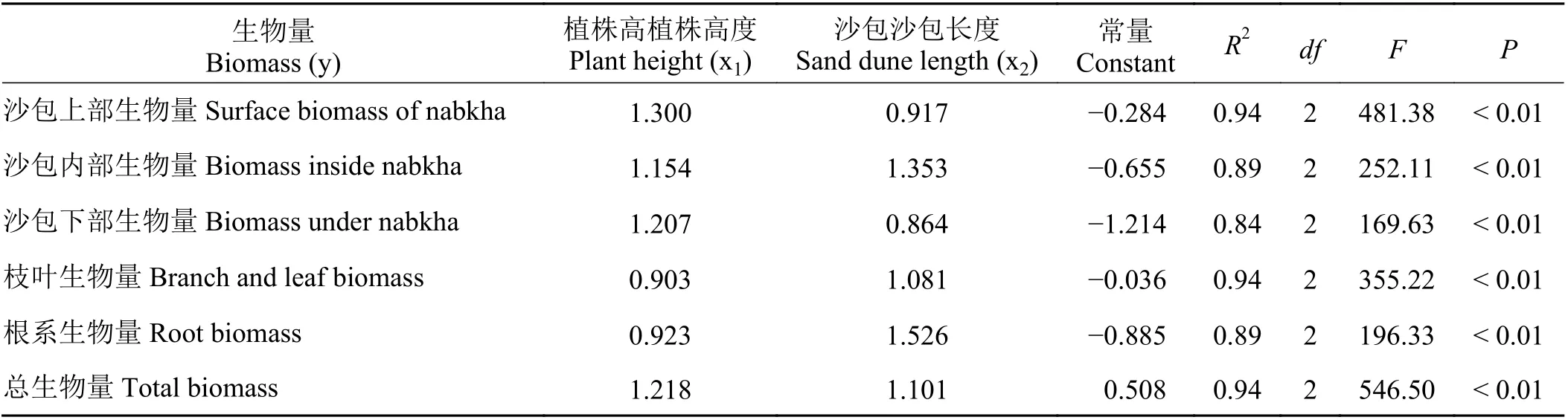
表3 生物量预测模型Table 3 Biomass prediction model
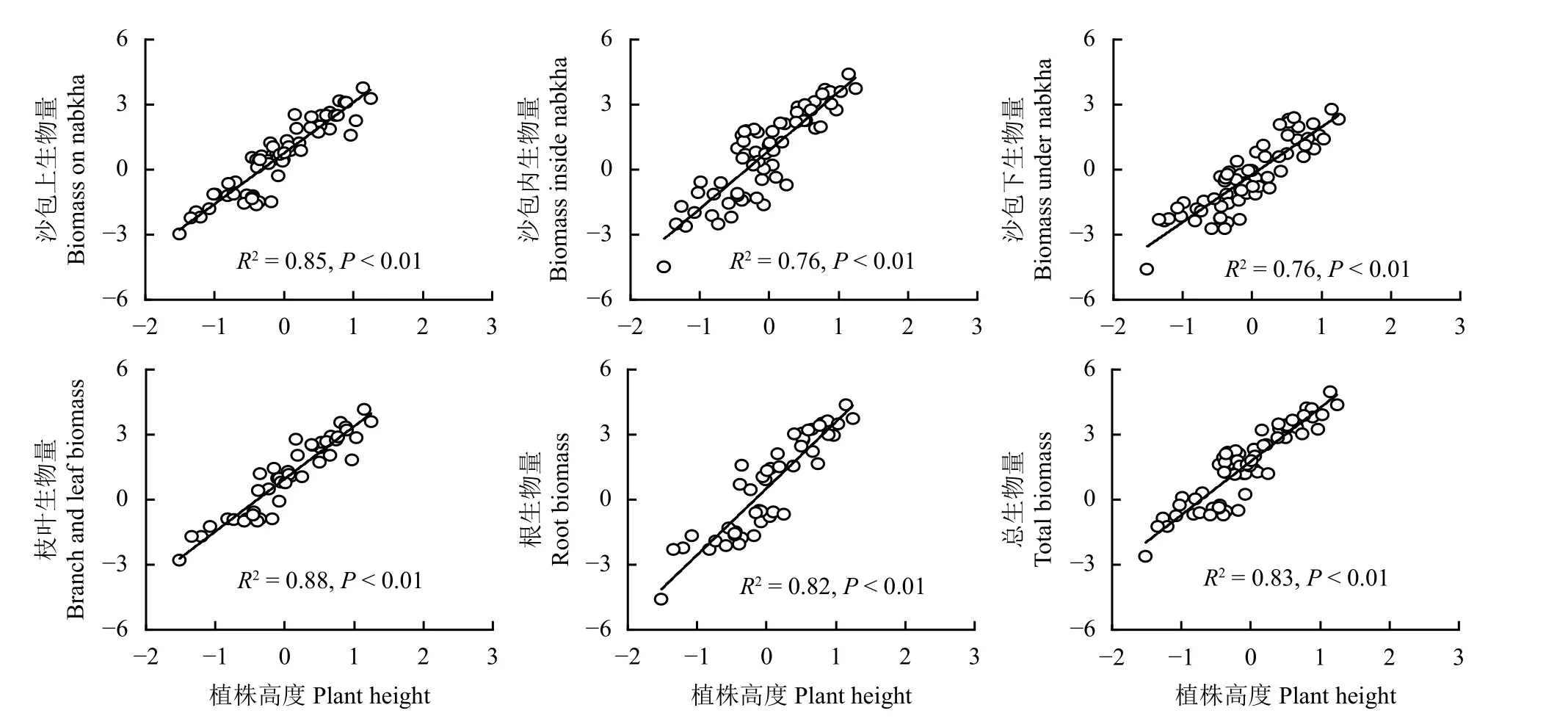
图3 灌丛沙包生物量与植株高度的相关关系Figure 3 Correlation between nabkha biomass and plant height
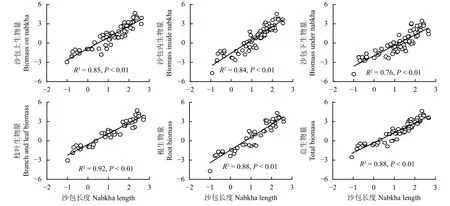
图4 灌丛沙包生物量与沙包长度的相关关系Figure 4 Correlation between nabkha biomass and nabkha length
3 讨论
3.1 白刺属植物灌丛沙包生物量预测
对于基径不明显或无法测定的灌木,常见生物量模型多通过植株高度和冠层长度、宽度(或冠层直径)预测灌木单株及各组件生物量[6-7,16]。在生物量模型的构建与预测中,自变量的选取是构建回归模型的关键[17-18]。大量研究结果表明,不同灌丛由于形态结构的差异,导致最优生物量预测模型的自变量和最优方程并不完全相同;但是,绝大多数的灌木生物量与基径和株高的复合因子间或者冠幅和株高的复合因子间会存在显著相关关系[17-20]。基于已有研究成果,本研究选用沙包长度和灌丛高度反映灌丛沙包水平和垂直方向生长的形态指标,通过自然对数处理,建立了灌丛沙包整株生物量及各组件生物量模型。该模型能够解释灌丛沙包总生物变化的94%,对其他组件生物量的解释度达到了84%~94%,模拟结果优于已发表的同类型模型模拟结果[16]。利用树木易测的指标因子构建生长回归方程来推算植物生物量,该方法简单、破坏性小且估算迅速,同时生物量数据可以进行外推,从而可以实施野外跟踪调查研究[21-22]。本研究中所有的生长方程都达到了极显著水平(P< 0.01),对白刺属植物灌丛沙包生物量变化的解释程度平均为90.6%。
3.2 白刺属植物灌丛沙包生物量分布
植物生物量分配格局的差异是其形态结构和生理特征共同作用的结果;同时,其也反映了植物对生存环境要素的响应规律及其生长策略[9,23]。植物可以通过调节生物量在各器官的分配格局来适应外界环境,而且在不同的生境下分配模式不同,从而更好地利用限制性资源维持自身的生存与生长[24-25]。唐古特白刺的根系生物量最大,这与其具有较强的抗旱、抗寒和耐盐碱的特性相关,说明其萌发力强,根系发达。个体大小、环境以及遗传性状等均影响植物生物量分配[4,6,26-27]。全球数据分析显示,小型植物叶与茎、叶与根的生物量比更高[4]。随着总生物量的增加,叶、根生物量占总生物量的比例明显降低,茎生物量占总生物量的比例显著增加[4]。本研究没有单独统计叶生物量及其占比,但考虑灌丛沙包上生物量随高度增加而降低,可以推断白刺属植物灌丛叶生物量占总生物量的比例随灌丛沙包高度增加而降低(图2)。但是,根系生物量占总生物量的比例随灌丛沙包高度的增加而增加(图2),出现这一结果可能与植株机械稳定及生境变化相关。通常,直立植物在生长过程中需要不断强化支撑作用,从而确保植株机械稳定性[13-14]。因此,随着植物总生物量的增加,茎干生物量占比会不断增加[3]。然而,对于沙包上生长的丛生植物,有了沙子外力支撑后,基本不需要额外的支撑,这为植物向其他器官投资更多生物量提供了可能。另外,根系是植物从土壤中吸收和输送水分和养分的重要器官,同时发挥着固定植株的作用[28-31]。干旱区土壤水分供给能力有限,白刺属植物灌丛生长过程中需水量会不断增加,加大了供需矛盾。这可能是植物提升水分捕获器官(根系)投资比例的重要原因。沙包内部根系投资比例增速远远高于沙包下部根系,这可能与沙包土壤较为疏松,水分更容易入渗存储,根系更容易从沙包内部获取较多的土壤水分有关[19,31]。
4 结论
随着沙包高度增加,白刺属植物灌丛沙包上部生物量占比呈现减少趋势,而沙包内部生物量、沙包内根系及总根系生物量占总生物量比例均具有增加趋势,但是沙包下部生物量占总生物量比例变化趋势不明显。同时,各组件生物量与总生物量具有良好的线性正相关关系,总生物量对各组件生物量的解释度在72%~97%。通过灌丛沙包长度、植株高度两个指标能够很好地预测白刺属植物灌丛沙包各组件生物量和总生物量,二者对高灌丛沙包各组件生物量及总生物量的解释度达到89%~94%。
