基于系统聚类分析的黄淮海区夏大豆性状研究
2021-07-19刘博卫玲乔羽佳肖俊红杨海峰段学艳
刘博,卫玲,乔羽佳,肖俊红,杨海峰,段学艳
(山西农业大学/山西省农业科学院小麦研究所,山西临汾041000)
大豆是五谷之一.我国是大豆原产国和起源地,有悠久的饮食传统和种植历史.我国已从世界最大的大豆出口国,转变为最大的进口国.进口量从1995年的100万t左右,增长到2020年1亿t以上,25年增长约100倍,对外依存度约85%.目前,巴西、美国、阿根廷的转基因大豆约占进口大豆90%.国产大豆平均产量约在2 000 kg/hm2,非转基因大豆价格比进口转基因大豆高约60%.提高大豆产量和自给率,必须有优良品种支持,需要拓宽育种遗传基础.黄淮海地区曾是我国大豆种植面积最大区域,后因种植结构调整,玉米、花生面积增加等原因,该区大豆面积缩少至230万hm2左右,目前仍是我国大豆第二大主产区[1].该区以冬小麦收获后,复播夏大豆类型为主,大豆生育期多集中在105 d左右.该区自然生态环境复杂,造就了其大豆品种(种质)多样性丰富、适应性和抗逆性较强等特点.这些特点为丰富种质资源,拓宽遗传基础,发掘优良基因和变异,改良性状等提供了物质和理论基础.充分利于这些条件,有利于提高大豆育种品质和产量.为此,对黄淮海地区近年主要大豆品种(品系)的性状进行了分析,以期对发掘和利用优质种质,促进大豆育种及生产提供帮助.
1 材料与方法
1.1 试验材料
黄淮海夏大豆品种(系)54个,名称和供种单位见表1.
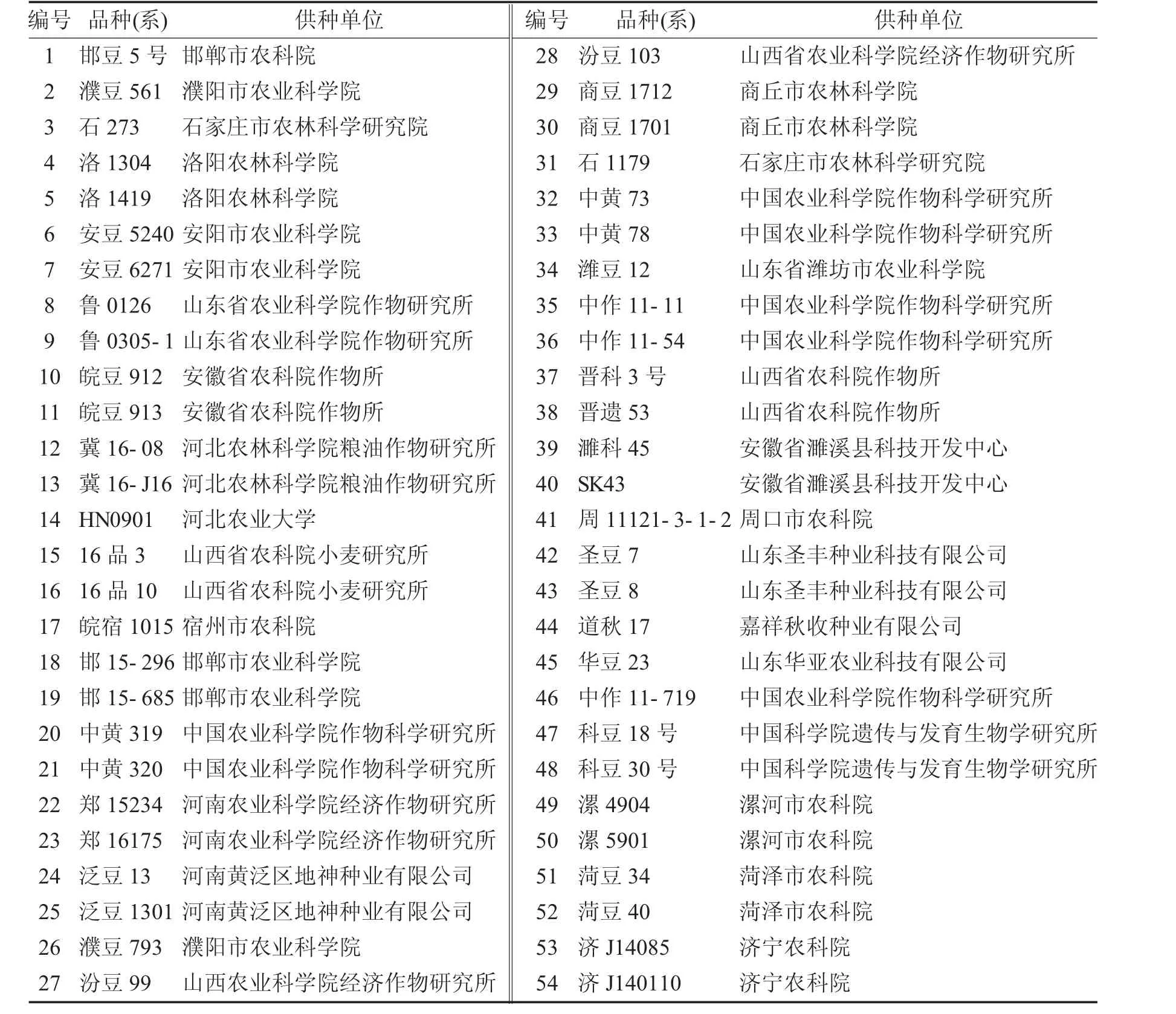
表1 大豆品种(品系)及提供单位Tab.1 varieties(lines)of soybean and their supply units
1.2 试验地概况
试验于2017年在山西农业大学/山西农业科学院小麦研究所韩村国家试验基地进行,3次重复.土壤为壤土,含速效氮量为34.61 mg/kg、含速效磷量为2.63 mg/kg、含速效钾量为265 mg/kg.前茬作物为冬小麦,秸秆还田后免耕复播种植大豆.
1.3 试验设计和方法
间比设计3次重复,小区长6 m,宽2.5 m,行距:0.5 m,株距0.11 m,密度18.75万株/hm2,其他各项操作均参照国家区试常规要求种植.
使用Excel 2007和SPSS PASWStatistics18进行数据处理和分析.
2 结果与分析
2.1 大豆群体性状描述分析
由表2可知,参试大豆品种(系)性状变异丰富.从极值增幅(极差/极小值)看,各性状表型变异从大到小可分为4类.第1类:增幅4倍以上,即无效荚数、单株粒质量、有效荚数、单株荚数,主要为库容因子;第2类,增幅3倍以上:单株粒数、分枝数、底荚高;第3类,增幅1倍以上:株高,主要为株型因子;第4类,增幅0.5倍以上:节数、百粒质量、产量.从变异系数看,可分为3类,第1类:无效荚数、分枝数位列前2,超过70%;第2类:单株粒数、单株荚数、有效荚数、单株粒质量、底荚高超过35%;第3类:株高、百粒质量、产量、节数,超过10%.

表2 大豆品种(品系)性状描述Tab.2 Traits description of soybean varieties(lines)
2.2 大豆性状间相关系数
对参试大豆品种(系)性状进行相关分析,结果见表3.
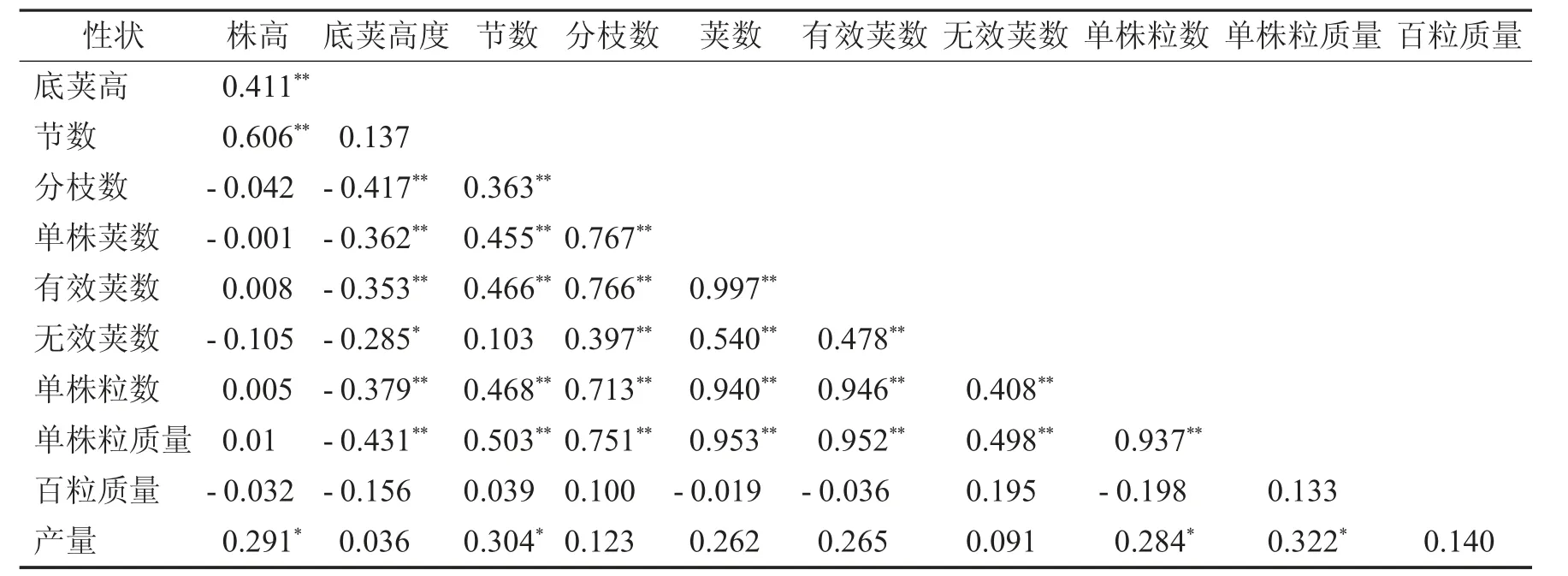
表3 大豆品种(品系)性状相关系数Tab.3 Related Traits coefficient of soybean varieties(lines)
株高与底荚高、节数、呈极显著正相关,与产量呈显著正相关;底荚高与单株粒质量、分枝数、单株粒数、单株荚数、有效荚数呈极显著负相关、与无效荚数呈显著负相关;节数与单株粒质量、单株粒数、有效荚、单株荚数、分枝呈显著正相关,与产量呈显著正相关;分枝数与单株荚数、有效荚、单株粒质量、单株粒数、无效荚呈显著正相关;单株荚数与有效荚数、无效荚数、单株粒数、单株粒质量呈极显著正相关;有效荚与无效荚、单株粒数、单株粒质量呈极显著正相关;无效荚与单株粒数、单株粒质量呈极显著正相关;单株粒数与单株粒质量呈极显著正相关、与产量呈显著正相关;单株粒质量与产量呈显著正相关.
2.3 大豆性状与产量回归分析
大豆性状与产量进行回归分析见表4和表5.
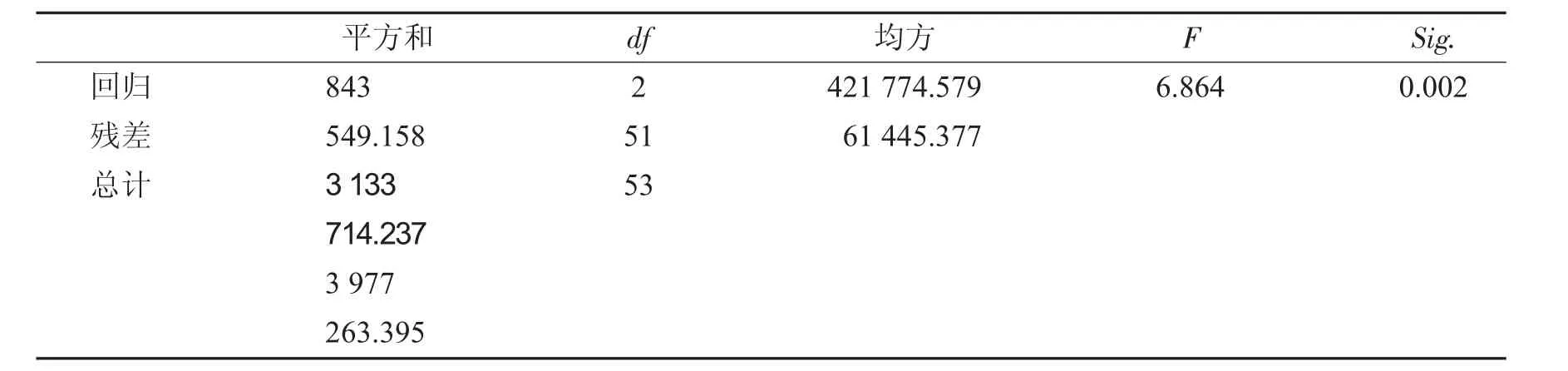
表4 大豆性状与产量线性回归Tab.4 Traits of soybean and output linear regression
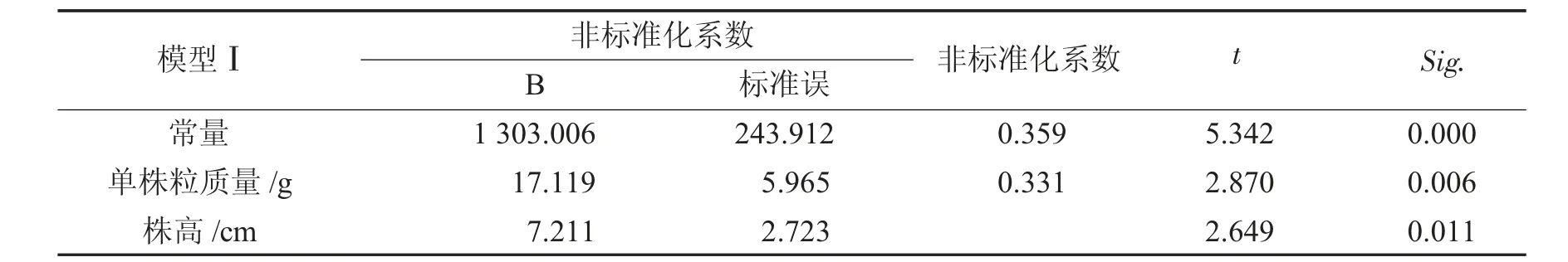
表5 大豆产量线性回归模型Tab.5 Linear regression model of soybean ou t
由表4和表5可知,产量回归方程为:y=1 303.006+17.119x9+7.211x1sig.=0.002<0.01达极显著水平,模型有效.单株粒质量对产量影响极显著、株高对产量影响显著.
主因子特征值、贡献率及累计贡献率,见表6.
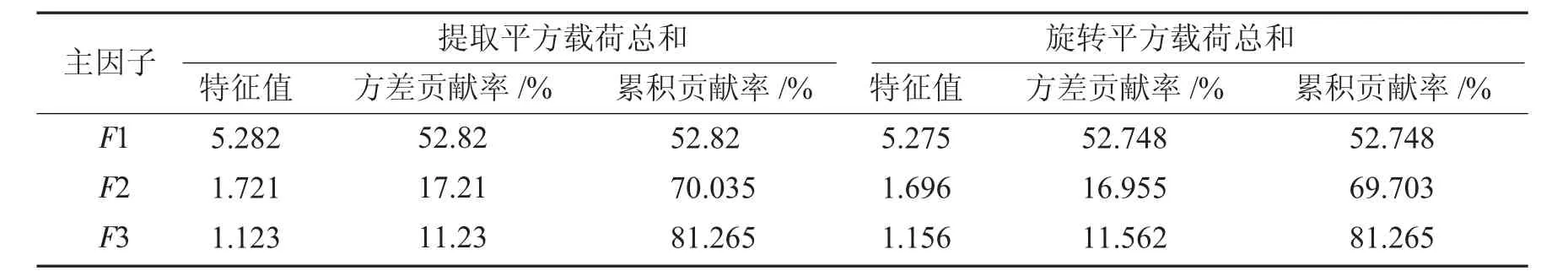
表6 主因子特征值、贡献率及累计贡献率Tab.6 Main Traits value,rate of contribution,accumulative total rate of contribution
由表6可知,3个主因子特征值均大于1,且累计贡献率81.265%,可反映原始表型因素的绝大部分信息.
2.4 大豆性状因子分析
旋转后载荷矩阵和大豆品种(系)性状主因子得分、综合得分及排序(前3和后3)见表7和表8.
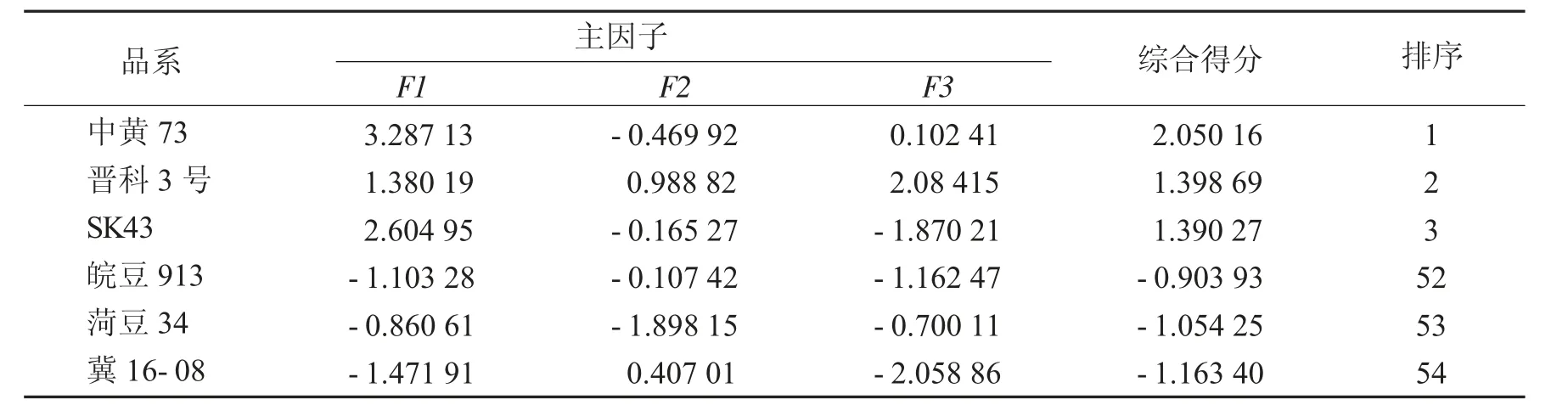
表8 大豆品种(系)性状主因子得分、综合得分及排序Tab.8 Scores of main factors,comprehensive scores and rank of soybean varieties(lines)
由表7可知,第1主因子主要由单株荚数、有效荚数、单株粒质量、单株粒数、分枝数构成,均与光合产物存贮的库有关,可称为库容因子;第2主因子主要由株高、节数、底荚高度构成,均与株型和植株高度有关,可称为株型/高度因子;第3主因子主要由百粒质量和无效荚数组成,遗传较稳定,可称为稳定因子.可见贡献是库容因子>植株因子>稳定因子.
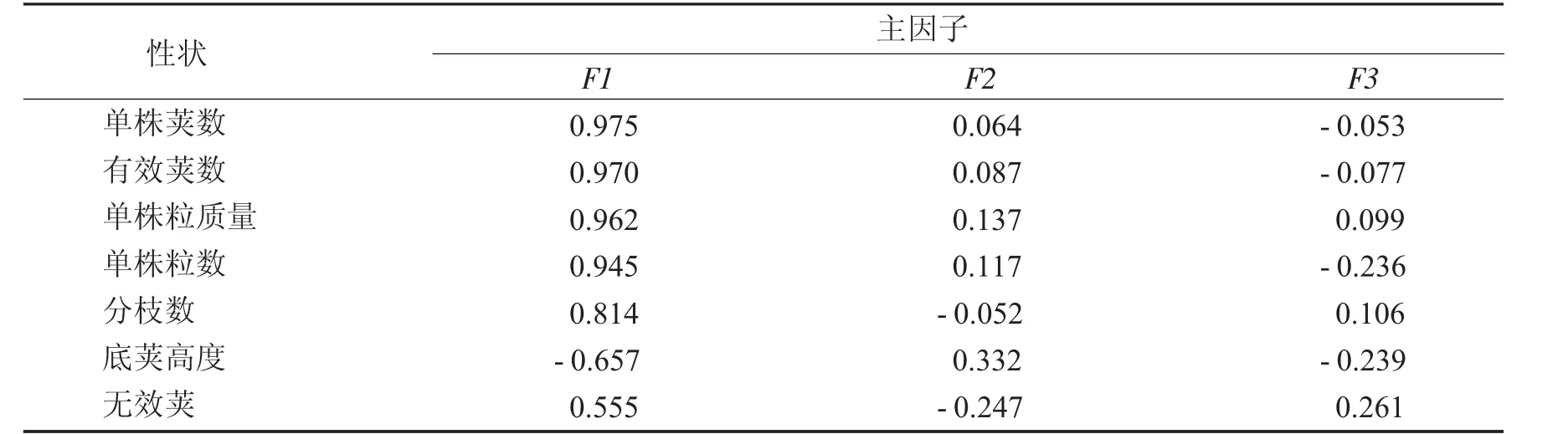
表7 旋转后载荷矩阵Tab.7 Loading matrix after spin
由表8可知,大豆各品种(系)主因子得分不同,综合得分中黄73、晋科3号、SK43居前3位,皖豆913、荷豆34、冀16-08居后3位.中黄73等可作为重点亲本用于育种.
2.5 大豆群体性状聚类分析
大豆群体性状系统聚类如图1所示。
使用系统聚类对参试大豆品种(系)性状进行了聚类分析.由图1可知,在距离值为9时,可分为4个群:中黄319、泛豆13号、邯豆5号等11个为第1群,主要分布在北京、河南、河北等;郑15234、郑16175、濮豆561等6个为第2群,主要分布在河南、安徽、北京;安豆5240、中作11-54、周11121-3-1-2等20个为第3群,黄淮海各地均有分布;科豆30号、漯4904、汾豆99等17个为第4群.
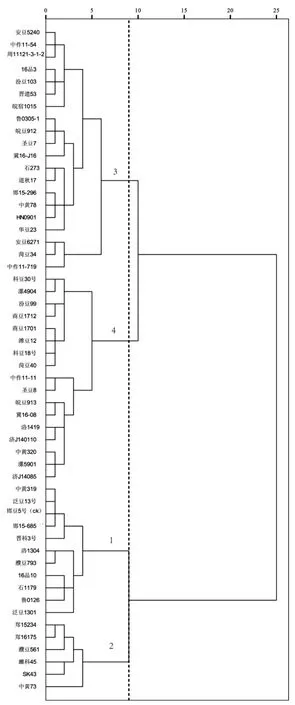
图1 大豆群体性状系统聚类Fig.1 System cluster of soybean group traits
不同类群性状均值比较见表9.
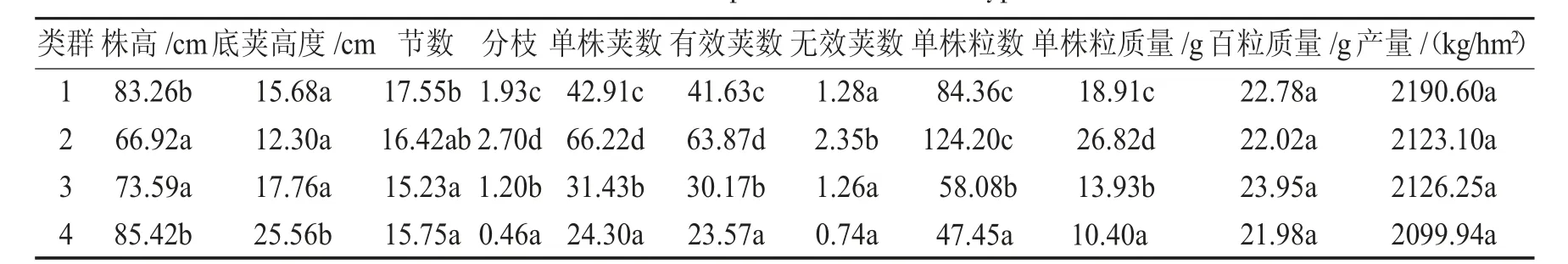
表9 不同类群性状均值比较Tab.9 Mean value comparison of different types of traits
表9显示,在4个群体中,第4群、第1群体之间株高无显著差异,但显著高于第3和第2群(后2个之间无显著差异);第4群的底荚高度显著高于其他群体(其他群之间无显著差异);第1群的节数显著高于第3、第4群(第3、第4之间无显著差异);单株荚数、单株有效荚数、单株粒质量4个群体间差异均显著,且顺序一致,均为第2群最高,第4群最低;无效荚数第2群也显著高于其他群体(其他之间差异不显著);单株粒数第2群最高,第4群最低.百粒质量和产量在4个群体间差异不显著.第1群产量最高.
3 讨论
在黄淮海夏大豆54份大豆性状方面,研究了相同环境下,不同来源的品种(系)总体表现.结果发现,群体性状主要农艺性状,按极值增幅可分为4类,按变异系数可分为3类,2种分类方法均显示,无效荚、分枝数、单株粒数、单株荚数,有效荚数在群体中变幅较大,节数、百粒质量在群体中变幅较小,但也有主茎节数变异系数较大的报道[2].因此,在育种工作中,需要重视对荚数、单株粒数、分枝数等库容因子进行适当选择,以提高育种效率.
本研究中,百粒质量变幅较小,且4类群体间均值差异不显著.大豆百粒质量受到籽粒大小影响显著.大豆籽粒大小的遗传受多基因控制,以基因加性效应为主,属数量性状遗传,百粒质量的遗传是相对稳定[3,4],氮、磷、灌溉等对百粒质量影响也不显著[5].潘丽媛等利用限制性两阶段多位点全基因组关联分析方法(RTM-GWAS),对大豆RIL群体百粒质量进行了QTL检测,相比复合区间作图法和基于混合线性模型的全基因组关联分析方法2种方法,RTM-GWAS检测QTL数目最多,解释表型变异最多[6].陈强等研究发现,在石家庄国内外的205份大豆资源材料中,百粒质量遗传力为96.88%[7].宋世玉报道,百粒质量性状的广义遗传率在85%以上[8];孙亚男等报道了16个与大豆百粒质量性状相关的QTL[9].Lu等报道,PP2C-1优异等位基因的引入可提高大豆粒质量或大小[10].另有研究表明,大豆籽粒的单粒质量呈现明显的粒位效应.在多粒荚内,基粒的粒质量最小.不同粒位粒质量在主茎上呈明显的“纺锤形”分布,下部节位籽粒单粒质量略小,而中部节位籽粒单粒质量较大.不同粒位的籽粒大小与灌浆顺序并无直接关系[11].有研究表明,对粒形性状和百粒质量主效QTL检测,获得81个相关QTL,分布于18条染色体,贡献率1.66%~30.70%,主效QTL对粒形性状遗传影响最大,上位性次之,环境互作最小[12-13].
在大豆性状相关性方面,本研究结果显示,单株粒质量、节数、株高、单株粒数与产量呈显著正相关,且相关系数依次减小.底荚高度与分枝数、荚数、单株粒数、单株粒质量均呈显著负相关.百粒质量与其他9个性状相关性未达显著水平.前人对株高研究发现,13个候选基因与大豆株高有关,9个基因被注释为编码生长素响应蛋白,3个基因被注释为编码脱落酸相关蛋白,1个基因为GH3生长素响应启动子[14].在株高研究方面,目前已有报道构建基于Kinect 2.0的大豆冠层图像同步采集平台,并在三维重建大豆冠层结构形态基础上,提出了基于深度信息的个体和群体大豆株高计算方法[15],有望高通量,客观采集株高信息,减轻对劳动力依赖.
在大豆产量方面,周长军研究发现除单株粒数、单株粒质量外,荚期植株鲜质量与大豆产量呈极显著正相关,应关注荚期植株鲜质量[16].李灿东等发现在高密度条件下,有效分枝数与大豆产量密切相关[17].单株荚数宜在低世代选择,株高、主茎节数、单株粒数、单株粒质量和百粒质量宜在高世代选择,环境条件对株高和百粒质量的选择效果影响较大[18].小区产量和底荚高、分枝数、荚数等呈显著或极显著正相关[19].本研究对大豆性状与产量进行了回归分析,发现单株粒质量对产量影响极显著、株高对产量影响显著.试验条件、研究对象和试验方法等不同,可能是造成研究结果大同小异的主要原因,但普遍认为单株粒质量对产量有显著影响.产量与单株粒数极显著相关[20-21],与本研究结果也完全吻合.
本研究对大豆性状进行了因子分析,结果发现,3个主因子特征值大于1,且累计贡献率81.265%,反映大部分信息.因子贡献表现为:库容因子(单株荚数、有效荚数、单株粒质量、单株粒数、分枝数)>植株因子(株高、节数、底荚高度)>稳定因子(百粒质量和无效荚数),分别反映约50%、20%、10%信息.与吕美琴对福建省11个春大豆品种研究前3个主成分,分别为荚粒性状因子(单株荚数、单株粒数)、产量性状因子(百粒质量、单株粒质量)和株型性状因子(主茎节数)结果相近[2].本研究对主因子得分进行了综合排序,中黄73、晋科3号、SK43前3位,皖豆913、荷豆34、冀16-08居后3位.综合排序可作为育种组培亲本开展育种工作的参考.
本研究对大豆群体性状进行了系统聚类分析,结果显示,群体可分为4个.群体间株高、底荚高度、节数、无效荚数、单株粒数存在显著差异;单株荚数、单株有效荚数、单株粒质量在4个群体间差异均显著,且顺序一致,均为第2群最高,第4群最低;百粒质量和产量在4个群体间差异不显著.对应到产量上,群体株高最高或群体单株粒质量最高,只有单项最优的群体,产量均不是最高,而是两者协调较高,没有明显短板群体,平均产量最高.不同类群的划分,可作为区分品种(系)相似度划分依据,可为配置杂交组合提供帮助.
4 结论
对黄淮海54份不同来源大豆参试品种(系),在同一环境下群体进行了性状研究.11种性状,从极值增幅和变异系数分类结果看,库容因子(荚数、粒数等)变幅较大,株高、百粒质量、产量、节数变幅较小.
从相关性分析看,产量与单株粒质量、节数、株高、单株粒数呈显著正相关,相关系数依次降低.
对产量进行回归分析表明,单株粒质量对产量影响极显著、株高对产量影响显著.
因子分析显示,3个主因子特征值均大于1,且累计贡献率81.265%,可反映原始表型因素的绝大部分信息.对表型变异贡献依次是库容因子、植株因子、稳定因子,分别占比约50%、20%、10%.
利用系统聚类,将该群体在距离值为9时,划分为4个群.群体11个表型均值均有不同程度差异,可根据需要,在群体中选择目标性状优良品种(品系)进行优良性状聚合育种.
在大豆育种工作中,若以高产为育种目标,不但要重视对库容因子的选择,而且要重视植株因子协调.节数在群体中显示变幅较小,可适当减少关注.通过对群体主要性状间相关性、主因子、系统聚类等研究,为育种和性状研究,推动精准育种和进一步开展相关性状研究提供帮助.本研究仅在表观性状上,使用多种方法对本区域大豆品种(系)进行表观性状初步探索,在遗传、分子、转录、翻译、蛋白、细胞、生理生化等相关机理机制等深层研究方面还需要加强.
