玉米dhn1和dhn2基因渗入普通自交系的分子标记辅助选择研究
2021-07-16潘金卫杨文鹏柏光晓
潘金卫, 邓 磊, 王 伟, 杨文鹏, 任 洪, 柏光晓
(1.贵州大学农学院, 贵阳 550025; 2.贵州省农业科学院旱粮研究所, 贵阳 550006)
玉米是世界上主要的饲料和粮食作物,其生长发育受到各种环境因素的影响。其中干旱是影响玉米生长的主要限制因素。中国是水资源十分短缺的国家之一,干旱、半干旱耕地面积占总面积的52.5%[1],每年受旱面积达2.0×107~2.7×107hm2,平均每年玉米因干旱而造成的产量损失达15%~20%[2]。在我国西南地区,春旱和伏旱较为严重[3],而玉米散粉吐丝期需水量较大,伏旱是导致减产最主要的因子之一。
为提高玉米的耐旱性,人们在耐旱基因表达[4]、耐旱相关基因的克隆及功能鉴定等方面做了大量的研究工作[5],分离和鉴定了耐旱相关的一些候选基因及蛋白[6],并对相关基因进行了遗传转化[7]。Mao等[8]在玉米第10染色体上的GRMZM2G127379(ZmNAC111)基因上游572 bp处启动子区发掘一个82 bp的MITE插入位点,普遍存在于温带玉米种质中,而热带玉米种质中缺失,并且证明与玉米苗期耐旱性有关。Wang等[9]发掘ZmVPP1基因组中的遗传变异,并证明可以有效提高玉米苗期的耐旱性。玉米的耐旱性由数量性状位点(QTL,Quantitative Trait Locus)控制[10],目前,定位的耐旱QTLs涉及形态学性状[11]、产量性状及其产量构成因子[12]、开花-吐丝间隔期(ASI)[13]等,以及在干旱胁迫下可能产生遗传变异的一些生理生化性状[14]。 脱水素(dehydrin,DHN)是LEA蛋白(late embriogenesis abundant protein,LEA)中的一类,它具有较强的亲水性,在植物响应非生物逆境环境如干旱、低温冻害、高盐碱等胁迫过程中起着重要的作用[15]。玉米中与脱水素有关的DHN基因有8个(http://www.maizegdb.org/),对干旱胁迫处理表现出不同程度的响应,其中,Zmdhn1和Zmdhn2基因在玉米各个组织中均有稳定表达[16-18],通过调控DHN基因的表达,可以显著提高植物的抗逆性。Liu等[19]根据dhn1中的单核苷酸A/G多态性,开发了CAPS 标记dhnC 397,可以用于玉米的耐旱评估。这些研究均为利用分子标记辅助选择玉米耐旱种质提供了可能。
为进一步拓宽玉米耐旱性种质资源,本研究以引进的玉米耐旱材料和普通玉米自交系为试材,利用常规育种和分子标记辅助选择技术,将耐旱主效基因dhn1与dhn2渗入到普通玉米材料中,以期获得玉米耐旱性种质,为玉米抗旱育种提供参考。
1 材料与方法
1.1 供试材料与群体构建
以2份引自CIMMYT的玉米耐旱材料,3份普通玉米自交系为试材,具体见表1。
利用耐旱性玉米材料和普通玉米自交系杂交获得F1,用普通自交系回交获得BC1F1,在BC1F1世代于盛花期进行人工混合授粉(每个基础群体混粉约50个单株)获得BC1F1/S1,共构建6个回交群体:CML 538/QR 273//QR 273、 CML 538/PH 6 WC//PH 6 WC、CML 538/LX 9801//LX 9801、CML 539/QR 273//QR 273、CML 539/PH 6 WC//PH 6 WC、CML 538/LX 9801//LX 9801。
1.2 分子标记
对于耐旱dhn1基因,参照柳思思等[20]开发的功能性CAPS分子标记dhnC 397进行检测,其中dhn1基因序列397位点发生了G与A的转换,PCR扩增产物经StyI酶切后,含有G位点的目的片段被酶切成131 bp和33 bp两个片段,而含有A位点的由于缺少StyI酶切识别位点,因此不能被酶切,不被酶切的大片段与玉米的耐旱性关联,被酶切的小片段与玉米的不耐旱性关联。对于耐旱dhn2基因,选用与dhn2连锁的SSR标记umc 1634进行检测(表2)。

表2 耐旱基因及分子标记信息
1.3 DNA提取及PCR扩增
采用分子标记辅助选择的玉米微量DNA分离提取的CTAB法提取DNA[21]。用超微量紫外分光光度计(NanoDropTMOne/OneC,产地美国)检测DNA的浓度与质量,将DNA样品浓度稀释至10 ng·μL-1,-20 ℃下保存。
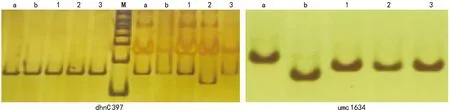
注:M为DL 2000 Marker;a为CML 538;b为CML 539;1为QR 273;2为PH 6 WC;3为LX 9801。图1 dhn1和dhn2目的基因标记在亲本之间的多态性Fig.1 Polymorphism of markers for dhn1 and dhn2 between parents

注:1为供体亲本CML 538;2为轮回亲本QR 273;3为3~32为世代群体的扩增条带。图2 利用umc 1634标记在玉米BC1F1/S1世代中部分单株扩增结果Fig.2 Amplification of some single plants in maize BC1F1/S1 generation by umc 1634 marker
PCR扩增反应体系(10 μL):模板DNA 3 μL,10×Buffer 1 μL,MgCl2(25 mmol·L-1)1 μL,Glycerol(99%)1 μL,dNTP(2 mmol·L-1)0.75 μL,Primer(F+R)1.2 μL(5 μmol·L-1),Taq(5 U·μL-1)0.075 μL,最后补ddH2O至10 μL。PCR扩增程序在BIO-RAD C 1 000Thermal Cycler型扩增仪上进行。PCR扩增程序为:93 ℃预变性1 min;93 ℃变性1 min,58 ℃退火2 min,72 ℃延伸2 min,35个循环;72 ℃延伸10 min;4 ℃保存。针对dhn1基因,PCR反应程序参照柳思思[20]的操作方法,PCR产物,利用限制性内切酶StyI进行酶切,温度为37 ℃,时间为4 h。用8%聚丙烯酰胺凝胶,对PCR产物进行检测。
1.4 靶基因选择与背景分析
对BC1F1/S1分离世代,于苗期提取的基因组DNA,利用CAPS标记dhnC 397,和SSR标记umc 1634进行靶基因选择。对获得的中选株系,用40对SSR标记(参考标准NY/T 1432-2014)进行遗传背景分析。用于靶基因选择和背景分析的SSR标记引物的序列来源Maizegdb网站(http://www.MaizEgdb.org/),由上海捷瑞生物工程公司合成。
1.5 萌发期的耐旱性鉴定
参照《玉米抗旱性鉴定技术规范》(DB 13/T 1282—2010)选取整齐一致、颗粒饱满、无破损的玉米种子进行萌发期的抗旱鉴定。每份材料选取25粒,共3次重复。
1.6 统计分析
理论背景回复率计算公式:E[G(g)]=1-(1/2)g+1;基于分子标记的背景回复率计算公式:G(g)=[L+X(g)]/2L[22-23],式中G(g)为在g代的遗传背景回复率;g为用于回交的世代次数;L为参与分析的分子标记数量;X(g)为回交g代中表现为与轮回亲本相同带型标记的数量。用Microsoft Excel 2010软件和SPSS 23软件对相关数据进行统计分析。
2 结果与分析
2.1 多态性引物筛选
利用CAPS标记dhnC 397和SSR标记umc1634对供试材料进行多态性筛选。结果表明,在dhn1位点,PH 6 WC的PCR扩增产物可被StyI酶切,表明PH 6 WC在dhn1位点不具有耐旱性,而CML 538、CML 539、QR 273、和LX 9801在dhn1位点的PCR扩增产物不能被StyI酶切,表明这些自交系在dhn1位点具有耐旱性。在dhn2位点,连锁标记umc 1634在CML 538、QR 273、PH 6 WC和LX 9801的扩增产物均有多态,CML 539与PH 6 WC、QR 237和LX 9801的扩增产物均有多态(图1)。
2.2 靶基因选择结果
在BC1F1/S1分离世代,于苗期利用CAPS标记dhnC 397和SSR标记umc 1634进行靶基因选择(图2)。构建的6个群体共586个单株,共中选了56个单株,根据田间植株表现,最终收获22份株系。以PH 6 WC为轮回亲本的2个群体,中选24个单株,实际收获8株;以QR 273为轮回亲本的2个群体,中选19个单株,实际收获7株;以LX 9081为轮回亲本的2个群体,中选13个单株,实际收获7株,其中获得渗入dhn1基因的株系共1份,编号为NH 13;同时渗入dhn1与dhn2基因的株系共2份,编号为NH 4和NH 16;其余19个株系都渗入了dhn2基因(表3)。后继续种植收获的22份株系自交加代直至稳定。
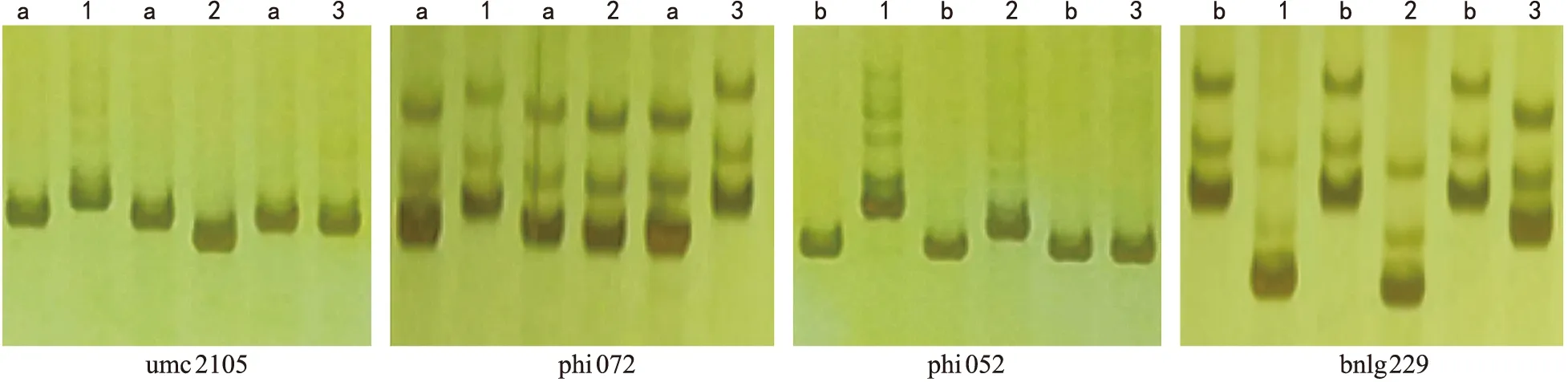
注:a为CML 538;b为CML 539;1为PH 6 WC;2为QR 273;3为LX 9801。图3 部分SSR标记在亲本间多态性的筛选Fig.3 Screening of polymorphism among parents with partial SSR markers

表3 BC1F1/S1世代目标基因选择结果
2.3 遗传背景分析结果
用40对SSR标记对获得的稳定株系进行遗传背景分析(表4)。结果表明,各群体收获材料的背景回复率介于72.5%~92.5%之间,平均值为82.78%,CML 538/PH 6 WC//PH 6 WC群体最高,达到84%,比理论值高9%;CML 539/PH 6 WC//PH 6 WC群体最低,为81.17%,比理论值高6.17%。在22份材料中,除了CML 538/LX 9801//LX 9801群体的NH 15,遗传背景回复率为72.5%,低于理论背景回复率75%,其余都高于75%,最高为CML 538/PH 6 WC//PH 6 WC群体的NH 4,高达92.5%;最低为CML 538/LX 9801//LX 9801群体的NH 9,低至72.5%。
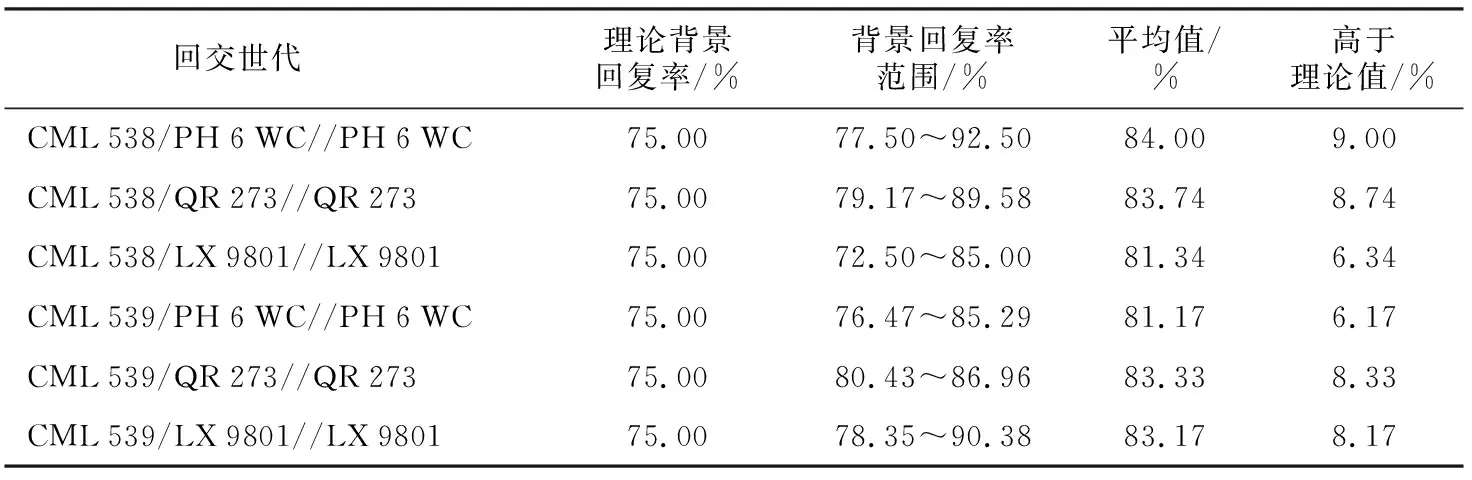
表4 22份材料背景回复率简单统计
2.4 中选材料的耐旱性鉴定
参考DB 13/T 1282-2010标准,在渗透压为-0.50 MPa的PEG-6000干旱胁迫下,对22份材料进行萌发期的耐旱性鉴定。结果表明,种子萌发生长明显受到抑制,对照组和实验组种子最终萌发率存在极显著差异(p<0.01)。22份材料的耐旱指数介于45.54%~71.07%之间,耐旱指数最高的为NH 17,高达71.07%;最低为NH 2,低至45.54%,其中,4份材料为弱耐旱型,17份材料为中等耐旱型,1份材料为强耐旱型(表5)。NH 4与NH 16同时渗入耐旱dhn1和dhn2基因,耐旱指数分别为61.23%和67.05%,明显高于同一群体的其他收获材料,说明耐旱基因dhn1和dhn2同时渗入时,对耐旱性具有一定累加作用。
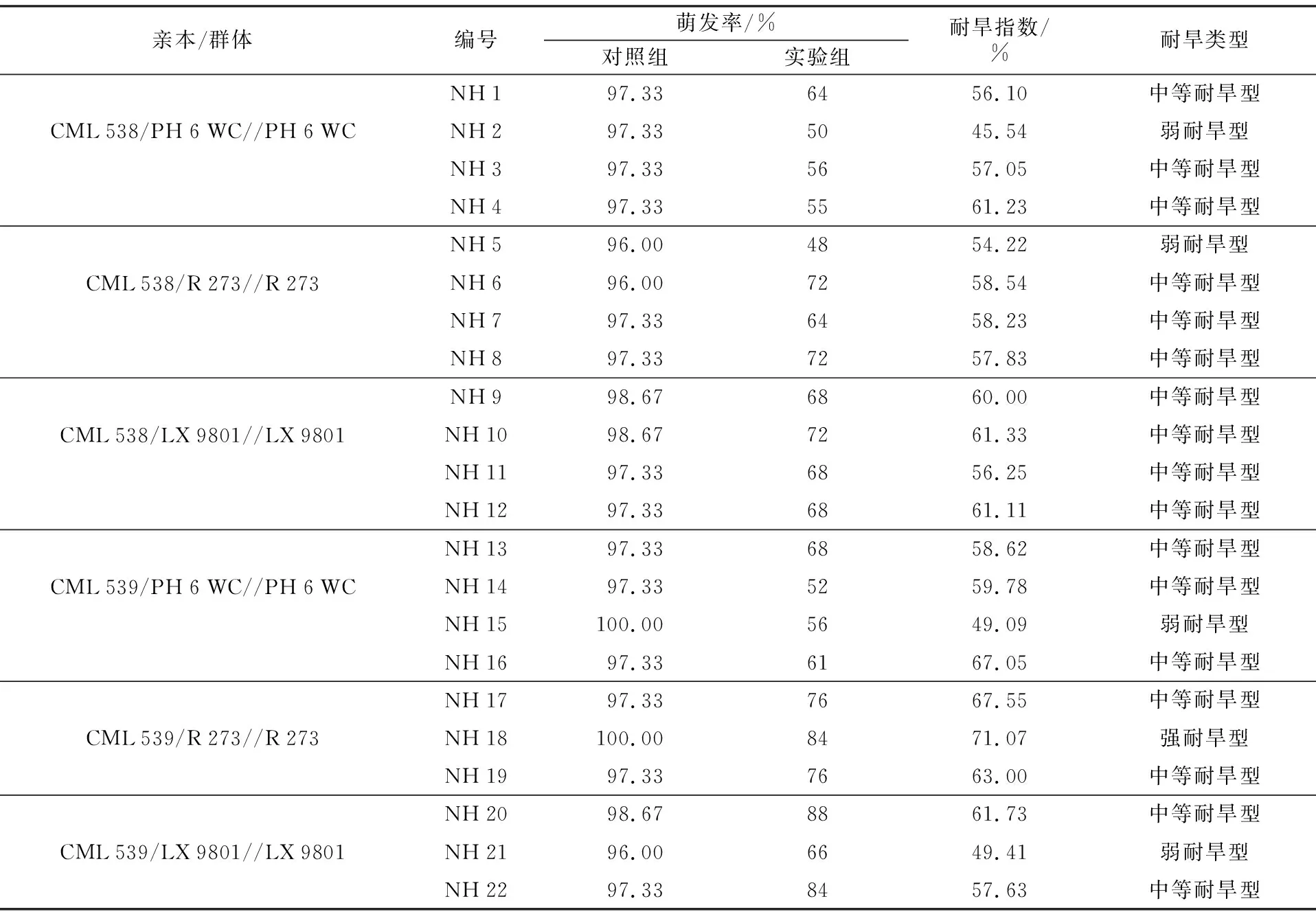
表5 22份材料萌发期的耐旱性鉴定结果
3 结论与讨论
本研究以引进的耐旱性玉米材料作为供体,与普通玉米自交系作为受体构建了6个回交群体,利用MAS进行靶基因选择,将耐旱主效基因渗入普通玉米自交系中,共获得22份材料,根据萌发期的初步抗旱鉴定,18份材料达到中等及以上耐旱性,可以为玉米抗旱育种提供材料支撑。
为提高玉米耐旱性,Ribaut等[24]使用分子标记辅助育种,将玉米开花吐丝间隔期较短的耐旱供体Ac 7643中5个ASL QTLs,通过回交转育渗入到优良热带干旱敏感受体CML 247中,获得了开花吐丝间隔期较短的玉米材料。柳思思[20]利用CAPS功能标记dhnC 397,对210份自交系进行分子筛选,结果证明,138份材料具有抗旱性,与210份自交系基本抗旱信息一致。Liu等[25]通过对热带和温带地区的368个玉米品种候选基因进行关联分析,找到了与玉米抗旱有关的基因ZmDREB,通过回交导入与分子辅助等方法,进一步证明该基因的表达可以增强玉米的耐旱能力。耐旱主效基因或是调控基因对耐旱的强弱具有决定性作用[26]。本研究利用MAS技术,将耐旱性主效QTL(或基因)dhn1与dhn2渗入普通玉米自交系中,经耐旱性鉴定,获得的18份中等及以上耐旱性材料,对耐旱玉米的分子改良具有参考作用。
种子萌发期模拟干旱处理是耐旱性鉴定的常用方法。郭效龙等[27]以抗旱性较强的玉米自交系郑58和不抗旱自交系E 28为试材,利用不同浓度PEG-6000水溶液在萌发期模拟干旱胁迫,结果表明,郑58的种子萌发率、萌发耐旱指数均高于E 28。贾凯旋等[28]以河北省推广面积较大的37个玉米品种为试验材料,采用渗透压为-0.4 MPa的PEG-6000溶液在萌发期模拟干旱胁迫,通过萌发指数进行分级,获得强抗耐旱型及以上的品种10份。姚玉波等[29]以35份玉米自交系为试验材料,采用渗透压为-0.4 MPa的PEG-6000溶液模拟干旱胁迫,与对照组比较,种子最终萌发率存在极显著差异,通过对萌发指数的分析,鉴定出抗旱型材料4份。本研究参考DB 13/T 1282-2010标准,在渗透压为-0.50 MPa的PEG-6000干旱胁迫下,对中选的22份材料进行萌发期的抗旱鉴定,与前人研究结果一致,但由于选取的处理浓度(渗透势为-0.50 MPa)较高,因此本实验耐旱指数研究结果与之相比偏低,导致抗旱类型划分略有差异。玉米耐旱性是由多种数量性状和质量性状复合遗传因子组成,十分复杂,在创制耐旱种质时,不但要考虑耐旱主效QTL位点(或基因)的分子标记检测的准确性,还应考虑对获得的中选株系进行耐旱性鉴定,二者结合起来,才能获得更准确的耐旱性种质。
