Toll样受体2与异丙肾上腺素诱导的心肌纤维化的相关性研究
2021-07-21乌日娜李猛岳强宋秀荣李晓琼徐立松
乌日娜,李猛,岳强,宋秀荣,李晓琼,徐立松
心肌纤维化指心肌正常组织结构中出现的以成纤维细胞增殖,细胞外基质(ECM)尤其是胶原纤维的过度沉积为主要表现的疾病。它是多种心脏疾病发展到一定阶段的共同病理生理改变,如缺血性心肌病、扩张型心肌病、高血压性心脏病、糖尿病性心肌病,甚至心脏移植后出现的供体心脏衰竭[1-3]。随着心脏持续的氧化应激反应和炎症损伤,心肌纤维化逐渐占据主导地位,导致心肌重构和心脏功能紊乱,从而诱发心力衰竭和心律失常,甚至心源性猝死[4,5]。目前尚无有效手段可阻止心肌纤维化的发生和发展。因此了解心肌纤维化的发生相关机制及如何有效的逆转心肌纤维化,一直是心血管领域研究的热点问题。
目前有研究显示:心肌成纤维细胞的活化与功能调节与自噬相关。自噬的主要作用是降解、清除细胞内无效的细胞成分[6]。在生理情况下,自噬在所有细胞内维持在一较低的水平,起到了维持细胞稳态的作用。在养分不足、缺血缺氧、应激状态等因素诱导下,细胞可启动自噬以产生能量,并降解受损的细胞器,从而起到自我保护的作用。但过度的自噬激活可以导致细胞死亡。心脏中基础水平的自噬对于心肌细胞的功能和稳态的维持是至关重要的[7,8]。自噬在诱导纤维化反应中的意义为临床治疗心肌纤维化和心功能不全开拓了新的领域。高迁移率族蛋白1(HMGB1)是一个高度保守的核蛋白,存在于所有真核细胞中,可在多种炎症介质的刺激下主动分泌出胞,也可在细胞坏死或受损时被动释放至胞外,参与启动非感染性炎症以及细胞凋亡和自噬[9]。Toll样受体(TRL)家族分子(包括TRL1-TRL10)可在多种细胞内表达,激活不同的配体,发挥相应的功能,TLR2和TLR4是TLR家族的两个核心成员[9],既往研究表明,HMGB1可以被免疫细胞或者心肌组织细胞所表达的模式识别受体(PRRs),如TLR2(Toll-like receptor 2)或TLR4(Toll-like receptor 4)所识别,引起心肌长时间大规模的慢性炎症反应或免疫应答[10]。目前,对HMGB1与TLR4受体相互作用的生物学功能及信号通路的研究较为广泛[11-13],然而,关于HMGB1与TLR2受体相互作用的相关研究仍鲜有报道。
本研究欲在证明TLR2能否介导细胞外的HMGB1导致自噬抑制从而引起心肌纤维化,明确TRL2、HMGB1与心肌纤维化之间的关系,这为临床治疗心肌纤维化提供了理论依据和研究线索。
1 材料和方法
1.1 实验动物SPF级C57BL/6(雄性,8周龄,平均体重20.5 g)购自北京维通利华实验动物技术有限公司。TLR2-/-(B6.129-Tlr2tm1Kir/J)小鼠购自南京大学模式动物研究所(合格证号:201503321)。饲养室恒温(22~24℃)、恒湿(55%±5%),自由饮食,每天人工光照明暗各12 h。在母鼠哺乳条件下,准确记录基因敲除小鼠出生时间,分笼(5只一笼),取8周龄的TLR2KO雄性小鼠(平均体重20.5 g),用于实验。
1.2 实验材料小鼠重组HMGB1蛋白购自美国Biolegend (San Diego,CA)。HMGB1蛋白抗体、α平滑肌肌动蛋白(α-SMA)抗体,Collagen I抗体购于英国Abcam公司。GAPDH、p-mTOR抗体、mTOR抗体购于美国Cell Signaling Technology公司。TLR2抗体购于美国R&D公司。LC3Ⅱ/Ⅰ、P62抗体购于美国Sigma公司。Alexa Fluor 488和647抗体分别来自美国Invitrogen公司。HE染色试剂盒、天狼星染色试剂盒购自上海碧云天生物技术有限公司(上海)。异丙基肾上腺素A9525购自Sigma(U.S.)。
1.3 实验方法
1.3.1 异丙肾上腺素诱导的小鼠心肌纤维化模型雄性C57BL/6和TLR2-/-小鼠(8周龄),平均体重20.4 g,ISO组于肩胛间皮下注射异丙肾上腺素(ISO),给药浓度7 mg/kg,3/d,连续15 d[14]。对照组于肩胛皮下注射相同体积的生理盐水,3/d,连续15 d。注射结束后15 d为终点(表1)。
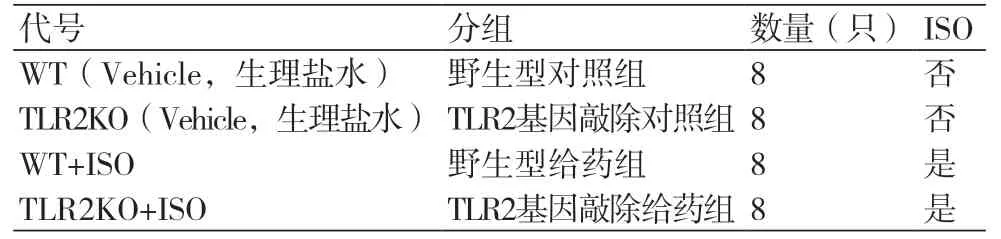
表1 实验分组情况
1.3.2 取材造模第30 d待小鼠进行超声心动图检测后取材。小鼠处死前过夜禁食,称体重。自剑突下纵行向上剪开胸腔,在大血管根部取出心脏,用冰生理盐水心脏灌注,洗尽血液,肉眼观察心脏大体形态,减除左、右心耳及残余大血管后以滤纸滤干,垂直于心脏长轴将心脏切开,一部分置于4%福尔马林溶液中固定保存,用于组织病理学检测。一部分制成冰冻切片,剩余部分存于-80 ℃超低温冰箱中保存。
1.3.3 心脏成纤维细胞的分离与培养造模成功后各组小鼠用50 mg/kg戊巴比妥钠麻醉后,仰卧位固定于鼠板上,打开腹腔,经肝门静脉注射10000 U/kg肝素使其肝素化,防止凝血。无菌条件下取出心脏后立即悬挂在Langendorf[15]装置上,首先使用KHB溶液(重碳酸盐缓冲液)灌注5 min以去除残留血细胞,再用酶溶液:0.7 mg/ml的Ⅱ型胶原酶+0.2 mg/ml的透明质酸酶+0.1%的牛血清白蛋白(BSA)+25 mmol/L的氯化钙(CaCl2)灌注10 min,有效的消化细胞外基质,随后在酶溶液中加入15 μl 100 mmol/L的CaCl2,继续灌注5 min。灌注结束后取下心房,移除心房、心包以及多余的心脏组织,将剩下的心肌组织破碎后过筛网,制成单细胞悬液,1000 rpm离心10 min后,弃上清。沉淀用8 ml含10%胎牛血清(FBS)的M199培养基重悬,并接种于细胞培养皿中。1 h后,用PBS洗去残留未贴壁的成纤维细胞,皿底为纯度较高的成纤维细胞。用含10%FBS和抗生素(青霉素以及链霉素)的M199培养基培养细胞。每7 d传代一次。
1.3.4 超声心动图检测心功能每只小鼠在处死前行经胸壁超声心动图检测。小鼠以10%水合氯醛30 ml/kg腹腔注射麻醉后,剔除颈、胸部的毛后进行超声心动图检查,该实验所有超声心动图检测均由同一技术人员进行。控制小鼠心率在500 次/min左右,在生理状态下获得测量数值。采用加拿大Visual Sonics公司Vevo 770TM型高分辨率显微超声仪,选用RMV-716探头,频率为17.5 MHz。用二维切面超声,选取受检小鼠左侧胸骨旁左室乳头肌水平短轴切面结合M型和组织多普勒超声进行检查。通过M型模式测定并计算左室射血分数(LVEF),左室短轴缩短率 (LVFS),左心室舒张末期内径(LVEDD)。所有数值均测量三个连续心动周期,取其平均值,以求尽可能减少人为原因所致的误差[16,17]。
1.3.5 形态学和组织学评价注射ISO后心肌细胞损伤及纤维化程度取小鼠新鲜心脏组织样本,4%多聚甲醛固定后石蜡包埋用于组织学检测。制成最大横截面积3 μm切片的样本,按照标准化程序用于HE染色和天狼星染色。应用高清晰彩色病理图文分析系统Spot Advanced 3.0获得高清晰的病理天狼猩红图片(400倍放大)。使用Image-Pro Plus 5.1测量每个视野天狼猩红染色后胶原着色面积。每组分析8个标本,每个标本随机选取15个视野,取均值代表一个动物胶原组织在心脏组织内的含量和表达强度。通过非参数方差分析比较各组胶原面积。
1.3.6 免疫共沉淀和免疫荧光共定位法评价HMGB1与TLR2,α-SMA与p62间的相作用细胞和组织用含1 mmol/L的苯甲基磺酰氟和蛋白酶抑制剂的放射免疫沉淀法(RIPA)裂解液处理,BCA定量试剂盒检测总蛋白含量,每孔上样40 μg进行十二烷基硫酸钠 聚丙烯酰胺凝胶(SDS-PAGE)电泳,PVDF转膜,将膜与指定的一抗孵育过夜,然后与辣根过氧化物酶标记的二抗孵育过夜,然后使用增强的化学发光检测系统(Amersham)进行信号检测(USA)。从心脏或细胞匀浆中提取蛋白裂解物,使用C0-IP裂解液,辅以1 mmol/L PMSF和蛋白酶抑制剂混合物。4℃离心机1200 rpm离心30 min,取上清液于新的离心管中。每管取50 μl作为INPUT,INPUT加入12.5 μl 5×loading buffer 98℃变性10 min。剩余裂解液每管加入20 μl Protein A/G Plus-Agarose及1:100稀释的P62抗体,混匀后,置于4℃冰箱缓慢摇动过夜。次日离心机300 rpm离心5 min,弃去上清液,每管加入500 μl Co-IP洗液洗4次。弃去上清液,加入30 μl 2×loading buffer,98℃变性10 min。变性后的INPUT和IP液8000 rpm离心 5 min即可上样,进行SDS-PAGE电泳,并采用免疫印迹检测P62及α-SMA,检测两者之间是否存在相互作用。
1.3.7 免疫荧光实验及共聚焦显微镜将冰冻心脏组织切片(5 μm厚度)或盖玻片培养的心脏成纤维细胞加入4%的多聚甲醛,室温固定10 min,加入0.2% Triton X-100,室温静置10 min。使用3%的BSA室温孵育45 min。用PBS稀释的一抗(一抗:PBS=1:100),4℃冰箱孵育过夜。PBS2次洗涤后,切片或盖玻片与用荧光素标记的经过PBS稀释的二抗(二抗:PBS=1:200),避光37℃孵育1 h。DAPI封片液室温染色40 min。通过共聚焦显微镜(USA)获得图像,并使用Leica confocal software获得分析结果。
1.3.8 电子显微镜观察心脏的超微结果和自噬小体取材即刻置入3%多聚甲醛22.5%戊二醛固定,磷酸盐缓冲液(PH7.2)冲洗; 再经1%四氧化锇室温固定120 min;双蒸水室温冲洗3次,每次2 min;50%乙醇室温10 min两次-70%乙醇室温10 min两次-90%乙醇室温10 min两次-100%乙醇25℃10 min两次,梯度乙醇脱水至环氧丙烷;环氧丙烷25℃固定10 min两次;环氧丙烷:树脂=1:1 28℃固定40 min;环氧丙烷:树脂=1:3 28℃固定40 min;纯树脂28℃固定40 min;Epon812包埋,染色后,光学显微镜定位后,制备超薄切片,醋酸双氧铀、枸橼酸铅染色,用JEM-1400(JEOL,日本)透射电子显微镜观察。
1.4 统计学分析应用SPSS 20.0统计软件,计量资料以均数±标准差的方式表达,如符合正态性分布则进行方差齐性检验,多组间比较采用单因素方差分析,组间两两比较采用LSD-t检验;如不服从正态性分布,则转化为常用对数后进行t检验。P<0.05为差异有统计学意义。
2 结果
2.1 TLR2KO改善ISO诱导的心功能不全对雄性野生型小鼠和TLR2基因敲除小鼠肩胛皮下注射异丙肾上腺素(ISO)15 d,在造模第30 d行超声心动图检查发现,注射异丙肾上腺素后野生型小鼠左室射血分数(LVEF),左室短轴缩短率(LVFS)较对照组明显减小,左心室舒张末期内径(LVIDd)较对照组显著增大。而敲除TLR2基因的小鼠在注射ISO后,左室射血分数(LVEF),左室短轴缩短率(LVFS)与对照组相比变化不大,左心室舒张末期内径(LVIDd)较对照组亦无明显变化(图1)。
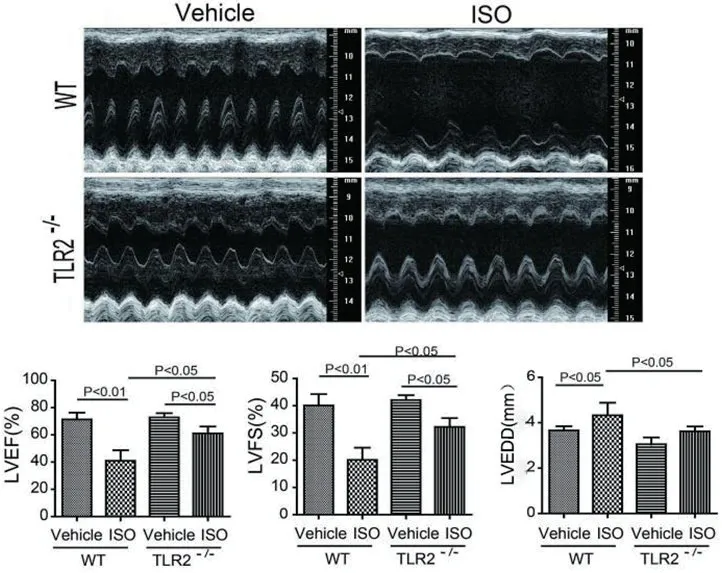
图1 各组小鼠超声心动图检测结果
2.2 TLR2KO抑制ISO诱导的心肌炎症反应和心肌细胞丢失比较HE病理结果(图2)和电镜结果(图3),可见WT+ISO组心肌损伤最重,心肌细胞的肌原纤维被破坏,线粒体发生肿胀和液泡化,灶性心肌细胞消失,细胞间隙扩大,心肌成纤维细胞和肌成纤维细胞增殖,胶原分泌成倍增加。而TLR2KO+ISO组炎症改变小,心肌损伤较轻,心肌细胞丢失少,继发纤维增生反应轻。
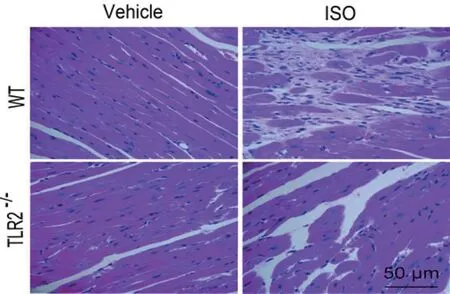
图2 各组小鼠HE染色结果
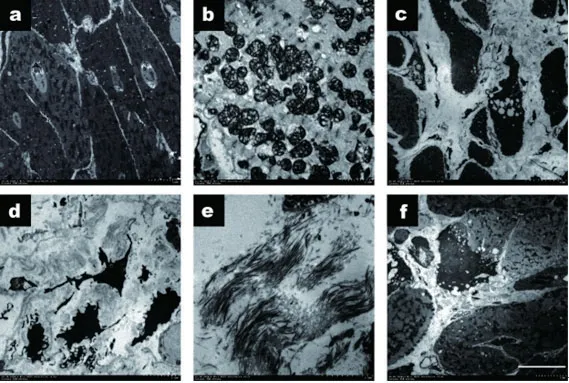
图3 电镜下的心肌超微结构改变
2.3 TLR2KO抑制ISO诱导的心肌纤维化各组小鼠天狼星染色可见,WT+ISO小鼠心肌疤痕区胶原积聚较多,而TLR2KO小鼠心肌胶原分泌明显减少,且两者胶原面积分数比有差异(P<0.01)(图4)。胶原面积分数与心肌细胞的灶性坏死面积大小相关。经免疫印迹实验证实,WT+ISO组小鼠的心肌组织中collagen Ⅰ(Ⅰ型胶原)的表达与TLR2KO+ISO组比较明显增加(图5)。
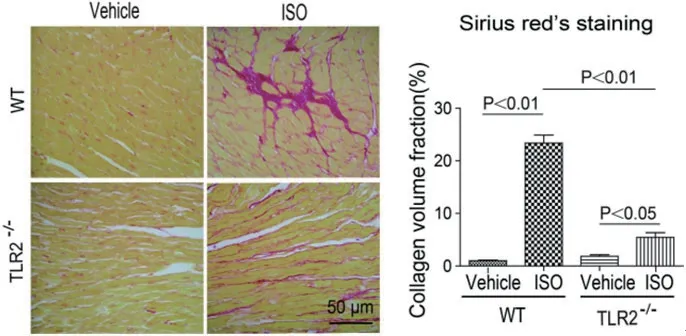
图4 各组小鼠天狼猩红染色结果
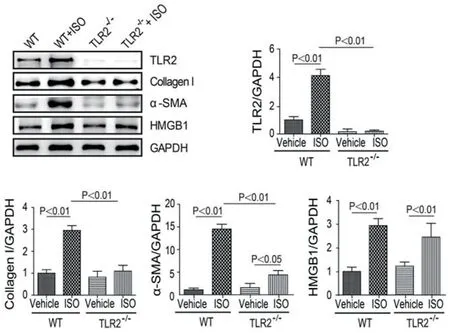
图5 各组小鼠心脏CollagenI、α-SMA、HMGB1蛋白表达情况
通过免疫印迹(图1-6)和免疫荧光检测(图6),均提示WT+ISO小鼠的心肌组织中α-SMA的含量较对照组比较明显升高,而TLR2KO+ISO小鼠心肌组织中α-SMA的含量较对照组稍有升高,但与WT+ISO比较差异有统计学意义(P<0.01)。提示WT+ISO组心肌纤维化明显,但TLR2KO+ISO组仅有轻度的纤维化。提示TLR2KO抑制ISO诱导的心肌纤维化。

图6 免疫荧光检测各组小鼠心脏α-SMA蛋白表达情况
2.4 HMGB1与TLR2的相互作用导致了心肌纤维化在免疫荧光共定位实验中(图7),发现心肌纤维化严重的WT+ISO组HMGB1含量增加,导致与TLR2相互作用增强,但因为TLR2基因被敲除,所以注射ISO的TLR2KO组仅有HMGB1表达增加,而无HMGB1与TLR2相互作用。
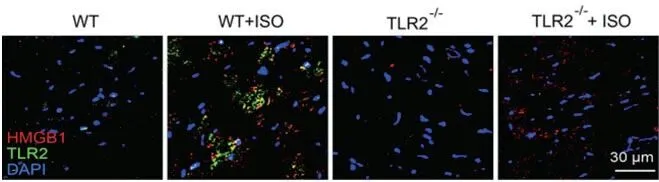
图7 免疫荧光检测HMGB1与TLR2的相互作用
2.5 ISO使小鼠心脏成纤维细胞中自噬流发生阻塞,而TLR2KO恢复了阻塞的自噬流通过免疫印迹的手段检测各组小鼠心脏成纤维细胞中自噬标志性蛋白的表达:磷酸化的mTOR,mTOR,LC3Ⅱ/Ⅰ,可溶性的p62和不可溶性的p62。同时也检测了TLR2和HMGB1的表达。与TLR2KO小鼠比较野生型小鼠在注射ISO后心脏成纤维细胞中可溶性的P62显著的增加,提示自噬是活化的;然而磷酸化的mTOR和mTOR的比值也在增加,这提示自噬受到抑制;同时心脏成纤维细胞中LC3II/I的比值显著的减小,更证实了自噬受到抑制。此外,与其他组相比,WT+ISO组不可溶性P62发生了堆积,更证实自噬流受到了阻塞。且与对照组比较,自噬流受到抑制的WT+ISO组的HMGB1及TLR2的表达均增强,而TLR2KO+ISO组仅HMGB1表达增强(图8)。说明异丙肾上腺素使小鼠心脏成纤维细胞中自噬流发生阻塞,而TLR2KO恢复了阻塞的自噬流。
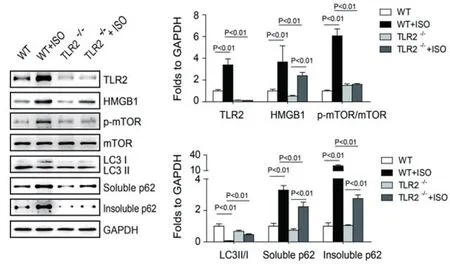
图8 各组小鼠心脏成纤维细胞中TLR2、HMGB1、p-mTOR、mTOR、LC3、可溶性P62及非可溶性P62蛋白表达情况
2.6 ISO使小鼠心肌细胞中自噬小体数目减少电镜下可观察到心肌细胞中的自噬小体,心肌细胞中的线粒体自噬,在富含肌丝中的肌成纤维细胞中也可观察到自噬小体(图9 Ac),及成纤维细胞中的自噬小体,同时发现野生型的小鼠在注射ISO后心脏组织的自噬小体数量明显少于其他组,而TLR2基因敲除小鼠在注射ISO后心脏组织中的自噬小体较野生组显著增加,且差异有统计学意义。
3 讨论
心肌纤维化指心肌正常组织结构中出现的以成纤维细胞增殖,细胞外基质(ECM)-尤其是胶原纤维的过度沉积为主要表现的疾病。它是心室重构的主要病理表现,与多种心脏疾患的病理生理改变相关,如心房颤动、缺血性心肌病、扩张型心肌病、高血压性心脏病、糖尿病性心肌病、心脏瓣膜病、心肌淀粉样变性、贫血性心脏病,甚至心脏移植后出现的供体心脏衰竭[18]。心肌纤维化发生发展的分子机制包括炎症反应,线粒体的损伤,心肌细胞的凋亡,心脏成纤维细胞的增殖,以及成纤维细胞向肌成纤维细胞表型的转换[19,20]和胶原纤维的沉积。随着心脏持续的氧化应激反应和炎症损伤,心肌纤维化逐渐占据主导地位,导致心肌重构和心脏功能的紊乱,从而诱发心衰和各种心律失常,甚至心源性猝死[4]。然而,目前没有任何有效的手段可以阻止心肌纤维化的发生和发展。因此了解心肌纤维化的发生相关机制及如何有效的逆转心肌纤维化,一直是心血管领域研究的热点问题。
研究发现细胞外HMGB1的新角色——通过与天然免疫受体TLR2的相互作用引起心肌纤维化,提示我们可通过阻断细胞外HMGB1或TLR2受体的生物学功能来治疗心肌纤维化。虽然之前的研究表明自噬是心肌纤维化的发病机制中不可缺少的防御机制[2,9],但胞外的HMGB1对自噬活动的抑制和自噬相关促纤维化蛋白的降解的阻碍却无相关报道。
在本研究中,异丙肾上腺素导致HMGB1聚集在心脏细胞膜表面,并且增强了与细胞膜表面TLR2受体的相互作用,导致了心肌纤维化。而将TLR2基因沉默后,虽然细胞间隙中HMGB1表达增强,但因无法与TLR2受体相互作用,而阻止了心肌纤维化。更重要的是,在体实验发现心肌纤维化的成纤维细胞中的自噬流发生了阻塞,且自噬小体的数量减少,而体外实验则证明细胞外的HMGB1导致成纤维细胞内自噬流的阻塞是浓度依赖型的,以及心脏成纤维细胞中I型胶原和α-SMA的也发生了浓度依赖型堆积。表明将HMGB1作为治疗靶点,为抗纤维化治疗提供了更大的潜在前景。
作为损伤相关分子模式家族(DAMPs)的重要成员,HMGB1是一个多功能的,普遍存在于细胞内和细胞外的蛋白质[21],它在各种生理和病理过程中起关键作用,包括细胞的增殖、分化、EMT、凋亡和死亡,及免疫和各种代谢过程[21,22]。HMGB1基因剔除大鼠出生后不久即死亡,说明HMGB1对动物生存不可或缺。HMGB1参与多种心血管疾病的发生发展,并在多种心血管病理组织中高表达。HMGB1在动脉粥样硬化斑块中表达升高,HMGB1可促进斑块的形成与发展[23,24]。受损的血管内皮主动或者被动释放HMGB1,促进炎症,趋化单核/巨噬细胞和血管平滑肌细胞向内皮迁移,促进斑块的形成。在斑块晚期,HMGB1可加速斑块进展导致血栓形成[25]。急性冠脉综合征患者血清HMGB1的浓度较非冠心病患者明显升高,并且与发生心血管事件的风险呈正相关[26]。在临床研究中发现,急性心梗患者血清HMGB1的水平与心血管急性事件的再发生有相关性。Giallauria等发现心梗后进行康复训练的患者较未进行康复训练的患者HMGB1水平低,他们发生心肌重构、心血管事件死亡率和全因死亡率均明显低于HMGB1水平仍高的患者[27,28]。HMGB1与心力衰竭的发生发展密切相关[29]。临床研究发现,心衰患者中HMGB1水平与其他心衰标志物如BNP、NYHA分级、射血分数等具有相关性。HMGB1水平不仅在心衰患者中表达升高,并且与心衰的严重程度相关[30]。在1型糖尿病患者中,HMGB1水平与全因死亡率、心血管事件发生率呈独立相关性[31]。在2型糖尿病患者中,血清HMGB1水平的升高与心血管事件的发生密切相关[32]。
但HMGB1在自噬与心脏纤维化过程中的作用目前还尚未看到相关报道。胞外的HMGB1需与细胞膜表面相应的膜受体结合才能发挥其生物学效应,这包括了Toll样模式识别受体(TLRs)和晚期糖基化终产物受体(RAGE)[32]。TLR家族有很多成员,目前已鉴定出TLR2、TLR4、TLR9可与胞外HMGB1结合而产生效应。其中TLR4作为LPS(内毒素脂多糖)受体被广泛熟知,TLR2则主要在革兰氏阴性细菌和真菌引起的炎症中起作用。TLR2和TLR4基因敲除小鼠证实有减少心肌梗死面积,减轻心脏炎症反应和改善心功能的作用[33,34]。本研究中,我们观察到心肌纤维化与HMGB1转移至细胞膜表面和细胞间隙有关,而沉默TLR2基因相当于阻断HMGB1与TLR2的相互作用,从而改善心肌纤维化,这表明细胞外HMGB1可能是心肌纤维化发病机制中的危险因素。
为了明确纤维化和自噬的相关性,我们进一步检测了异丙肾上腺素诱导的小鼠纤维化模型中自噬的标记性蛋白。p62是自噬途径中经典的货车蛋白,参与自噬途径中各种蛋白及分子的降解过程[35,36]。通过免疫印迹实验发现可溶性及不可溶性p62的表达均增强,提示自噬流受到阻塞,p62发生堆积,无法将待降解底物转运至自噬溶酶体。磷酸化mTOR与非磷酸化mTOR比例的显著减少和LC3Ⅱ与LC3Ⅰ比例的明显减少也提示成纤维细胞中自噬流被阻断。由于α-SMA被认为是组织纤维化的标志,我们假设阻断的自噬流导致了α-SMA的堆积是通过p62与α-SMA的相互作用来完成的。实际上,通过免疫共沉淀和免疫荧光共定位实验,发现不论在体内或者体外α-SMA和p62间都存在相互作用。说明沉默TLR2基因或阻断HMGB1后恢复了阻塞的自噬流,加速了货车蛋白p62对α-SMA的转运和降解,减轻了α-SMA的聚集,逆转了成纤维细胞的表型转换,从而改善了心脏纤维化。而α-SMA和p62之间过多的相互作用也是自噬功能障碍的重要结果。
综上所述,我们的研究结果表明异丙肾上腺素引起的心脏损伤促使HMGB1释放到细胞外。细胞外的HMGB1与TLR2受体相互作用导致心脏成纤维细胞中的自噬功能障碍使p62发生堆积,堆积的p62反过来又阻碍了α-SMA的降解,导致了心肌纤维化。因此,将HMGB1作为治疗靶点,具有重要的临床意义和药物研发价值。
