草鱼养殖模式对气单胞菌属细菌群落影响分析
2021-07-21徐先栋付辉云欧阳敏万国湲黄江峰周智勇刘礼辉
徐先栋,付辉云,欧阳敏,万国湲,饶 毅,黄江峰,周智勇,刘礼辉
( 1.农业农村部渔用药物创制重点实验室,广东省水产动物免疫技术重点实验室,广东 广州 510380; 2.江西省水产科学研究所,江西 南昌 330039; 3.江西省养蜂研究所,江西 南昌 330052 )
草鱼(Ctenopharyngodonidellus)是我国淡水水产养殖主养品种,养殖产量多年来约占据全国淡水水产养殖总产量的1/5[1],为我国水产品市场提供了价格实惠、质量优良的蛋白质来源。经过多年的养殖技术发展,目前草鱼养殖形成了以池塘精养为主,山塘养殖、鱼菜共生养殖等为补充的多种养殖模式。池塘精养和鱼菜共生模式全程投喂配合饲料,山塘养殖主要投喂草料,或少量搭配配合饲料。研究者对不同养殖模式下草鱼品质和生理功能等方面开展了研究,如张爱芳等[2]研究了此3种模式下草鱼的营养成分和氨基酸的组成,雷晓中等[3]研究了3种模式下草鱼肝胰脏结构与功能的变化。此类研究以草鱼本身为对象,研究不同养殖模式对草鱼自身指标的影响,为草鱼健康养殖提供了部分理论依据。不过,鲜见有不同养殖模式对草鱼养殖环境(水体和底泥)系统,特别是对细菌群落影响的对比研究,常见单个养殖模式对水体或底泥细菌群落的研究,如马富平[4]研究了草鱼精养模式池塘微生物群落结构,程莹寅等[5]研究了草鱼精养池塘底泥微生物群落多样性,何涛[6]研究了草鱼精养池塘水体的细菌群落结构。目前仅见其中2个模式的菌群差异的研究,如Qin等[7]研究了草鱼精养和草饲池塘细菌多态性。细菌是养殖生态系统中的重要组成部分,充当着分解者角色,关系到整个养殖环境中的物质循环和能量流动。研究草鱼养殖系统环境中的细菌群落,特别是病原菌的分布及其多样性,对于养殖病害的防治具有重要的理论和现实意义。为此笔者研究草鱼不同养殖模式下气单胞菌属细菌菌群结构与分布,为丰富草鱼健康养殖理论提供参考资料。
1 材料与方法
1.1 草鱼养殖模式
草鱼3种养殖模式分别为:草鱼池塘精养模式,产量约15 000 kg/hm2,投喂配合饲料;草鱼山塘养殖,产量约4500 kg/hm2,投喂黑麦草等草料;草鱼鱼菜共生养殖,产量约15 000 kg/hm2,投喂配合饲料,种植10%水空心菜(Ipomoeaaquatica)。采集池塘水样和底泥,采样时间为2017年5—7月(养殖投喂旺盛期),每种养殖模式选取2口池塘,在池塘两端和中部各设置1个采样点。
1.2 池塘水体细菌采集
用采水器在池塘两端和中部采集表层15 cm处水样,现场混匀后,装入灭菌塑料瓶中,低温保存,4 h内送实验室检测。采用RS培养基平板涂布法,分别取0.1 mL水样和经10倍和100倍稀释的样品涂布,28 ℃培养24 h后,取菌落数为30~100个的平板,根据菌落形态特征,挑取不同形态单菌落纯化培养,每种形态菌落至少挑取2~3个,经鉴定为同一种菌则记录为1株菌。纯化后的菌株以营养肉汤培养基培养18~24 h,加终体积分数为15%无菌甘油,-80 ℃保存备用。
1.3 底泥细菌采集
在池塘两端和中部用采泥器采集表层底泥,现场各无菌称取5 g底泥,置于装有135 mL的无菌生理盐水的塑料瓶中,振荡混匀,放置采样箱低温保存,4 h内带回实验室进行细菌分离。底泥混悬液静置后,吸取上清液0.1 mL涂布于RS培养基平板上。28 ℃培养24 h后进行细菌分离。细菌分离方法同1.2。
1.4 细菌鉴定
采用16S rDNA序列测序方法鉴定细菌。序列经16S rDNA通用引物27F/1492R PCR扩增,产物由生工生物工程(上海)股份有限公司测序。序列上传至GenBank网站比对,进行细菌鉴定。对气单胞菌属细菌,进一步结合可变区特征序列鉴定到种[8],如维氏气单胞菌(Aeromonasveronii)的16S rDNA第1可变区特征性的序列TACTGGAAACGGTA,第2可变区特征性的序列TGGTAGCTAATAACTGCCAG。数据分析采用Excel 2007软件进行。
2 结 果
2.1 细菌分离
试验共分离54株细菌。其中精养池塘分离13株,底泥分离细菌少,且菌落形态单一,经随机挑取3个菌落,鉴定同为维氏气单胞菌,记录为1株菌,水体12株;山塘养殖池塘分离20株,底泥9株,水体11株;鱼菜共生池塘分离21株,底泥8株,水体13株(表1)。

表1 养殖水体和底泥分离细菌信息
细菌经16S rDNA测序鉴定分析,共分为10属22种,其中气单胞菌属细菌占优势,为23株,其后为假单胞菌属细菌8株、柠檬酸杆菌属6株、肠杆菌属7株、泛菌属及沙雷氏菌属各3株,其他果胶杆菌属、沙门氏菌属、克吕沃尔菌属、勒克菌属各1株(图1)。按种类分,维氏气单胞菌最多,为12株,嗜水气单胞菌次之,为10株。烟粉肠杆菌6株,弗氏柠檬酸杆菌5株,其他菌1~2株不等(表1,图1)。

图1 分离细菌种属分布Fig.1 The genera of isolated bacteria
2.2 细菌种类分析
按养殖模式来源分:鱼菜共生养殖池塘分离细菌种类最多,为13种21株菌,分离的嗜水气单胞菌和维氏气单胞菌占比较少,分别为3株和4株;假单胞菌属细菌4株;山塘养殖环境中共分离到12种20株菌,其中嗜水气单胞菌占多数,为7株,维氏气单胞菌3株次之;精养池塘中分离细菌种类最少,只分离到5种菌,13株,其中维氏气单胞菌和柠檬酸杆菌属占优势,各为5株(图2)。
按细菌环境来源分:底泥中共分离15株菌,分属5种,以气单胞菌属和柠檬酸杆菌属细菌为主,其中嗜水气单胞菌占绝大多数,为9株,其他细菌如弗氏柠檬酸菌为4株,豚鼠气单胞菌、维氏气单胞菌以及烟粉肠杆菌各1株。水体中共分离39株菌,分属21种,优势种属为气单胞菌属、假单胞菌属及肠杆菌属细菌,分别为12株和5株,其中维氏气单胞菌分布最多,为11株,其他种属细菌,如泛菌属、果胶杆菌属细菌等1~2株不等(图3)。
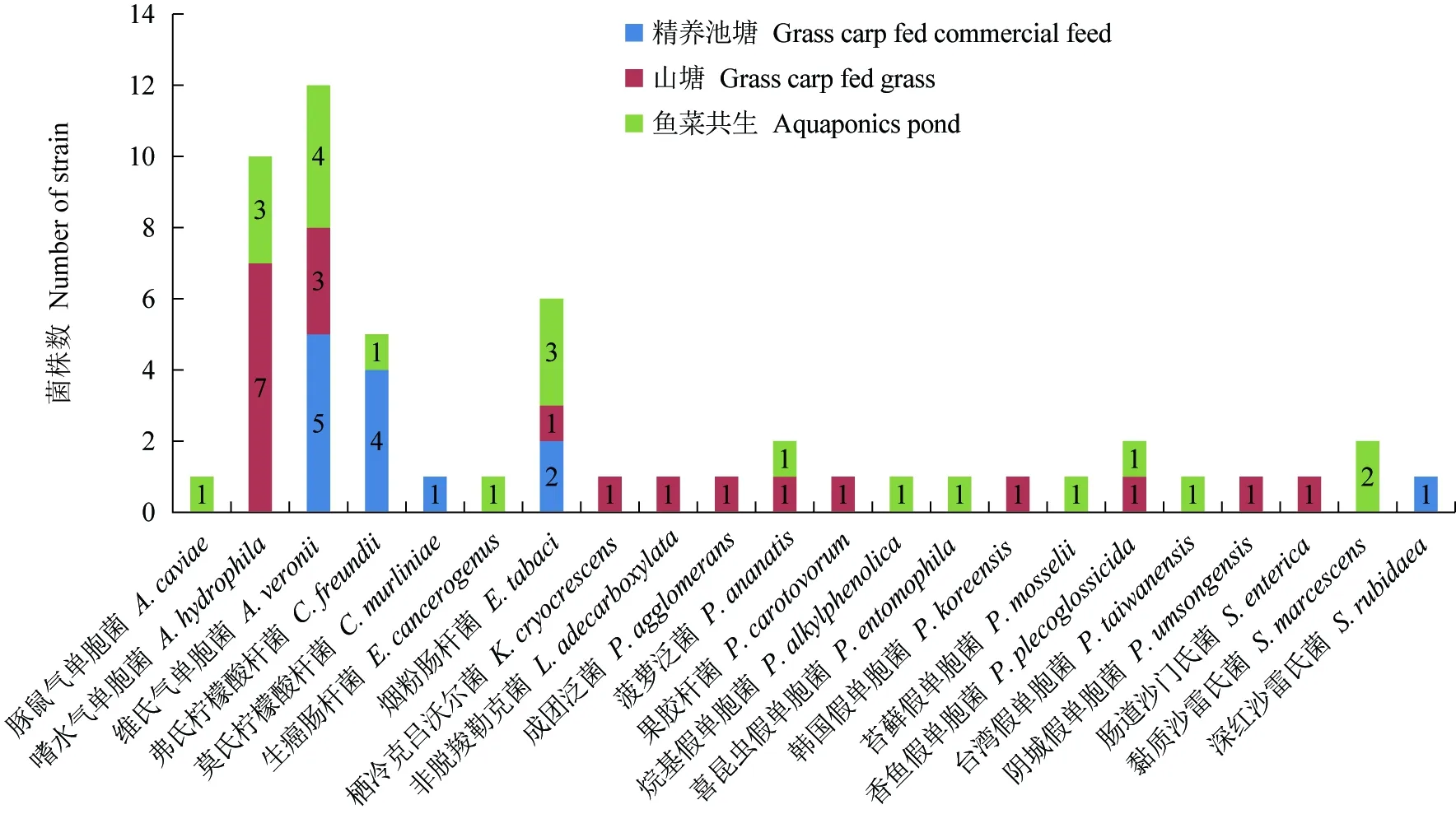
图2 不同养殖模式分离细菌统计Fig.2 The statistics of bacteria isolated from different culture modes

图3 底泥和水体分离细菌种类分布Fig.3 The bacterial distribution in sediment and water
3 讨 论
3.1 RS培养基平板分离细菌分析
RS培养基是20世纪70年代初期研制的最早是用于嗜水气单胞菌的选择性培养基,根据嗜水气单胞菌发酵麦芽糖产生黄色菌落而鉴别[9]。气单胞菌属细菌是淡水养殖主要病原菌,本试验主要是研究草鱼不同养殖模式环境中的气单胞菌属细菌分布情况,为草鱼在不同养殖模式下健康养殖提供理论参考,故采用RS培养基进行细菌的分离。在RS培养基上,根据细菌的不同生化特性,还会产生3种不同菌落形态,如黄色菌落中心现黑点,该类细菌具有发酵麦芽糖并产生H2S,包括变形杆菌属(Proteus)、柠檬酸杆菌属细菌等;黄绿色菌落,显示赖氨酸或(和)鸟氨酸脱羧,包括沙门氏菌属、假单胞菌属、肠杆菌属细菌等;绿色菌落中心现黑色表示的是赖氨酸或(和)鸟氨酸脱羧,并且产生H2S,该类细菌有柠檬酸杆菌属、沙门氏菌属、爱德华氏菌属(Edwardsiella)等。20世纪70年代初期,气单胞菌属中许多细菌还未有分离报道,现在随着细菌分离和鉴定技术的发展,发现许多气单胞菌属细菌表现不同的生理生化特性,如维氏气单胞菌、温和气单胞菌(A.sobria)具有赖氨酸脱羧特征,杀鲑气单胞菌(A.salmonida)产生H2S等[10],从而使RS培养基在鉴定气单胞菌属细菌显示了多种形态。为研究细菌分布多态性,将可以在RS培养基上生长的细菌进一步分离纯化鉴定,从而使鉴定的种类出现了假单胞菌属、柠檬酸杆菌属细菌等。
3.2 不同养殖模式下细菌分布情况分析
从不同养殖模式细菌种类上可以看出,鱼菜共生养殖环境中细菌分离种类最多,暗示细菌多态性可能也最高,细菌多态性高的池塘,水质更好,最不易发病[11-12]。如孙苏燕等[13]研究了健康和患病三疣梭子蟹(Portunustrituberculatus)养殖池塘表层底泥细菌群落的变化,结果表明,病害塘中变形菌门细菌无论是多样性还是丰度均有所降低。山塘养殖池塘分离细菌种类也较多,不过因其产量低,效益不高,所以代表性不强。与已有的研究结果[14]相似,本试验中精养池塘细菌种类较少,潜在致病菌如维氏气单胞菌、嗜水气单胞菌等含量多。分析原因可能是[14]:一方面沉积在底层的残饵、粪便等有机物超出了环境自净能力,阻碍了营养物质向微生物输入,未被充分氧化分解的残饵、粪便产生有害物质,也阻碍了微生物生长;另一方面,随着配合饲料等的持续性加入,对菌群进行了定向性驯化,从而导致了菌群种类的降低。因而,精养池塘养殖鱼类更易发生病害,鱼肉品质也稍有下降[2]。相比而言,鱼菜共生系统,综合了水产养殖和水栽培技术,在净化水质和促进物质转移方面具有显著作用,可产出优质水产品和新鲜蔬菜,获得双重效益,值得大力推广。
3.3 水体和底泥细菌分布特征
试验分离的气单胞菌属细菌,如维氏气单胞菌和嗜水气单胞菌分布具有明显的环境差异。水体中以维氏气单胞菌为主,12株维氏气单胞菌中11株分离自水体,占比为91.6%;底泥中以嗜水气单胞菌为主,本试验10株嗜水气单胞菌中有9株分离自底泥,占总数的90%。结果与已有的研究相似,如杨五名[15]研究淡水池塘中气单胞菌的生态学与流行病学,结果也显示,嗜水气单胞菌在淡水养殖水体中的分布并不广泛,丰度也不高。维氏气单胞菌是在水体中分布最广泛、丰度也较高的气单胞菌种类。诸多研究均表明,维氏气单胞菌是近年来淡水养殖主要病原菌[16-18]。维氏气单胞菌是条件致病菌,在水体中分布广泛,与鱼体直接接触,当鱼体因捕捞生产操作受伤,或环境变化,引起的免疫力低下等因素,易患维氏气单胞菌病,可能正是近年来该病流行的原因。已有研究表明,底泥是鱼类细菌病的病原库[5],本试验中分离的嗜水气单胞菌、柠檬酸杆菌属细菌[19]等致病菌大都存在于精养池塘的底泥中,也证明了这一点。致病菌和环境中其他微生物处于一种动态平衡之中,彼此竞争,如果养殖环境恶化时,微生物群落结构也会发生变化,许多条件致病菌就会大量繁殖,养殖动物就很容易患病。研究结果提示,要做好维氏气单胞菌的防控工作,同时要维护好整个养殖系统环境稳定,使嗜水气单胞菌不能在底泥中释放,以降低草鱼疾病发生风险。
4 结 论
草鱼池塘精养殖、山塘草饲养殖和鱼菜共生养殖3种养殖模式中底泥和水体的气单胞菌属细菌分布研究表明:鱼菜共生养殖模式,水体和底泥中细菌种类多,潜在致病菌数量少,与前两种模式相比,草鱼鱼菜共生养殖模式在降低疾病暴发风险和提高养殖综合效益上具有优势。
