8个酿酒葡萄品种的抗寒性比较
2021-07-24贾金辉刘慧纯蔡智军田晓玲张海涛
贾金辉,徐 凌,刘慧纯,蔡智军,田晓玲,张海涛
(辽宁农业职业技术学院,营口115009)
随着我国酿酒葡萄的发展,辽宁省也开始出现葡萄酒厂,比如本溪桓仁有五女山酒厂,也有张裕的分厂,但葡萄酒业在辽宁省发展较慢,最主要的原因是酿酒葡萄品种大多数从国外引进,抗寒性较差,很容易受到冻害,为了防止冻害,需要进行埋土防寒,增加成本,耗时耗力[1]。所以,对酿酒葡萄品种的抗寒性研究意义重大,本试验以8 个酿酒葡萄品种作为材料,通过对枝条的相对电导率、芽段萌发率、可溶性糖含量、游离脯氨酸含量、丙二醛含量,根系的过冷能力、半致死温度、平均隶属度值等指标进行相关性分析[2],得出酿酒葡萄品种抗寒性的判断依据,为东北寒冷地区的酿酒葡萄种植与发展提供一定的参考。
1 材料与方法
1.1 试验材料
试验材料取自辽宁农业职业技术学院酿酒葡萄园(位于辽宁省营口市熊岳镇),8 个品种分别为‘双丰’‘左优红’‘北红’‘北醇’‘双红’‘北冰红’‘威代尔’‘公酿一号’。2018 年11 月剪取枝条和根系,每个品种随机选15 株树,每株随机选2~3 根长势良好的1 年生枝条和根系,枝条长度25~30 cm,根系直径2~3 mm,纯净水清洗干净,滤纸吸干枝条和根系的表面水分,将其封入样本袋内待测[3]。
1.2 试验方法
将供试枝条一部分置于4 ℃作为对照,其余分别在-10~-35 ℃每隔-5 ℃设置温度梯度进行冷冻处理,并在达到设定温度后保持15 h,再升至4 ℃保持1 d。然后随机取样进行相关指标的测定,相对电导率采用电导仪测定,芽段萌发率采用比值法测定,可溶性糖含量采用蒽酮法测定,丙二醛含量采用硫代巴比妥酸法测定,脯氨酸含量采用茚三酮比色法测定[4]。
将供试根系放入人工霜冻试验箱内,并将温度传感器一端固定在根系表皮上,另一端与计算机和数据采集系统连接,设置计算机连续自动记录数据,分析根系表面温度变化。设置人工霜冻试验箱从室温降至-13 ℃,以3 ℃/h 进行降温。绘制出温度变化曲线,可确定根系的结冰点和过冷却点。
根系的过冷能力=过冷却点温度-结冰点温度[5]
1.3 数据处理和分析
试验数据采用Excel 2016 和SPSS 15.10 软件进行整理和分析。
2 结果与分析
2.1 枝条在低温下的变化
2.1.1 枝条的相对电导率在低温下的变化
从表1 可以看出,随着温度的降低8 个品种枝条的相对电导率都增加。-20 ℃时,‘公酿一号’的相对电导率大于40%,且极显著高于其他品种,由此可知‘公酿一号’的抗寒性最弱[6]。-25 ℃时,‘北冰红’‘威代尔’的相对电导率均大于50%,均极显著高于除‘公酿一号’外的其他5 个品种,且其他5 个品种的相对电导率差异不显著,说明‘北冰红’‘威代尔’的抗寒性比除‘公酿一号’外的其他5 个品种弱。当温度降至-30 ℃时,‘北醇’‘北红’‘双红’‘左优红’的相对电导率均大于50%,且均极显著高于‘双丰’,说明‘双丰’的抗寒性最强[7]。

表1 8 个品种枝条在各低温处理下的相对电导率
2.1.2 枝条的芽段萌发率在低温下的变化
从表2 可以看出,随着温度的降低8 个品种枝条的芽段萌发率都降低,且差异显著。4 ℃时,8个品种枝条的芽段萌发率都大于90%,且差异不显著,说明8 个品种的枝条芽段健康良好[8]。当-10 ℃及更低温度时,‘公酿一号’的芽段萌发率极显著低于其他品种,故‘公酿一号’的抗寒性最弱。除‘公酿一号’外,‘北冰红’和‘威代尔’的芽段萌发率均极显著低于其他品种,且二者间差异不显著,说明‘北冰红’和‘威代尔’的抗寒性比‘公酿一号’强,比其他品种弱。在-15 ℃及更低温度时,‘左优红’‘北红’‘北醇’‘双红’的芽段萌发率均极显著低于‘双丰’,说明‘双丰’的抗寒性最强。

表2 8 个品种枝条在各低温处理下的芽段萌发率
2.1.3 枝条的可溶性糖含量在低温下的变化
由表3 可知,随着温度降低8 个品种枝条的可溶性糖含量都先升高后降低。‘公酿一号’在-10~-15 ℃的可溶性糖含量极显著低于其他品种,但随温度降低‘公酿一号’受低温冻害影响可溶性糖含量突然增加,其抗寒性最差[9]。在-10 ℃及更低温度(-30 ℃除外)时,‘北醇’‘双红’‘北冰红’‘威代尔’可溶性糖含量极显著低于除‘公酿一号’外的其他品种,抗寒性居中。在-10~-25 ℃,‘北红’和‘左优红’的可溶性糖含量差异不显著。
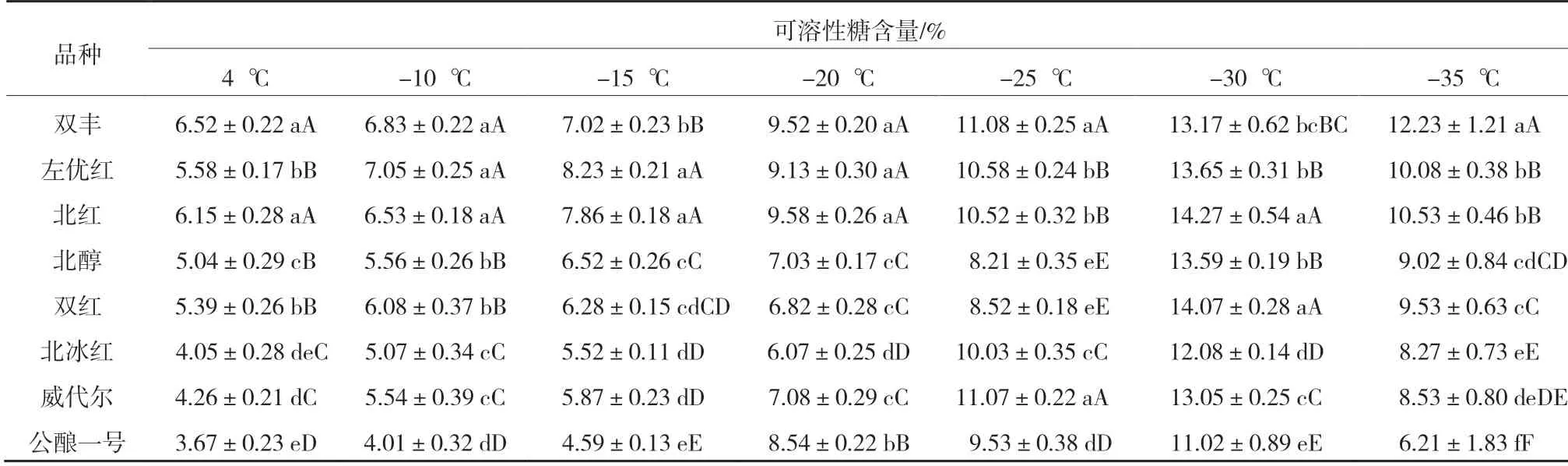
表3 8 个品种枝条在各低温处理下的可溶性糖含量
2.1.4 枝条的游离脯氨酸含量在低温下的变化
由表4 可知,8 个品种枝条的游离脯氨酸含量除‘双丰’随温度的降低持续升高外其他品种均先升高后降低,说明‘双丰’抗寒性最强。-25 ℃时‘公酿一号’的游离脯氨酸含量最低,且极显著低于其他品种,抗寒性最弱。-30 ℃时‘北冰红’和‘威代尔’的游离脯氨酸含量极显著低于除‘公酿一号’外的其他品种,说明‘北冰红’和‘威代尔’的抗寒性较弱。在-10~-25 ℃,‘北红’‘北醇’ ‘双红’的游离脯氨酸含量无显著差异。
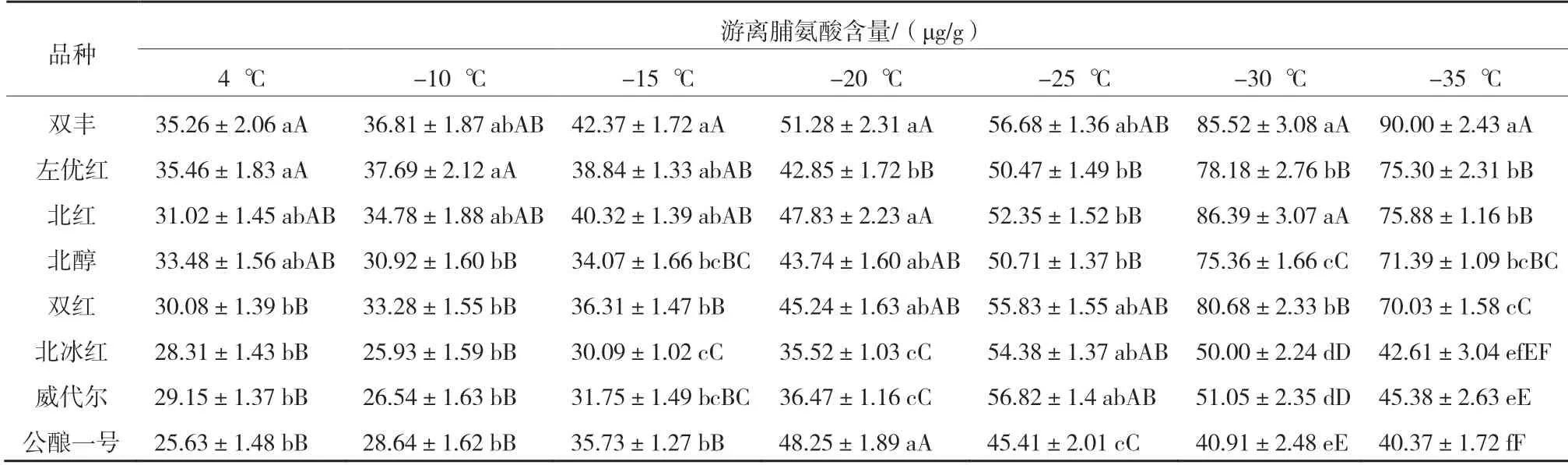
表4 8 个品种枝条在各低温处理下的游离脯氨酸含量
2.1.5 枝条的丙二醛含量在低温下的变化
从表5 可以看出,随着温度的降低8 个品种枝条的丙二醛含量除‘双丰’持续升高外其他品种均先升高后降低。‘公酿一号’在-20 ℃时丙二醛含量最高,极显著高于其他品种,抗寒性最弱[10]。在-25 ℃时‘北冰红’和‘威代尔’的丙二醛含量较高,极显著高于其他品种,抗寒性较弱。‘左优红’‘北红’‘北醇’‘双红’在-30 ℃时丙二醛含量均达到最高值,‘双丰’则在-35 ℃时丙二醛含量达到最高值。

表5 8 个品种枝条在各低温处理下的丙二醛含量
2.2 根系在低温下的变化
2.2.1 根系的过冷却点与结冰点
研究表明葡萄根系组织抗寒性与过冷却点呈负相关[5]。0 ℃以下时,根系组织细胞为过冷却的状态,此状态下的细胞内水分仍呈液态,不会受到冰冻伤害。所以,抗寒性越强的根系,其过冷却点就越低。由表6 可知,‘双丰’根系的过冷却点最低,抗寒性最强,能在-8.5 ℃中存活,其次是‘北红’‘左优红’‘双红’‘北醇’根系能在-7.8~-7.1 ℃中存活,‘北冰红’和‘威代尔’根系则能在-5.9~-5.8 ℃中存活,‘公酿一号’根系的过冷却点最高,能在-5.1 ℃中存活。

表6 8 个品种根系的过冷却点与结冰点
2.2.2 根系的过冷能力
葡萄根系组织的过冷能力可用结冰点与过冷却点温度差来表示,过冷能力越强,温度差越大[5]。由图1 可知,‘双丰’的过冷能力最高,抗寒性最强;‘公酿一号’的过冷能力最低,抗寒性最弱;其他品种的过冷能力由高到低依次为‘北红’‘左优红’‘双红’‘北醇’‘威代尔’‘北冰红’。此抗寒能力强弱排序基本与枝条、叶片得出的结果一致。

图1 8 个品种根系的过冷能力
2.3 抗寒性的综合评价
2.3.1 建立Logistic 方程和求解半致死温度
研究表明温度与相对电导率之间的关系为曲线[7],Logistic 方程也是曲线方程,故能用Logistic方程进行拟合。y=K/(1+ae-bx)为Logistic 方程,LT50=-lna/b 为半致死温度即曲线的拐点[11-12]。
根据8 个葡萄品种枝条在预设温度下的相对电导率建立Logistic 方程、求解半致死温度LT50和拟合度(R2),见表7。从表7 可知,‘双丰’半致死温度约为-32.9 ℃,极显著低于其他品种,抗寒性最强。‘左优红’‘北红’‘北醇’‘双红’4 个品种的半致死温度在-27.5~-25.4 ℃。‘北冰红’和‘威代尔’的半致死温度在-22.0 ℃左右。‘公酿一号’的半致死温度约为-18.4 ℃,极显著高于其他品种,抗寒性最弱。

表7 8 个品种的半致死温度
2.3.2 利用隶属函数法求解抗寒性强弱
研究表明平均隶属度与抗寒性有相关性[13],抗寒能力越弱的平均隶属度越小。隶属函数方程为X(u)=(X-Xmin)/(Xmax-Xmin)或X(u)=1-[(X-Xmin)/(Xmax-Xmin)]。利用此方程求解8 个品种的抗寒性位次。由表8 可知,8 个品种抗寒性由弱到强排序为‘公酿一号’<‘北冰红’<‘威代尔’<‘北醇’<‘双红’<‘左优红’<‘北红’<‘双丰’。抗寒性结果与半致死温度一致。

表8 8 个品种的隶属度值
2.3.3 抗寒各指标的相关性分析
为了研究抗寒性指标间的关联程度,对8 个品种的枝条、根系各指标、半致死温度和平均隶属度值进行相关性分析[14]。由表9 可知,与其他各指标的相关系数均达到极显著水平的有平均隶属度值、半致死温度和根系的过冷能力,而其他各指标之间的相关性较差,因此可以用平均隶属度值、半致死温度和根系的过冷能力评价葡萄抗寒性的强弱[15]。

表9 各指标间的相关性分析
3 讨论与结论
植物细胞膜的透性与环境温度紧密相关,温度较低时,膜透性将随之改变,功能逐渐降低甚至完全丧失,这会导致细胞内的溶液向外渗出。渗出的溶液越多,电导率就越高。抗寒性和细胞膜的透性存在负相关性,通过测定枝条的电导率来判断枝条的抗寒性是一种经典的试验方法[16-18]。Logistic 方程是典型的S 曲线方程,相对电导率与温度之间的关系曲线能较好地用Logistic 方程进行拟合[19-20]。曲线的拐点即为半致死温度[21]。半致死温度可以作为评价抗寒性的一项重要指标[22-23]。研究表明,植物的抗寒能力与枝条的芽段萌发率呈正相关性,植物的抗寒能力越强,其在低温下的芽段萌发率越高。许多研究认为,植物受到低温胁迫时,体内的可溶性糖会发生变化,随之改变细胞的渗透浓度,温度越低,可溶性糖含量会越高,使渗透浓度增加,导致冰点随之降低[24-25]。艾琳、王敏等研究表明,在常见氨基酸中游离脯氨酸的溶解度通常比较大,游离脯氨酸又是水溶性良好的氨基酸,所以它可以改变细胞的渗透压,来保护原生质体[26-27]。当植物受到低温冻害时,枝条上的游离脯氨酸含量会随之上升,增大细胞内蛋白质胶体亲水面积,增加植物的抗寒能力[28-29]。尤超等研究表明,植物在低温环境下过多的活性氧容易产生膜脂过氧化,引发过氧化产物丙二醛的大量积累,因此可以通过测定丙二醛含量来判断植物的抗寒能力[30]。
本试验通过测定一些抗寒性生理指标,分析各项指标与抗寒性的相关性得知:枝条的芽段萌发率、游离脯氨酸含量、可溶性糖含量以及根系的过冷能力与抗寒性呈正相关;枝条的丙二醛含量和相对电导率与抗寒性呈负相关。枝条的芽段萌发率、相对电导率、可溶性糖含量、丙二醛含量、游离脯氨酸含量与其他抗寒指标的相关性呈显著水平,平均隶属度值、半致死温度和根系的过冷能力与其他抗寒指标的相关性呈极显著水平,因此可以用后3项指标来评价抗寒性强弱。利用隶属函数法、半致死温度和根系的过冷能力比较8 个葡萄品种的抗寒性,抗寒能力由弱到强一致表现为‘公酿一号’<‘北冰红’<‘威代尔’<‘北醇’<‘双红’<‘左优红’<‘北红’<‘双丰’。
