转抗虫基因107杨对主要节肢动物种群的影响
2021-07-20任敏霞张子恒曾月霞李含笑王利峰李会平刘军侠
任敏霞,张子恒,曾月霞,李含笑,王利峰,李会平,2,刘军侠,2
(1河北农业大学 林学院,河北 保定 071000;2河北省林木种质资源与森林保护重点实验室,河北 保定 071000)
病虫害作为限制农林业发展、造成经济损失的重要因素,已无法通过传统方法获得有效抑制[1],因此利用转基因技术将特定基因转入植物体使其获得抗性来抑制病虫害的发生成为当前研究的热点[2]。杨树作为我国造林绿化重要树种,具有基因组小、离体操作容易的特点,常作为转基因林木模式树种进行研究。目前研究处于前列的为转(Bt)抗虫基因杨,自1993年将对鳞翅目昆虫具有毒性的Bt基因成功转入欧洲黑杨以来,已成功培育出多种抗虫基因杨[3],一部分已进入田间试验与商业化种植阶段,具有良好抗虫效果。
随着研究的深入,转抗虫基因杨树生态安全性问题引起众多学者广泛关注[4]。生态系统中,既包括害虫又包括天敌,害虫分为主要害虫与次要害虫,而天敌在抑制次要害虫数量以及调节害虫种群数量之中有非常大的作用[5,6]。因此近年来相关研究人员做了一些关于转抗虫基因杨对主要节肢动物种群影响的研究。目前的研究表明:转不同种类型的抗虫基因杨对靶标害虫均表现较高抗虫性,食叶害虫数量明显减少[4,7,8],且转基因杨树抗虫性会影响到下一代幼虫,使其死亡率提高,表明转基因杨树抗虫具有持续性[4,9,10]。对靶标害虫杨扇舟蛾种群生命表的研究中也证明转基因杨树杨扇舟蛾存活率与对照林地相比虫口密度显著降低、生长发育迟缓,表明转基因杨可以减少靶标害虫自然种群数量,甚至影响未来种群的发展[11-12]。转基因杨在有效抑制靶标害虫的同时,对非靶标害虫及天敌昆虫具有不同的效应[7]。一部分研究表明转基因杨在抑制靶标害虫的同时,对于非靶标害虫没有毒杀作用,且寄生于靶标害虫的天敌种类、数量和寄生率都高于对照林地[12],转基因杨使天敌种群数量增加。而其他研究表明非靶标害虫种类、数量具有增多趋势,转基因杨对主要捕食性天敌数量无影响,而寄生性天敌的数量则明显减少[13-15]。
目前转基因杨对主要节肢动物种群影响的研究尤其是对非靶标生物的影响结论不尽相同,需对其继续进行探究,可为其生态安全性提供科学依据并为以后更好地防治提供方法[16]。因此以转基因107杨为研究对象,通过研究转基因107杨对主要节肢动物种群数量及动态的影响,探究其生态安全性为进一步推广与应用提供理论依据。
1 材料与方法
1.1 试验材料
本试验林分为转抗虫基因107杨(BtCry1Ac和API基因),筛选出对美国白蛾、杨扇舟蛾等具有高抗虫的#1、#2、#3、#9株系[17]。
1.2 试验林概况
试验林地位于河北省滦南县国有林地苗圃场,属暖温带半湿润季风性气候,年平均气温10.6 ℃,常年降水量658 mm。试验林采用随机区组设计的方法,划分为4个区组,每个区组种植有#1、#2、#3、#9株系与对照107杨,每个株系为1个小区,每个小区种植30株树,株距2 m,行间距5 m。试验林地貌气候一致,管理方式一致,未施用任何化学农药。
1.3 调查方法
2019年5-10月,每20 d调查1次,共调查8次。采取随机选取样方的方法,每个区组随机选取5株样树,树冠:每株样树(东、西、南、北)4个方位上下各选取50 cm样枝,观察并记录其上主要节肢动物种类和数量;主干:调查距地表3 m内主干主要节肢动物种类与数量并记录;地面:选取的样地内5个1 m×1 m的样方进行调查,记录主要节肢动物种类与数量。对无法识别的昆虫放入装有酒精的指形管中,带回实验室并借助工具进行鉴定[18-19]。
1.4 数据处理
利用SPSS 21.0中单因素(One-Way ANOVA)方差分析对主要节肢动物种群数量进行显著差异性分析(P<0.05),相关数据整理与绘图在Microsoft Excel 2010中完成。
2 结果与分析
2.1 转抗虫基因107杨节肢动物群落组成
转抗虫基因107杨节肢动物群落组成见表1。
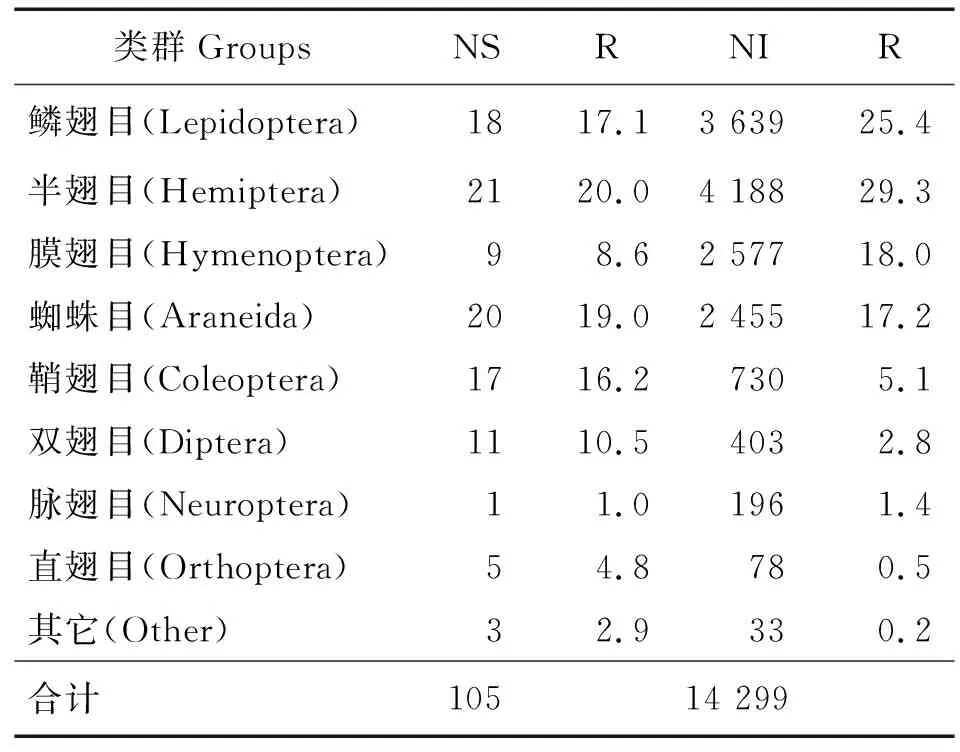
表1 转抗虫基因107杨节肢动物群落组成Table 1 Composition of arthropod community of transgenic insect-resistant gene 107 poplar
由表1可见,共获得节肢动物14 299头,分属11个目,105种。物种数以鳞翅目、半翅目、蜘蛛目、鞘翅目、双翅目为优势类群,占比为17.1%、20.0%、19.0%、16.2%。个体数量半翅目占优势地位,占比为29.3%,其次为鳞翅目、膜翅目与蜘蛛目,占比分别为25.4%、18.0%、17.2%。半翅目个体数量较多可能是由于转基因107杨对鳞翅目靶标害虫的控制作用,靶标害虫数量减少,导致半翅目个体数量增多。
2.2 主要靶标害虫个体数量及动态变化
2.2.1 主要靶标害虫个体数量 主要靶标害虫个体数量方差分析见表2。
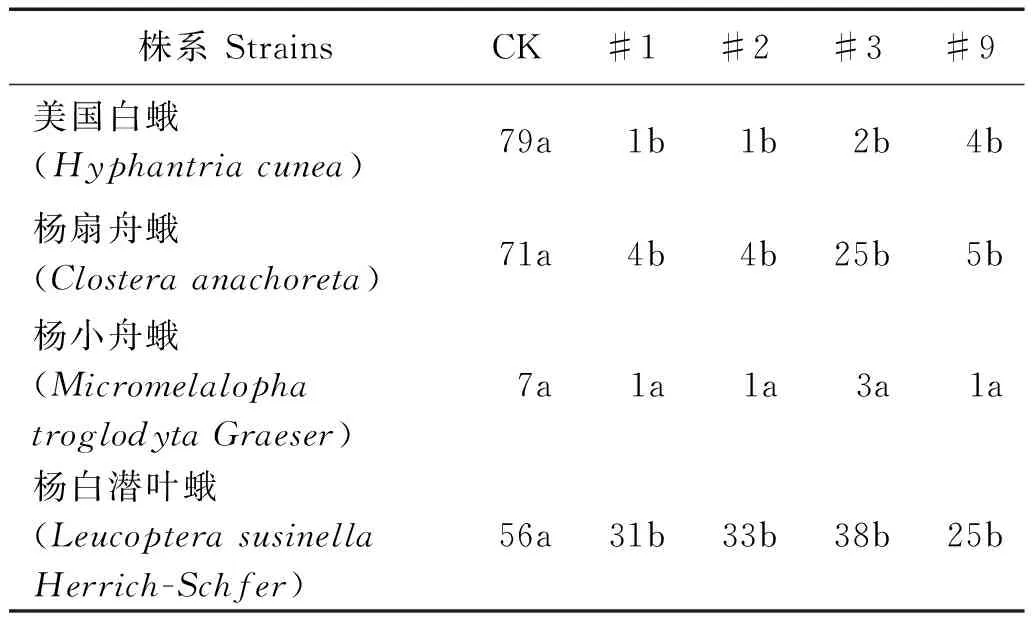
表2 主要靶标害虫个体数量方差分析Table 2 Variance analysis of main target pest in individual number
由表2可见,转基因107美国白蛾、杨扇舟蛾与杨白潜叶蛾个体数量显著低于对照组(P<0.05),杨小舟蛾个体数量相对较少,与对照组差异不显著(P>0.05),各株系间主要靶标害虫个体数量不存在显著性差异,#2株系害虫数量相对较少。上述结果表明转基因107杨对主要靶标害虫具显著抗虫效果。
2.2.2 主要靶标害虫个体数量动态变化 美国白蛾、杨扇舟蛾、杨小舟蛾及杨白潜叶蛾个体数量动态变化见图1-图4。
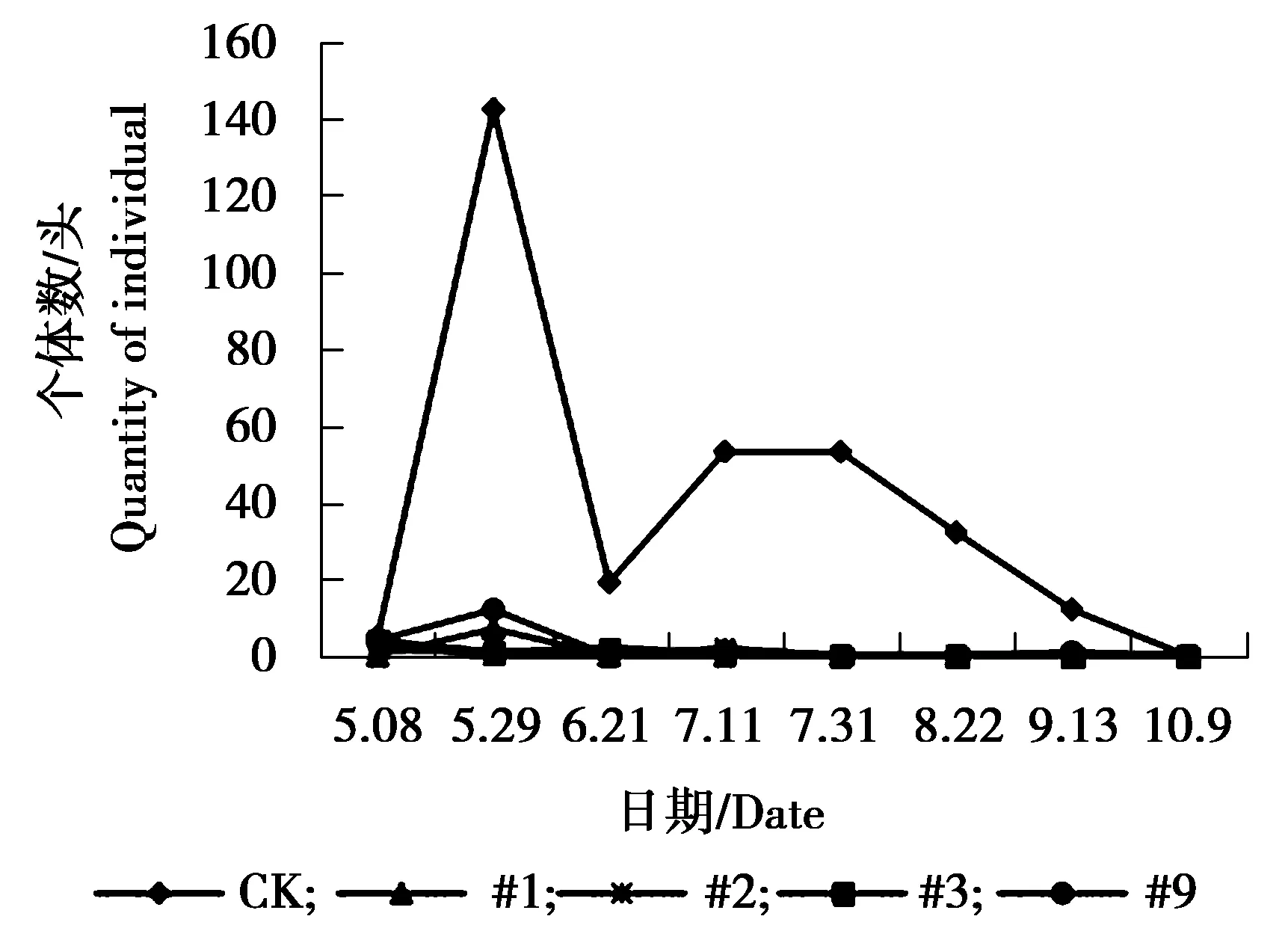
图1 美国白蛾个体数量动态变化Figure 1 The individual number dynamics of Hyphantria cunea
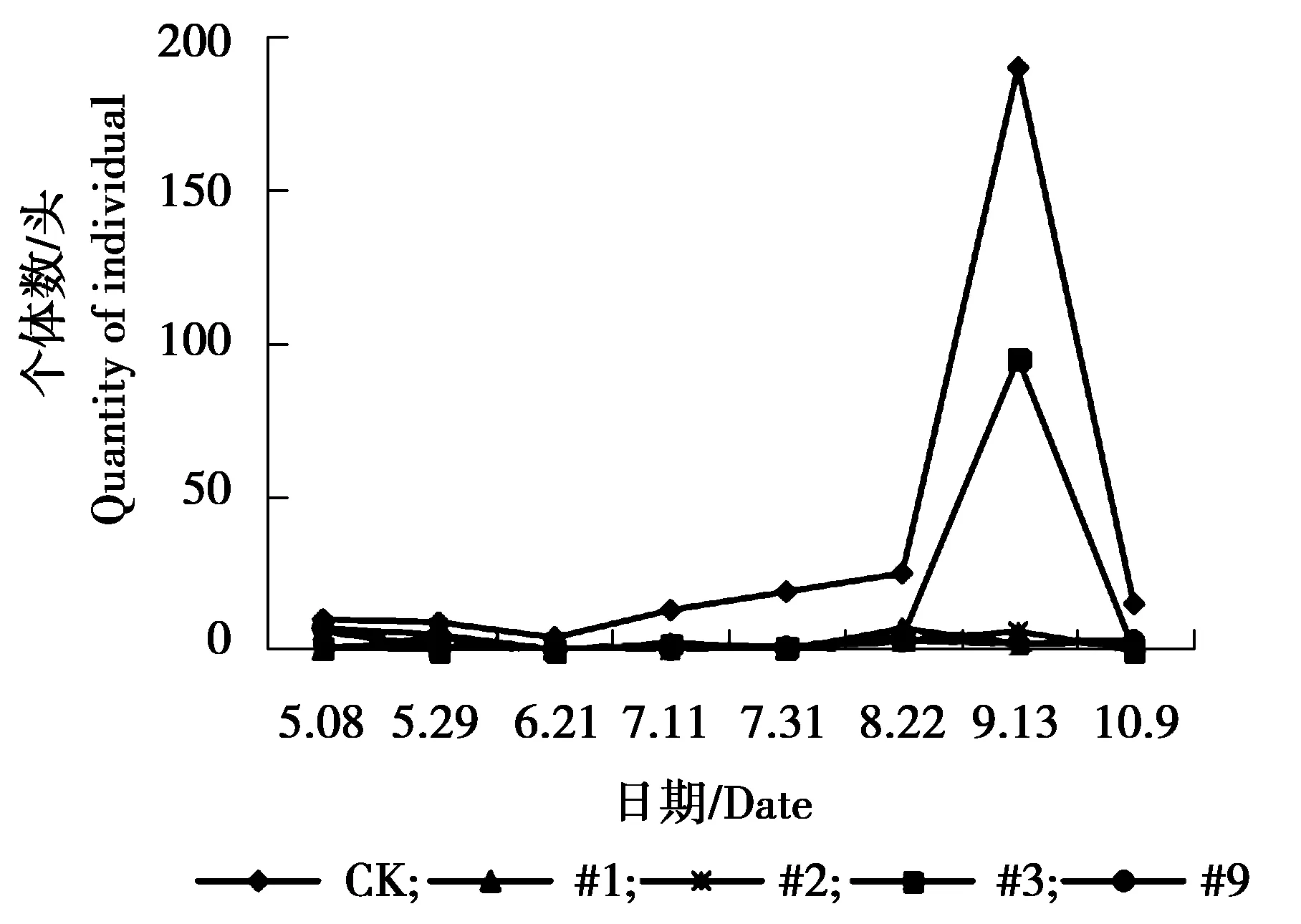
图2 杨扇舟蛾个体数量动态变化Figure 2 The individual number dynamics of Clostera anachoreta
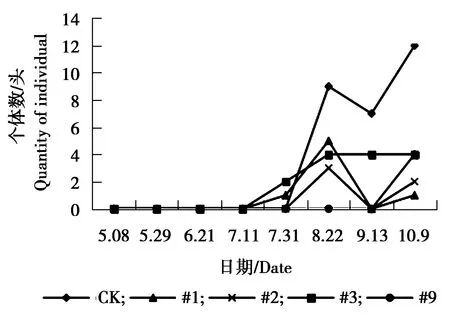
图3 杨小舟蛾个体数量动态变化Figure 3 The individual number dynamics ofMicromelalopha troglodyta Graeser
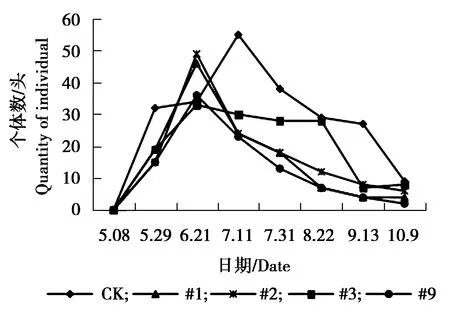
图4 杨白潜叶蛾个体数量动态变化Figure 4 The individual number dynamics of Leucopterasusinella Herrich-Schfer
由图1可见,各株系美国白蛾个体数量相对低于对照组,对照组美国白蛾个体数量在5月8日-5月29日为个体数量急剧增加的时期,5月29日达到发生最高峰,之后急剧下降,又上升,最终处于平稳状态。#1、#2、#3、#9株系美国白蛾个体数量相差不大,处于相对稳定偏低状态。由图2可见,对照组杨扇舟蛾个体数量相对高于各株系,8月22日-9月31日为其个体数量发生高峰期,#3株系杨扇舟蛾个体数量同样在8月22日-9月31日为发生高峰期,个体数量相对较多。其他各株系杨扇舟蛾个体数量较少一直处于相对稳定状态。由图3可见,对照组杨小舟蛾个体数量相对高于各株系,对照组杨小舟蛾在8月上旬为其发生高峰期,8月下旬至9月初个体数量减少,9月下旬至10月初个体数量又增多,之后个体数量减少趋于平稳。#2、#3、#9株系在7月31日个体数量开始增多,8月22日达到发生最高峰,之后逐渐减少趋于平稳,#1株系杨小舟蛾个体数量相对较少,一直处于较低水平。由图4可见,整体上各株系杨白潜叶蛾个体数量低于对照。对照组杨白潜叶蛾个体数量在6月21日-7月11日相对较多,7月11日达到发生最高峰。各株系杨白潜叶蛾个体数量在5月29日-6月21日为发生高峰期,之后呈逐渐下降趋势。
2.3 主要非靶标害虫个体数量及动态变化
2.3.1 主要非靶标害虫个体数量 主要非靶标害虫个体数量方差分析见表3。
由表3可见,主要非靶标害虫叶蝉sp.、皱大球蚧及大青叶蝉个体数量与对照组差异不显著(P>0.05),白杨毛蚜个体数量显著低于对照组(P<0.05),各株系间叶蝉个体数量存在一定差异,#3个体数量显著低于#1、#9,各株系间白杨毛蚜、皱大球蚧及大青叶蝉个体数量差异不显著。表明转基因107杨对主要的非靶标害虫未产生显著不利影响。
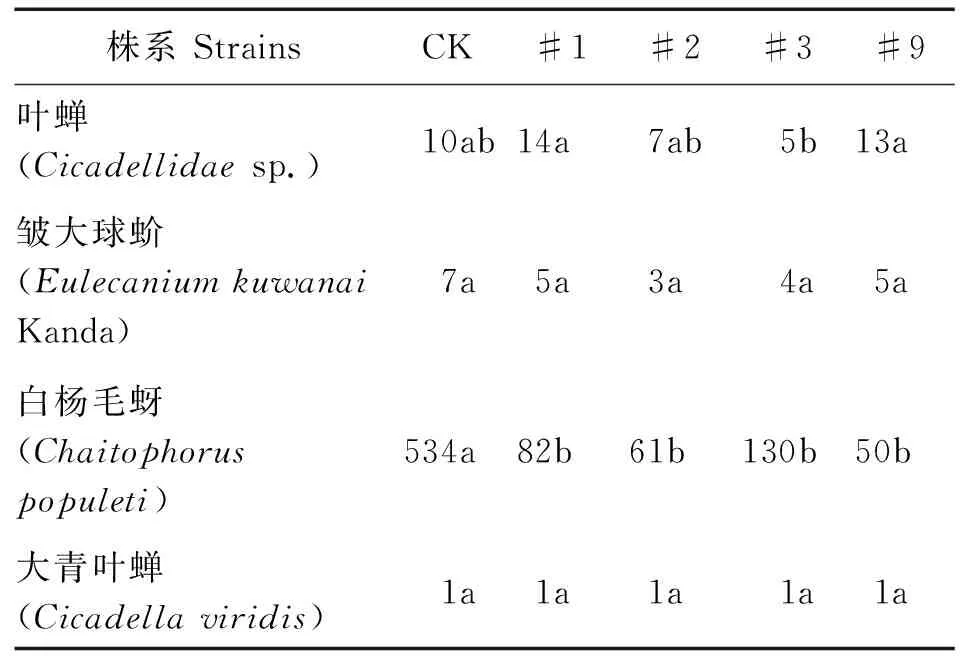
表3 主要非靶标害虫个体数量方差分析Table 3 Variance analysis of main non-target pest in individual number
2.3.2 主要非靶标害虫个体数量动态变化 叶蝉sp.、皱大球蚧、白杨毛蚜及大青叶蝉个体数量动态变化见图5-图8。
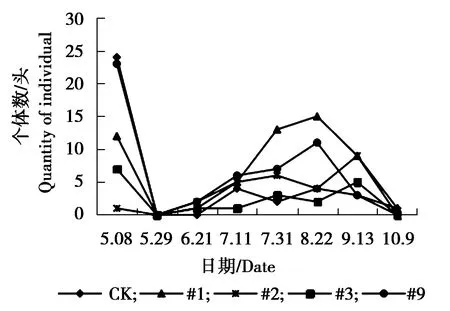
图5 叶蝉sp.个体数量动态变化Figure 5 The individual number dynamics ofCicadellidae sp.
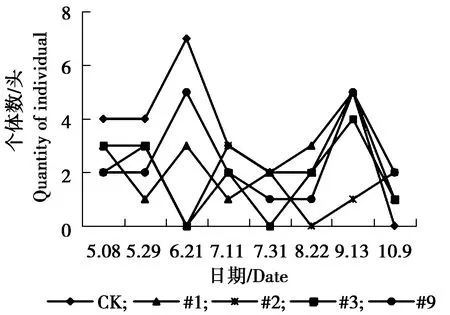
图6 皱大球蚧个体数量动态变化Figure 6 The individual number dynamics ofEulecanium kuwanai Kanda
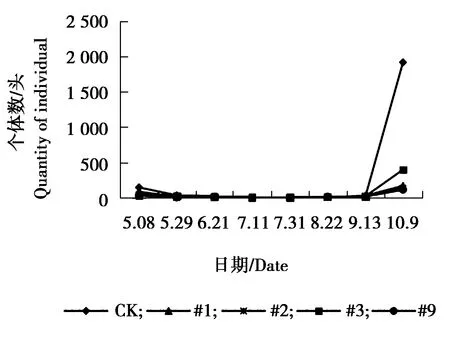
图7 白杨毛蚜个体数量动态变化Figure 7 The individual number dynamics ofChaitophorus populeti
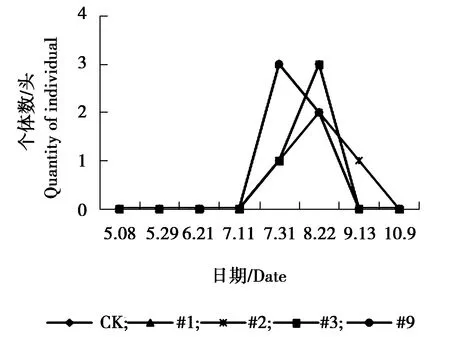
图8 大青叶蝉个体数量动态变化
由图5可见,#9株系与对照组叶蝉sp.个体数量5月29日为发生最高峰,5月29日之后,对照与各株系叶蝉sp.个体数量均略有增加,相对平稳。由图6可见,#1株系、#9株系与对照皱大球蚧个体数量在6月21日为发生最高峰,9月13日为#1、#9株系与对照个体数量发生小高峰期。#2株系、#3株系个体数量相对较少,一直处于较低水平状态。由图7可见,各株系白杨毛蚜个体数量低于对照组,各株系及对照组白杨毛蚜个体数量表现为相同变化趋势,在5月8日-9月13日个体数量一直较少,10月9日以后白杨毛蚜个体数量急剧增加,达到发生最高峰。由图8可见,对照、#3大青叶蝉个体数量在7月11日以后逐渐增加,8月上旬达到发生最高峰,之后数量逐渐减少,#9株系在7月11日为发生最高峰,之后个体数量下降,8月上旬个体数量又增多,为大青叶蝉个体数量发生小高峰期,#1、#2个体数量较少,变化幅度较小,相对稳定。
2.4 主要天敌个体数量及动态变化
2.4.1 主要天敌个体数量 主要天敌个体数量方差分析见表4。
由表4可见,转基因107杨主要天敌异色瓢虫、狼蛛sp.、圆蛛sp.、跳蛛sp.个体数量与对照组无显著性差异(P>0.05),部分株系主要天敌个体数量表现为一定程度的增多,表明转基因107杨对主要天敌未造成不利影响。
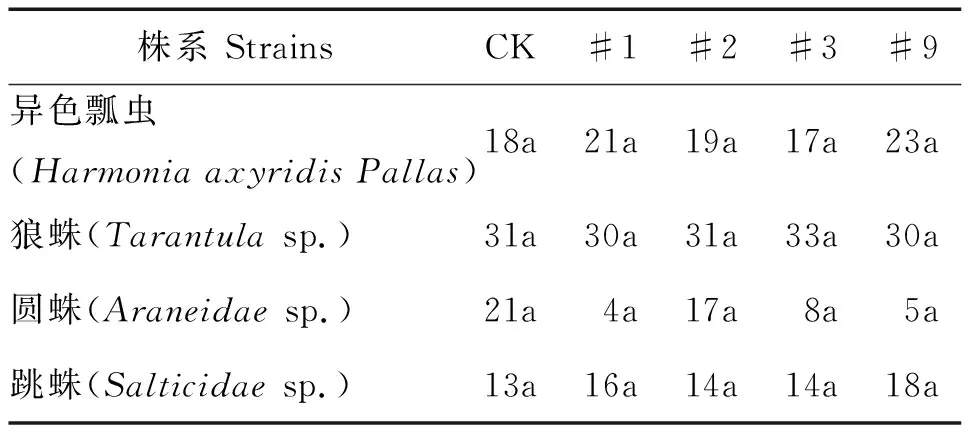
表4 主要天敌个体数量方差分析Table 4 Variance analysis of main natural enemiesin individual number
2.4.2 主要天敌个体数量动态变化 异色瓢虫、狼蛛sp.、圆蛛sp.及跳蛛sp.个体数量动态变化见图9-图12。
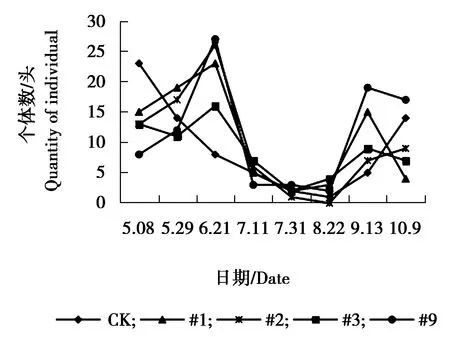
图9 异色瓢虫个体数量动态变化
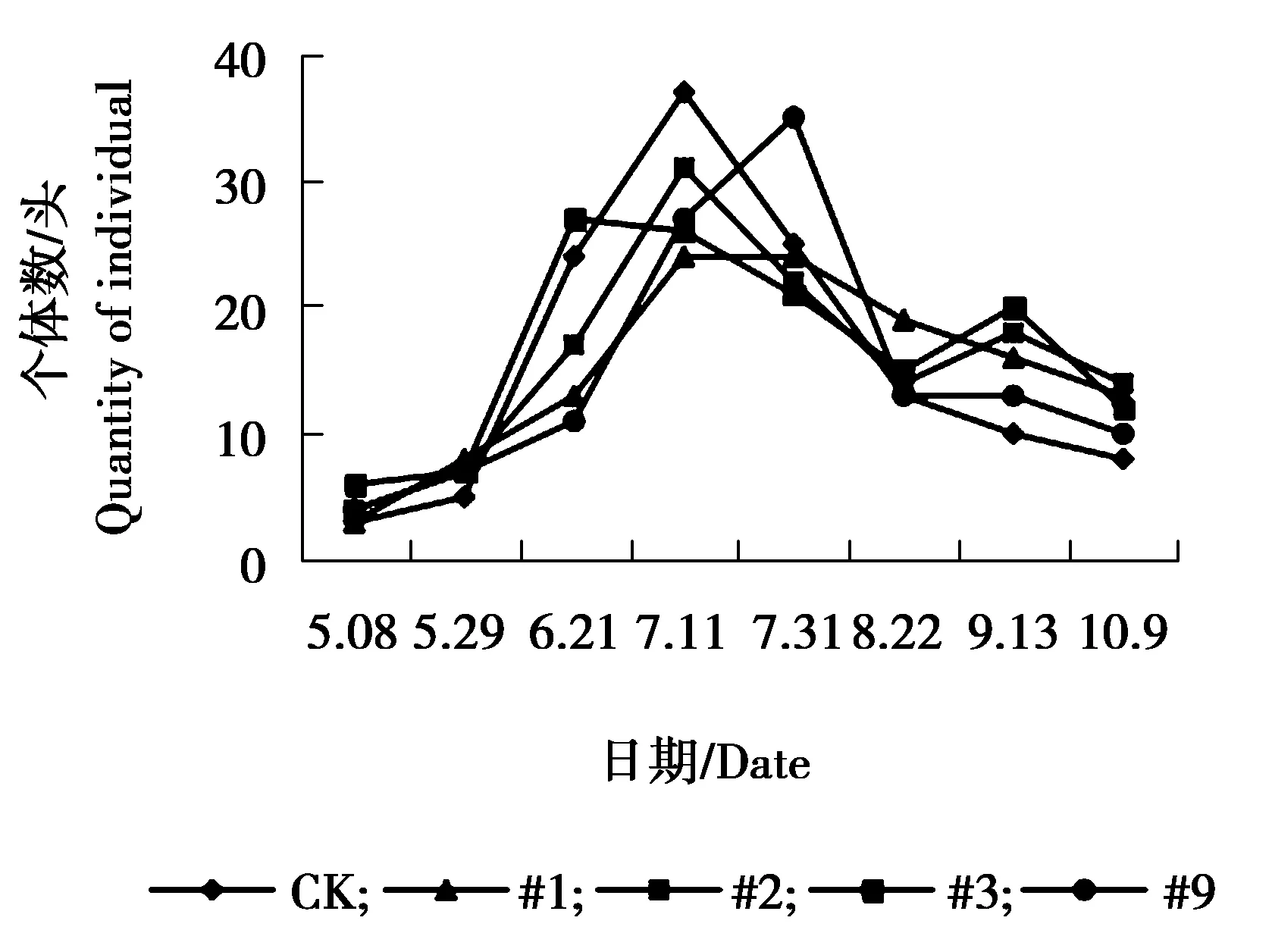
图10 狼蛛sp.个体数量动态变化
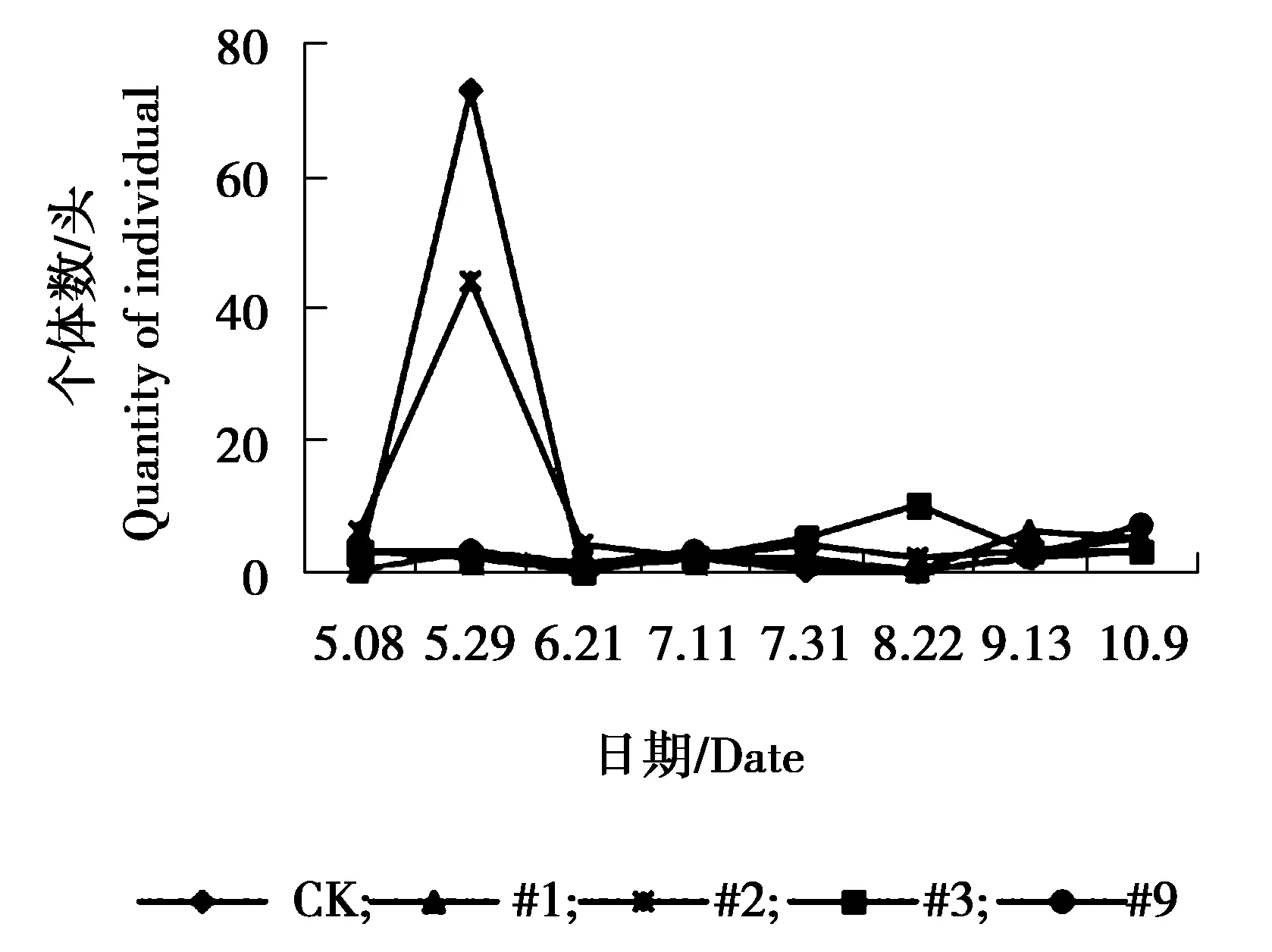
图11 圆蛛sp.个体数量动态变化
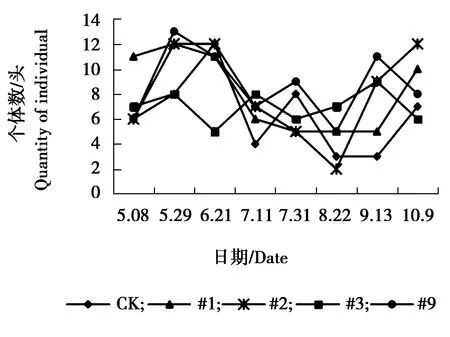
图12 跳蛛sp.个体数量动态变化
由图9可见,异色瓢虫各株系与对照组个体数量相差不大,#1、#3、#9株系与对照组个体数量在6月21日达到发生最高峰,9月13日为发生小高峰,#2株系在5月29日与9月13日个体数量相对较多,为发生高峰期。由图10可见,各株系与对照狼蛛sp.个体数量相差不大,对照组、#1、#2、#3狼蛛sp.个体数量在7月11日达到发生最高峰,#9株系7月31日达到发生最高峰,7月31日之后,各株系与对照个体数量呈逐渐下降趋势。由图11可见,5月8日-5月29日对照组与#2株系圆蛛sp.个体数量相对较多达到峰值,之后又逐渐下降,趋于平稳。其余各株系圆蛛sp.个体数量增幅不大,总体呈偏低、均衡的状态。由图12可见,各株系与对照组跳蛛sp.个体数量变化幅度较大,整体呈现为上升、下降、上升再下降的趋势。5-6月份为各株系及对照个体数量发生高峰期,7、8月份跳蛛sp.个体数量下降,相对较少,之后个体数量又表现为上升趋势,不同月份间跳蛛个体数量波动较大。
2.5 主要中性昆虫个体数量及动态变化
2.5.1 主要中性昆虫个体数量 主要中性昆虫个体数量见表5。

表5 主要中性昆虫个体数量方差分析Table 5 Variance analysis of main neutral enemiesin individual number
由表5可见,转基因107杨家蝇、家蚁个体数量与对照组无显著性差异(P>0.05),各株系家蝇个体数量相对于对照组表现为一定程度的增多,表明转基因107杨未对主要中性昆虫产生显著不利影响。
2.5.2 主要中性昆虫个体数量动态变化 家蝇、家蚁个体数量动态变化如图13、图14所示。
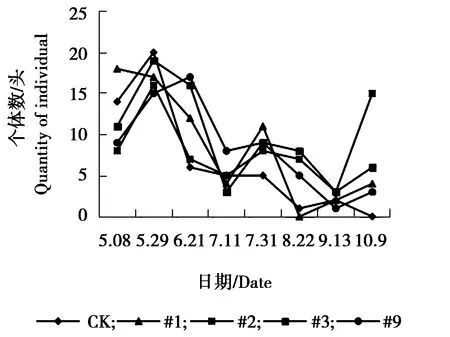
图13 家蝇个体数量动态变化Figure 13 The individual number dynamicsOf Musca domestica
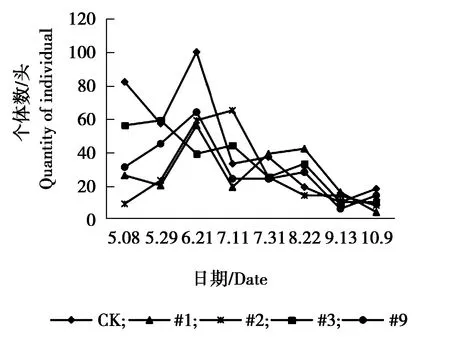
图14 家蚁个体数量动态变化Figure 14 The individual number dynamicsOf Monamorium pharaonis
由图13可见,各株系与对照组家蝇个体数量变化趋势相同,5月8日-5月29日个体数量较多,保持相对较高水平,为发生高峰期,其余月份各株系与对照个体数量随月份的变化幅度较大,整体呈现为下降、升高再下降的趋势。由图14可见,对照组与各株系间家蚁个体数量相差不大,但总体呈现的动态趋势不稳定,CK、#3、#9在5月29日-6月21日为发生高峰期,6月21日达到最高峰,#1株系在7月11日-7月31日为发生高峰期,在7月31日达到个体数量发生最高峰,不同月份之间家蚁个体数量波动较大。
3 结论与讨论
通过对转基因107杨主要节肢动物种群种类、数量及动态进行调查研究,得出结论如下:转基因107杨各株系美国白蛾、杨扇舟蛾等靶标害虫个体数量明显减少低于对照杨,转基因107杨对主要靶标害虫具有良好抑制效果;主要非靶标害虫叶蝉 sp.、皱大球蚧、大青叶蝉个体数量与对照无明显差异,白杨毛蚜个体数量低于对照,转基因107杨对非靶标害虫未产生不利影响;转基因107杨对主要天敌与中性昆虫未造成显著不利影响,部分株系异色瓢虫、狼蛛sp.、跳蛛sp.及家蝇个体数量相对于对照表现为一定程度的增多;大部分主要节肢动物种群发生高峰期集中在5-6月、8-9月,但杨白潜叶蛾7月为发生高峰期,白杨毛蚜在10月为发生高峰期。
转基因杨树为防治虫害、减少化学农药使用量方面做出巨大贡献的同时[20]其生态安全性受到广泛关注,本研究以转基因107杨为研究对象,通过研究其对主要节肢动物种群的影响为转基因杨生态安全性研究提供依据。结果表明:转基因107杨靶标害虫美国白蛾、杨扇舟蛾等个体数量明显减少,转基因107杨对主要靶标害虫具有显著的抑制效果,这与张益文等的研究结果相似[21]。昆虫之间是由食物网相连接的,转基因杨在抑制靶标害虫的同时对其他种群也有可能产生不利影响,因此研究过程中应综合考虑,且试验林地处于开放环境中,在分析时也应考虑种群的迁移和迁出。研究表明转基因107杨在抑制靶标害虫的同时,对非靶标害虫未产生显著不利影响,这与Zhang等的研究结果相似[22]。天敌在抑制害虫方面起到非常重要的作用,利用转抗虫基因杨树进行抗虫时不能对天敌产生不利影响[23],因此在对转抗虫基因杨树生态安全性进行评价时,需对其是否对天敌产生影响进行研究,本研究结果为转基因107杨对主要天敌未产生不利影响,部分天敌及中性昆虫个体数量表现为一定程度的增多,表明转基因107杨未产生毒杀作用,未对天敌及中性昆虫个体数量产生不利影响,这与姚丽等的研究结果相似[24]。部分株系昆虫数量波动较大,起伏不定可能是由于采用随机取样调查的方法、调查选取的时间、气候因素如风雨等及其它各种环境因素的变化对节肢动物种群的数量产生影响,导致节肢动物种群产生波动[25]。
转基因杨作为多年生林木,研究其对主要节肢动物种群的影响需对其进行高频率的调查与长期调监测,为科学评价转基因杨的生态安全性提供依据。
