基于根系与繁殖特征的风轮菜自我身份识别研究
2021-07-22宋会兴
刁 雪,宋会兴
(四川农业大学 风景园林学院,四川 成都)
【研究意义】克隆植物通过芽、枝条等无性繁殖方式产生出具有相同基因型的克隆分株,并能通过分株间的克隆整合和克隆分工等策略分担环境风险[1],实现对异质生境的自我适应,最大化利用环境资源并在群落生长中占取优势,使其在贫瘠地区的植被覆盖、群落演替演化及生态系统功能维持方面发挥着重要作用。克隆植物分株间的克隆整合和分工等策略是通过间隔子(Spacer,匍匐茎或根状茎等)来实现的[2],而动物啃食、践踏以及人类活动可能会破坏克隆植物间隔子的连接,使得植物面临一株与自己基因型完全一致的邻体植株。根据生态位分化假说[3],具有相似资源需求的植物之间会产生较大竞争。克隆分株间相同的基因型使其个体发展具有相似的生态位需求,以及分株个体间相邻的生存空间提供了潜在的种内竞争可能性,而个体的最大化竞争促使植物将获得的养分资源投入到营养生长中去,例如,像生物量和株高等竞争性状,往往会形成“公地悲剧(Tragedy of the commons)”而导致群体适合度的降低[4]。近年来有研究表明,植物间存在着亲缘或自我/非我的身份识别机制[5-7],植物个体间的遗传距离能通过影响自身的表型可塑性来使其获得更大的广义适合度,且关系越近,这种“利他”行为效益越强[8-9]。因此,研究克隆植物分株间的自我身份识别对于理解克隆植物种群构建与进化发展具有重要理论意义。【前人研究进展】Roiloa 等[10]对欧亚活血丹(Glechoma hederacea)和莫邪菊(Carpobrotus edulis)进行研究发现,欧亚活血丹亲缘分株间根系产生回避行为,避免竞争;莫邪菊亲缘分株根系生物量降低,但总生物量增加,2个物种都表现出了亲缘间互助的现象。大叶落地生根(Kalanchoe daigremontianaHamet&Perrier)则通过根系表现出自我识别的能力,减少了克隆分株间的竞争对抗,根系生物量降低的同时促进克隆繁殖[11]。但也有研究得到了相反的结果,如野草莓(Fragaria vesca)的亲缘分株间表现出了更强的竞争能力[10,12]。【本研究切入点】植物通过根系觅食获取生长所需养分,通过自身养分分配格局适应环境资源的分布,影响种群的适合度,还能通过地下根系的接触及分布感知邻域植物的存在[13-17],因此根系是植物身份识别机制实现的主要途径之一。研究假设植物根系特征是植物对邻域个体亲缘关系的响应,这种响应会进一步影响种群的繁殖特征。【拟解决的关键问题】以草坪常见杂草风轮菜为研究对象,进行根盒栽培实验,通过研究其根系行为与繁殖性状来探究:(1)克隆植物风轮菜分株间是否存在自我与非我的身份识别响应?(2)如果存在身份识别响应,是否会对风轮菜克隆分株的有性生殖产生影响?
1 材料与方法
1.1 试验材料
风轮菜(Clinopodium chinense)为唇形科、风轮菜属多年生草本植物,匍匐茎,节多分枝,基部匍匐生根,上部抬升,是典型的克隆植物。广泛分布于中国黄河流域及以南地区,也是日本城市草坪、森林生态系统中常见的杂草[18]。本试验所用风轮菜取自四川农业大学成都校区(30°41'N,103°49'E)周边弃荒地。
1.2 试验设计
2019 年3 月,取大小、长势一致的风轮菜分株段(高约7~8 cm,直径约为3 mm)于花盆内,于室外恢复生长10 d 后选取生长状况良好的风轮菜分株备用。试验设置3 种处理(图1):匍匐茎保持连接的分株对为连接组(记作L)、匍匐茎切断的分株对为自我组(2 个分株来自于同一植株,记作S),来自不同基株(基株间距离超过100 m)的分株对为非我组(记作NS)。每个分株对移栽至定制的透明PVC 根盒内,根盒内部长、宽、高尺寸分别为30,3,10 cm。根盒内栽培基质为原生地土与干净河沙按体积比1∶1 混合均匀构成。将根盒用黑色塑料袋包裹后露天栽培,栽培基质表面干燥时则每个处理等量浇水。每天观察,记录。在风轮菜生长至盛花期(种植38 d 后)时收获,测定相应指标。每处理10 个重复。

图1 试验设计示意图Fig.1 The diagram of experiment design
1.3 指标测定
根据试验观察,栽培的风轮菜在栽培20 d 后开始萌生花序,统计单株植物花序数,此后每隔6 d 进行一次花序数统计。在植物收获后,记录单株花序轮数以及最大轮间距,并通过根盒观察每株植物的根系分布(图2),测量植株可视根系两侧最大角度值,即α、β角[19],并按照如下公式计算角度差值比ADI。

图2 根盒分区与植物根系角度示意Fig.2 The division of root box and the angle of plant root system

当角度差值比ADI>0,表明两株植物根系趋于回避;当ADI<0,表明两株植物根系趋于竞争;当ADI=0,表明两株植物根系间无明显相互作用。
植物收获时,参考Schmid 等[19]的方法进行水平根系分布指数RDI的测量,公式如下:

MN1、MN2分别为分布于N1、N2 非共生区域的根系生物量,MC为分布于共生区域的根系生物量。RDI>1,表明两株植物根系背向生长,趋于回避;RDI<1,表明两株植物根系趋于相向生长;当ADI=1,表明两株植物根系分布趋于平衡。用Epson扫描仪记录图像并用WinRHIZO 软件测量相应根系参数,并利用根长(L,cm)和根体积(V,cm3)计算根长密度RLD=L/V[20-21],此外,比根长SRL=根长(cm)/根生物量(mg)。根系活力采用TTC法测定。
1.4 数据处理
用Excel 2016进行数据记录与整理,并运用SPSS完成数据分析,运用Origin 2018完成图表制作。符合正态分布与方差齐性的各处理组数据采用单因素方差分析,并采用LSD 法对数据进行多重比较。当数据不满足正态分布与方差齐性时,采用独立样本Kruskal-Wallis检验法。
2 结果与分析
2.1 风轮菜根系特征对不同分株邻体的响应
由表1 可知,根系平均直径、ADI 与根系活力在处理组间差异显著。连接组的风轮菜根平均直径显著小于另外两组,非我组的该指标最大(图3B);该趋势同样体现在ADI 上,但后两者间没有达到差异显著性,所有处理组的ADI 都大于0,连接组ADI 值接近于0(图3G);连接组拥有最强的根系活力,远高于其他处理组(图3I)。其余的根系相关指标在组间均没有达到差异显著性,但非我组的根表面积显著高于其他两组(图3A)。
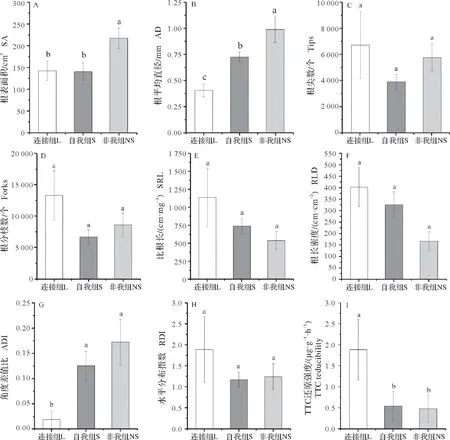
图3 不同处理组风轮菜各指标比较Fig.3 The comparation of traits in different treatments of Clinopodium chinense
2.2 风轮菜花序特征对不同分株邻体的响应
不同处理的风轮菜在花序轮数与花序数上表现出了差异显著性。由图4 可知,自移栽后,各处理组花序数量随时间而变化,但增量不同。风轮菜连接组花序数量显著高于其他处理组,自我组的花序数量在移栽后20~26 d内超过了非我组,并在32 d左右二者达到显著差异,但之后自我组花序数量增幅减小,非我组花序数上升,两组间差距变小。相同地,由图5B 可知,连接组花序轮数显著高于另外两组。而在轮间最大间距指标上,组间无显著差异(图5C),风轮菜的比叶面积指标在组间无显著差异(表1),其它组由大到小依次为连接组、自我组和非我组(图5A)。

图4 移栽后不同组间风轮菜花序数量变化Fig.4 The number of inflorescences of Clinopodium chinense in different groups changed after transplanting

表1 不同处理组风轮菜各指标比较Tab.1 The comparation of traits in different treatments of Clinopodium chinense
3 讨论与结论
根系是植物获取维持自身生长所需养分与水分的首要器官。根系形态特征及其空间分布是植物获取资源能力的体现,而细根数量更是养分获取的关键,反映了植物在异质性环境中所采取的策略[22]。研究中,连接组的风轮菜根平均直径显著小于其他处理组的,在根尖数、分枝数上占有优势,在表征根系收益和花费关系的比根长以及能够反映土壤资源有效性的根长密度指标[23]方面也表现出优势,体现了连接组风轮菜具有最强的根系活力(图3I)。ADI 与RDI 值则反映连接组分株外侧(回避邻近植株方向)根系生物量密度大于内侧,意味着连接组分株根系倾向于将觅食重心置于外侧,避免了连接分株间竞争的策略。连接组分株根系的这些特征可能与试验所用的根盒内资源有限有关。在资源受限条件下植物细根尽管寿命短,但其快速增殖-获取资源-快速衰亡策略能让植物本身的净效益最大化[13],而根系分枝数在资源贫匮的土壤中减少,在资源丰富的土壤中增多的报道更多[24]。
在自我组与非我组之间,非我组虽表现出更高的根尖数、分枝数与根表面积,但其根平均直径却显著大于自我组,这意味着非我组有相对更低比例的细根,且比根长与根长密度数值低,表明非我组根系资源获取效率更低,同样地,非我组小于自我组的比叶面积(图5A)表明其叶片的捕光能力更弱[25]。自我组与非我组向两侧的根系生物量分配比较均衡,但向内侧(与邻近植株竞争方向)分配的生物量密度更大,这与连接组恰好相反,自我组与连接组的不同根系行为表明了自我身份识别的存在。对Buchloe dactyloides的研究中发现根系行为对匍匐茎保持连接与否具有时间依赖性,相比起保持连接的野牛草,放置在同一容器的2 棵分株根系生长速度更快并且随着两分株分开时间的延长,其根系生长的增幅更大,这肯定了植物的自我/非我识别以及识别过程具有时间效应[26]。此外,细根向表层土移动,两者都表现出既向公共区域竞争资源,又向外侧延展以寻觅资源来回避竞争的行为,这种竞争与基于身份识别的利他行为过程会同时发生的观点被许多学者认可[27-28]。
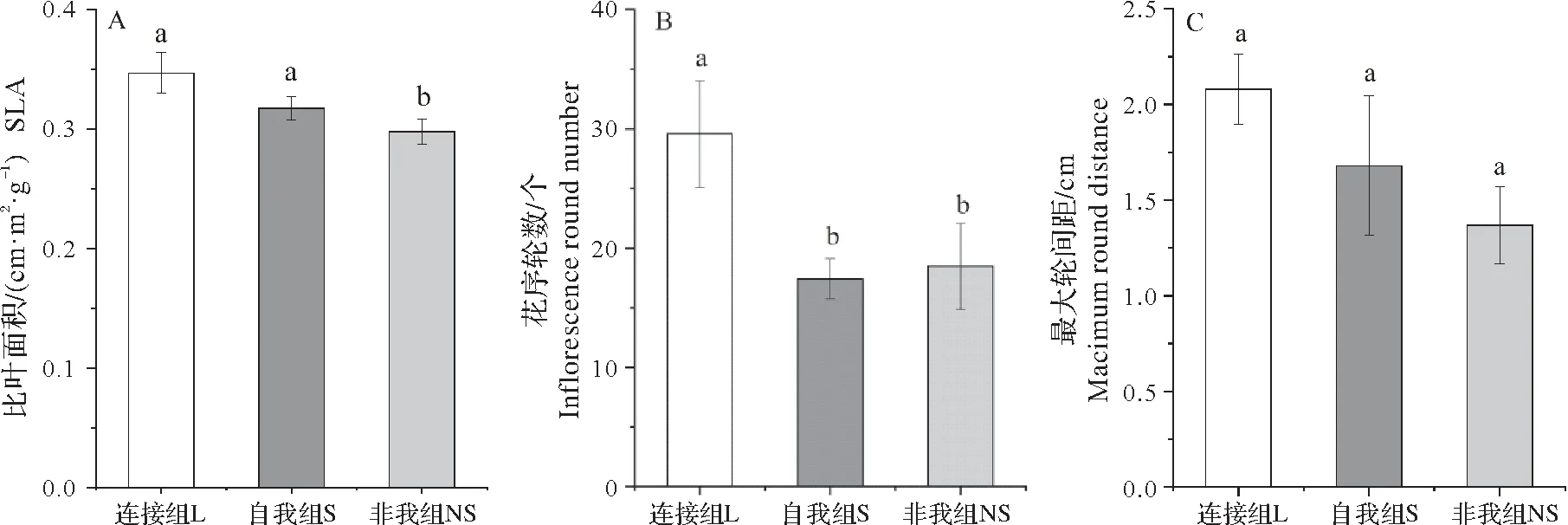
图5 不同处理组风轮菜各指标比较Fig.5 The comparation of traits in different treatments of Clinopodium chinense
通常认为将获取的养分投资到繁殖性状的生长是植物提升适合度的重要途径。研究中连接组的总体花序性状特征显著优于其他处理组,表明分株间的克隆整合作用能够有效缓解根系生长空间对植物生长的限制,这与前人研究结果一致[2,29]。自移栽后,自我组与非我组的花序生长表现出时间上的差异,邻株身份能影响植物对环境的适应能力从而体现在时间上,如早期对卡佩凤仙花的研究显示,远亲缘个体相比近亲缘个体开花时间被推迟[30],而最近对大麦(Hordeumsp.)的研究发现与同品种的大麦共种时,早熟品种的氮素积累峰值提前,晚熟品种的氮素积累峰值推迟,邻株身份影响了营养摄取的时间动态[31]。有研究表明,开花及营养摄取的过程会影响植物的种间作用响应,养分捕获的时间动态差异可以减少空间生态位的重叠,从而减少对资源的竞争[31-32],具有相似资源需求的自我组与非我组可以通过时间生态位分化来减缓激烈的竞争程度,因此,断开的风轮菜分株在花序数量上的时间差异表明其能识别遗传距离不同的分株。
综上所述,连接组的风轮菜通过匍匐茎使两分株的根系保持身份同一性,具有更高的根系活力且根系分布行为与匍匐茎断开的分株组合相异,表明风轮菜分株根系间存在一个自我识别过程。自我组相比起非我组在竞争性状上有微弱优势并在有性生殖上体现了时间差异。
