软枣猕猴桃的光合模型筛选与光响应特征比较
2021-07-20魏炳康李根柱王贺新
魏炳康,李根柱,王贺新,娄 鑫*
(1.大连大学生命科学与技术学院,大连 116622;2.大连大学现代农业研究院,大连 116622)
软枣猕猴桃Actinidiaarguta是猕猴桃属落叶藤本植物,在中国、日本、俄罗斯远东地区、库页岛和朝鲜半岛都有分布。在中国主要集中分布于东北、华北、西北地区以及长江流域[1-2]。其果皮无毛可直接食用,果肉酸甜多汁且风味独特,营养成分丰富,尤其是维生素C含量高,能达到430 mg/100 g,具有较高的营养价值[3]。野生软枣猕猴桃幼苗通常在林下生长发育,而成年大树能生长到林冠层结果,两个生长阶段的光环境存在极大的差异。而随着软枣猕猴桃产业的发展,软枣猕猴桃的栽培方式也多种多样,有露天栽培、温室栽培以及新兴的林下栽培方式。不同栽培模式下光环境也有不同。
光环境因子是生态因子的重要组成部分,也是植物人工栽培时的主要限制因子之一[4]。在光照强度高的区域进行露天栽培时,部分植物会发生日烧现象,如苹果、桃、枇杷[5-7];在设施内或低光强的区域栽培时,又可能会由于光因子不能满足植株的正常需求从而导致植株徒长,推迟花期,减少花数,降低产量[8];而在森林生境中,又以林下光环境差异最大[9],林下光环境是林下植被发育、生态系统作用过程的关键决定性因素[10]。因此,植物对光环境因子的需求及适应,是选择植物适栽区域及栽培模式的重要因素。
光合作用是植物生长的基础,为植物的生长发育提供必要的营养物质。光合作用主要通过叶片进行,叶片的光合响应特征可通过光合响应曲线来反映,获得的光合响应参数有助于分析叶片光合机构的运转状况和利用光资源的能力[11]。目前常用的模型有直角双曲线模型(RHM)、非直角双曲线模型(NRHM)、指数函数模型(EFM)和直角双曲线修正模型(YEM)[12]。不同植物对光照强度变化的敏感程度及响应方式差异较大。所以不同的植物,其适宜的光合响应模型不一,不同模型对同一植株光合响应的拟合也存在差异[13-14]。因为光合响应参数值的合理性会影响对植株光合生理过程的判断,所以最优拟合模型的选择变得尤为重要。
近年来,有关软枣猕猴桃光合特征的研究只有少量报道。李红莉等[15]认为,不同种源的软枣猕猴桃幼苗对光的利用效率存在显著差异,因此可以将光合能力作为种源选择的关键指标。袁继存[16]研究认为:软枣猕猴桃为喜光植物,同时也能适应弱光环境。范书田等[3]认为不同软枣猕猴桃之间最大净光合速率、光补偿点、光饱和点、初始光能利用率、暗呼吸速率存在显著差异,既有很强的耐阴性,又具备喜阳植物的特性;但均未涉及利用光合模型研究软枣猕猴桃的的光合特性。因此,现利用4 种不同的光合模型对8个品种软枣猕猴桃的光响应曲线进行拟合,比较不同模型拟合的光和特征参数和实测值之间的差异,选择最适合的光合模型。同时对8种软枣猕猴桃的光响应特征进行比较,以期对软枣猕猴桃栽培提供数据支持。
1 材料与方法
1.1 试验地概况
试验地位于大连大学软枣猕猴桃资源圃。地理坐标北纬39°06′,东经121°49′,海拔90 m,是亚湿润地区,属暖温带季风气候,四季分明,气候温和,降水集中,季风明显。年平均气温约10.5 ℃,极端最高气温37.8 ℃,极端最低气温-19.13 ℃。年平均降水量 550~950 mm,日照时数2 500~2 800 h,无霜期180~200 d。
1.2 试验材料
以2种软枣猕猴桃雄株‘月山’‘森茂猕雄’,6种软枣猕猴桃雌株‘魁绿’‘LD133’‘赤焰’‘大实(大実サルナシ)’‘贵太郎(貴太郎)’‘石榴红’多年生大树为实验材料。每种软枣猕猴桃为一组,各组随机排列,株距2 m×4 m,3次重复,常规水肥管理。
1.3 测定方法
每种软枣猕猴桃选择长势中等的3株树,每株树选取阳面中层正常生长的3片健康叶作为测定叶,编号并做标记。
测定试验选择在2019年7月晴朗天气进行,于上午9:00—11:00利用美国LI-COR 公司的 LI-6400 便携式光合仪测定选取叶的光响应曲线。叶室温度设定为 25 ℃,气流速度设为500 μmol/s;光响应曲线测定时光合有效辐射(photosynthetically active radiation,PAR)设为2 200、2 000、1 500、1 300、1 200、1 000、800、500、200、100、50、20、0 μmol/(m2·s),测定植株完全展开成熟叶片的净光合速率(Pn),并利用4种光合模型对光响应曲线进行拟合,求得光响应曲线的最大净光合速率(Pnmax)、光补偿点(Ic)、光饱和点(Is)、初始光能利用率(α)、暗呼吸速率(Rd)、决定系数(R2)、均方误差(mean square error,MSE)和平均绝对误差(mean absolute error,MAE)等[17-19]。
1.4 模型选择
1.4.1 直角双曲线模型(RHM)
直角双曲线模型的数学表达式为
(1)
式(1)中:Pn为净光合速率;α为初始光能利用率;Pnmax为最大净光合速率;I为光强;Rd为暗呼吸效率。
1.4.2 非直角双曲线模型(NRHM)
非直角双曲线模型的数学表达式为

(2)
式(2)中:θ为光响应凸度,反映光响应曲线的弯曲程度,一般0≤θ≤1,θ越大,曲线的弯曲程度越大;θ越小,曲线的弯曲程度越小。
1.4.3 指数函数模型(EFM)
指数函数模型的数学表达式为
(3)
式(3)中:e为自然对数。
1.4.4 直角双曲线的修正模型-叶子飘模型(YEM)
叶子飘(YEM)模型的数学表达式为
(4)
式(4)中:β、γ为与光强无关的系数。
光饱和点用Is表示为
(5)
最大净光合速率用Pnmax表示为
(6)
1.5 模型评价指标
决定系数R2的大小可以用来判定模型方程拟合精度的高低。但是,决定系数并不足以判断各个模型的优劣,还需要均方误差(MSE)和平均绝对误差(MAE)。MSE和MAE越小,说明拟合值越接近观测值[12]。
(7)
(8)

1.6 数据分析
利用光和计算4.1.1软件完成光合响应模型拟合并计算光响应特征参数,数据处理及作图利用Excel2010和Sigma Plot 14.0完成。
2 结果与分析
2.1 软枣猕猴桃的最优拟合模型筛选
YEM的R2在4个光响应曲线模型中均为最大,MSE和MAE 值(除魁绿外)与其他3种光响应曲线模型的相比均最小。因此YEM 为软枣猕猴桃的最适光合模型(表1)。
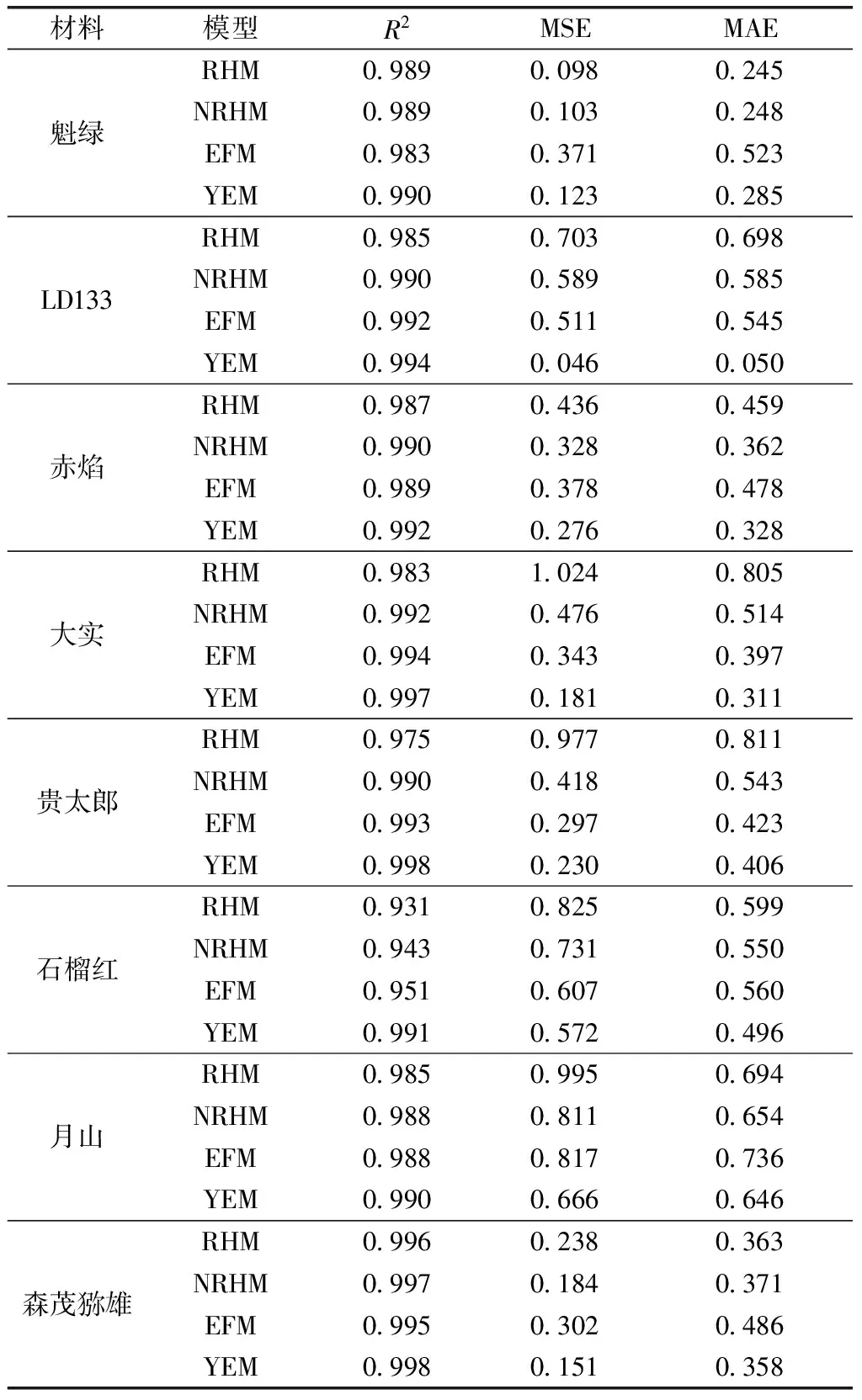
表1 4种光响应曲线模型的拟合优良度
2.2 软枣猕猴桃对光强的响应
利用YEM对8种软枣猕猴桃的光合响应曲线进行拟合。不同品种软枣猕猴桃的光响应曲线趋势基本一致,在低光照范围内[PAR≤500 μmol/(m2·s)],各软枣猕猴桃的Pn随PAR的增加而迅速上升,Pn对PAR响应敏感。随PAR 持续增强,各软枣猕猴桃的Pn缓慢增加,直至达到最大值(图1)。并且因为软枣猕猴桃的品种不同,最大净光合速率也不同(表2)。Pn增加至最大值后,叶片的Pn也不再随光强的增加而增加,尤其是 PAR≥2 000 μmol/(m2·s)时,叶片的Pn出现下降。

图1 软枣猕猴桃的直角双曲线的修正模型-叶子飘模型(YEM)光响应曲线
利用直角双曲线的修正模型-叶子飘模型对8种软枣猕猴桃的光强-光合响应曲线进行拟合,得到Pnmax、α、Rd、Ic和Is等光响应特征参数(表2)。其中森茂猕雄的最大净光合速率拟合值最大[Pnmax为19.85 μmol/(m2·s)],其次是月山[Pnmax为19.46 μmol/(m2·s)];石榴红的最大净光合速率拟合值最小[Pnmax为8.42 μmol/(m2·s)]。赤焰的暗呼吸速率拟合值最大[Rd为4.17 μmol/(m2·s)];石榴红的暗呼吸速率拟合值最小[Rd为1.44 μmol/(m2·s)],其次是森茂猕雄[Rd为1.60 μmol/(m2·s)]。LD133的光补偿点拟合值最大[Ic=56.77 μmol/(m2·s)];森茂猕雄的光补偿点拟合值最小[Ic=19.96 μmol/(m2·s)],其次是石榴红[Ic=21.16 μmol/(m2·s)]。森茂猕雄的光饱和点拟合值最大[Is=2 149.38 μmol/(m2·s)],石榴红的光饱和点拟合值最小[Is=824.43 μmol/(m2·s)]。

表2 叶子飘模型(YEM)拟合的软枣猕猴桃叶片光响应参数和观测值比较
3 讨论
利用4种光合模型对8个软枣猕猴桃品种的光强-光合响应曲线进行拟合,为保证拟合的光合响应特征参数的准确性,必须对不同的光响应模型的优劣与适用性进行分析。可用决定系数R2的大小来判定方程拟合精度的高低[11],R2越大,模型的精确度越高。同时,用模型的MSE、MAE来判别计算值与观测值之间的差异, MSE和MAE越小, 说明其计算值越接近观测值, 模型的精确度越高。在4个光响应曲线模型中,直角双曲线修正模型-叶子飘模型的R2均最大,且≥0.99;直角双曲线修正模型-叶子飘模型的MSE和MAE 是4个光响应曲线模型中最小的。所以,直角双曲线修正模型是软枣猕猴桃的最适光合模型。
直角双曲线模型、非直角双曲线模型和指数函数模型只能拟合出一条没有极值的渐近线,拟合得到的最大净光合速率远大于观测值,而且无法拟合出饱和光强。同时,这3种光合模型也不能准确地描述出净光合速率随光强增加而下降的光响应曲线。叶子飘改进的直角双曲线修正模型可以很好地解决这一问题。该模型是一条有极点的曲线,可以准确地拟合出最大净光合速率和光饱和点等光响应特征参数,同时可以用它来描述植物在光强超过饱和光强时的光响应曲线[13,20]。该模型自改进以来就被广泛应用。刘子凡等[12]在研究木薯光响应曲线模型时也得出直角双曲线修正模型是拟合木薯光响应曲线的最佳模型。还有张赟齐等[17]研究无患子的光合模型时,也认为直角修正模型能更好地拟合无患子光合响应曲线,得到的光合响应参数也比较准确。
光响应曲线是衡量植物光合作用的重要指标。不同品种软枣猕猴桃的光响应曲线趋势基本一致,在低光照范围内[PAR≤500 μmol/(m2·s)],各软枣猕猴桃的净光合速率随光强的增加而迅速上升,光合作用对低光照敏感响应。随光强持续增强,各软枣猕猴桃的净光合速率缓慢增加至最大值,达到最大净光合速率(Pnmax),该点的光强即为光饱和点。并且,因为软枣猕猴桃的品种不同,光补偿点、光饱和点、初始光能利用率值和最大净光合速率也不同。本研究拟合的软枣猕猴桃光响应曲线变化趋势与文献[3]的研究结果相一致。
当光强达到光饱和点时,净光合速率不再随着光强的增加而增大,反而出现下降现象,造成光抑制。光抑制是植物进化出的光系统保护机制,以消散过度吸收的太阳能,从而保护光合结构。所以在光照过强时要适当遮阳,避免出现日灼现象。
最大净光合速率是反映植物叶片光合的潜力。光饱和点和光补偿点直接反映了植物对光的需求和利用能力[21]。初始光能利用率可以较好地反映叶片对弱光的利用能力[22]。喜阳植物相较于耐荫植物具有较高的光饱和点和光补偿点[23]。一般喜阳植物的Is为1 000~1 500 μmol/(m2·s),Ic为10~40 μmol/(m2·s),α一般在0.04~0.07[12]。在本次研究中,8种软枣猕猴桃的Ic为20.06~56.77 μmol/(m2·s),α在0.076~0.105,Is为824.43~2 149.38 μmol/(m2·s)。结合文献[3]对软枣猕猴桃的研究成果,判定软枣猕猴桃大树有阳生植物特征。同时,在文献[3]的研究中,魁绿的光饱和点为1 590 μmol/(m2·s)。而本研究中魁绿的光饱和点为2 038.47 μmol/(m2·s)。光饱和点存在差异。这可能是地域、气候和水肥管理等差异造成的。
在本研究中,模型拟合得出α在0.076~0.105,数值相对偏高,说明其对弱光有较好的利用能力。但叶子飘等[24]认为在低光强条件下[≤200 μmol/(m2·s)]光合模型拟合的α的值具有一定的人为性,在描述植物利用光能时并不是一个理想指标,提出利用光补偿点处的量子效率来作为描述植物利用光能的一个指标。
8个软枣猕猴桃品种中具有高光饱和点的植物其最大净光合速率也相对较高;相反,具有低光饱和点的植物其最大净光合速率也相对较低。但光饱和点的高低与净光合速率大小并不呈现绝对一致性,说明植物最大净光合速率的大小不仅仅取决于光饱和点的高低,还要受到其他因子的共同影响。李林锋等[25]研究湛江红树林树种时发现,胞间CO2浓度、气孔导度、蒸腾速率、叶面大气蒸汽压亏、叶温和光照强度等都会影响树种的光合能力。而影响软枣猕猴桃光合作用的具体因素还需进一步研究。
4 结论
(1)叶子飘模型的R2在4个光响应曲线模型中均为最大,MSE和MAE 值(除魁绿外)与其他3种光响应曲线模型的相比均最小。因此最适合软枣猕猴桃的光合模型是直角双曲线修正模型-叶子飘模型。
(2)软枣猕猴桃的8个品种中,暗呼吸速率的拟合值从小到大的排序是:石榴红<森茂猕雄<月山<贵太郎<大实<魁绿
