水稻叶绿体基因CPN60-β多克隆抗体的制备
2021-07-16兰汉红陈海煌黄伟鑫何昕翼
兰汉红,陈海煌,黄伟鑫,何昕翼
(闽南师范大学生物科学与技术学院,福建 漳州 363000)
0 引言
【研究意义】水稻条纹叶枯病是由水稻条纹病毒(Rice stripe virus, RSV)引起的重大病害,曾在中国16个省市和自治区暴发,给水稻生产带来巨大危害,严重威胁着我国农业生产和粮食安全[1]。然而,当前对该病毒病的防控仍缺乏有效的措施。RSV属于纤细病毒属(Tenuivirus)病毒,其基因组为单链RNA(singal strain RNA, ssRNA)。对RSV的编码策略研究表明,其基因组RNA1为负链编码,RNA2、RNA3和RNA 4均为双义编码。RNA1互补链编码复制相关蛋白RdRp[2];RNA2毒义链及其互补链分别编码沉默抑制子蛋白NS2和介体传毒相关蛋白NSvc2[3];RNA3毒义链及其互补链分别编码沉默抑制子NS3和外壳蛋白CP[4−5];RNA4毒义链及其互补链分别编码病害特异蛋白SP和运动蛋白NSvc4[6−7]。病毒编码的蛋白较少,为了完成复制循环,病毒常与寄主的蛋白因子相互作用。因此,加深RSV与寄主的互作对于病毒病害的绿色防控具有重要意义。【前人研究进展】Liu等[8]研究发现,小麦矮缩病毒的复制酶蛋白A与小麦文库中的GRAB互作,GRAB的表达抑制小麦矮缩病毒在小麦种的复制增殖;唐超等[9]研究发现,小西葫芦黄花叶病毒RNA多聚酶蛋白能够与PABP氨基酸C端特异互作;Soellick等[10]研究发现,番茄斑萎病毒编码的运动蛋白NSm能够与烟草和拟南芥的DnaJ蛋白特异互作;Li等[11]研究发现,番茄花叶病毒的外壳蛋白能够与激发子反应蛋白特异互作,该蛋白基因沉默后,病毒的外壳蛋白无法长距离运动,这结果说明两者的特异互作对于病毒的运动是必须的;Desvoyes等[12]研究发现,番茄丛矮病毒的运动蛋白P22能够与亮氨酸拉链同源结构区蛋白特异互作,病毒的感染能够显著提高蛋白基因的表达水平。【本研究切入点】前人研究表明,RSV编码的运动蛋白NSvc4在模式植物烟草中参与病毒的长距离、胞间运动以及本氏烟感病症状的形成[8,13−15],证明NSvc4参与病毒的运动和致病等侵染过程。但是NSvc4的运动或致病机制仍然不是很明确。在前期的酵母双杂交试验中,笔者发现NSvc4与水稻叶绿体相关的一个蛋白-RuBisCO亚基结合蛋白Chaperonin-60-beta(CPN60-β)能够特异互作。猜测CPN60-β蛋白在NSvc4蛋白的运动和致病过程中发挥重要作用。但是,目前还缺乏针对CPN60-β的特异抗体,阻碍了对CPN60-β蛋白在细胞内定位情况、如何发挥功能等相关问题的探讨。【拟解决的关键问题】制备与RSV运动和致病过程有关的NSvc4蛋白的互作蛋白水稻叶绿体蛋白的RuBisCO亚基结合蛋白Chaperonin-60-beta(CPN60-β)多克隆抗体。明确CPN60-β基因原核表达条件,获得水稻叶绿体CPN60-β蛋白的多克隆抗体,用于后续研究CPN60-β蛋白在RSV编码的NSvc4蛋白介导的病毒运动和致病过程中的调控功能及作用机制。
1 材料与方法
1.1 主要材料
原核表达载体pET-28a(+)由本课题组实验室保存。植物组织总RNA快速提取试剂盒(RNA Easy Fast Plant Tissue Kit)和 质 粒 小 量 快 速 抽 提 试 剂 盒(TIANprep Rapid Mini Plasmid Kit)等购自北京天根生化科技有限公司;限制性核酸内切酶和聚合酶、逆转录试剂盒(FastKing One Step RT-PCR Kit)购自美国Thermo Fisher Scientific公司;弗氏佐剂购自美国Sigma-Aldrich公司;鼠源6×His单克隆抗体购自美国Proteintech Group公司。
1.2 原核表达载体构建
将水稻叶绿体相关基因CPN60-β序列在NCBI网站进行同源性比对,以Primer5.0设计特异性引物CPN60-β-Forward:5′-TATAGGATCCACCGCA AAAGCACTG GTT G-3′(斜体加下划线字体为酶切位点BamHI,加粗字体为保护碱基)和CPN60-β-Reverse:5′ -TA TACTCGAGGCGCTCGTTCAGTTTCTC-3′(斜体加下划线字体为酶切位点XhoI,加粗字体为保护碱基)。用植物组织总RNA快速提取试剂盒提取水稻幼嫩叶片总RNA后,以反转录试剂盒进行RT-PCR。反应体 系50 μL:RNA模 板0.1 μg,RT-PCR MasterMix buffer 25 μL,RT-PCR Enzyme Mix buffer 2 μL,上下游引物各1.5 μL,ddH2O补足至50.0 μL。PCR反应条件:42 ℃ 30 min;95 ℃预变性3 min;94 ℃ 30 s、58 ℃ 40 s、70 ℃ 40 s,40个循环;70 ℃延伸10 min。扩增产物和表达载体pET-28a(+)都用BamH І和XhoI进行37 ℃双酶切过夜。酶切体系为:RT-PCR产物或者 表达载体质 粒2 μL,Buffer 1 μL,BamH І和XhoI各0.5 μL,ddH2O补足至10 μL。酶切产物回收后与T4 DNA连接酶于16 ℃连接过夜,连接体系为:表达载体质粒3 μL,RT-PCR酶切产物5 μL,T4 DNA连接酶1 μL,Buffer 0.5 μL,ddH2O补足至10 μL。连接后,将产物热击转化大肠感觉感受态细胞DH5α,涂布LB琼脂培养平板,然后挑取培养平板单克隆进行PCR和测序验证,正确的重组表达载体命名为pET-CPN60-β。
1.3 CPN60-β基因的原核表达与检测鉴定
将重组表达载体pET-CPN60-β热击转化大肠杆菌BL21感受态细胞并涂布于含卡那霉素(50 μg·mL−1)的LB固体培养基,过夜培养后挑取单克隆接种到LB液体培养基(含卡那霉素50 μg·mL−1)中,37 ℃、220 r·min−1振荡培养12 h。第2 d按照1∶100倍比稀释后置于37 ℃、220 r· min−1振荡培养至对数期(OD600约为0.6~0.8),添加诱导剂IPTG 0.5 mmol·L−1,在37 ℃和220 r· min−1振荡培养条件下诱导目的基因表达3.5 h。12 000 r·min−1离心沉淀细胞,超声波裂解细胞适当时间,SDS-PAGE凝胶电泳跑胶,将胶进行考马斯染色初步确定CPN60-β基因是否表达。pET-CPN60-β重组质粒带有6×His标签,如果CPN60-β基因成功表达,那么CPN60-β蛋白和6×His能够融合表达,所以可以用His抗体检测鉴定目的基因是否成功表达。不同浓度的IPTG诱导表达pET-CPN60-β重组载体后,收集菌体蛋白进行Western blotting检测目的蛋白是否成功表达。以抗6×His抗体为一抗、辣根过氧化物酶HRP标记抗体为二抗,进行Western blot试验,鉴定CPN60-β蛋白是否融合6×His标签,从而进一步鉴定CPN60-β基因是否表达。在SDS-PAGE和Western blot试验过程中均设置不加IPTG的对照组。
1.4 抗血清的制备、纯化、效价和浓度测定
纯化CPN60-β蛋白,将其和一定量的弗氏完全佐剂充分混匀和乳化,乳化后免疫5月龄的雄性新西兰大白兔;随后每隔15 d加强免疫4~5次。为了监测免疫过程中是否产生抗体,在蛋白免疫前和第2次加强免疫后第15 d分别抽血采样15 mL,将血液样品在4 ℃冰箱静止保存过夜。为了收集血样中的抗血清,将血样在8 000 r min−1离心20 min。随后,将抗血清按照1∶500、1∶2 000、1∶5 000、1∶10 000、1∶100 000和1∶1 000 000的稀释比例稀释,然后在室温条件下孵育一抗60 min,孵育二抗 50 min,显色30 min。
在第4次加强免疫后15 d,采集兔子全血60 mL,按上述方法获取血清。将蛋白Beads添加至纯化柱子后,收集好的抗血清添加至柱子内部,室温条件下或者4 ℃冰箱和Beads共孵育;孵育结束后用PBS缓冲液冲洗5次;然后用洗脱液洗脱收集抗血清蛋白。采用上述ELISA方法检测纯化后抗血清的效价。将CPN60-β融合蛋白免疫大白兔前所采的血设为阴性对照。为了避免动物死亡,设置2个平行试验,即免疫2只动物。
抗体结构中重链和轻链分子大小分别为55 kDa和25 kDa,因此,可通过还原性SDS-PAGE分离后考马斯显色观察抗体重莲含量来检测抗体的浓度。
2 结果与分析
2.1 叶绿体基因CPN60-β的扩增与表达载体构建
以水稻幼嫩叶片总RNA为模板、CPN60-β-Forward/CPN60-β-Reverse为引物,RT-PCR扩增水稻叶绿体基因CPN60-β。RT-PCR产物经琼脂糖凝胶电泳检测表明,目的基因片段CPN60-β的大小大约为657 bp(图1-A)。目的基因经过克隆转化后,任意选取LB固体培养基平板上的单克隆菌落,PCR鉴定阳性单菌落。经琼脂糖凝胶电泳检测,结果表明,成功重组的质粒pET-CPN60-β能够扩增出大小约为657 bp的目的基因CPN60-β片段(图1-B)。阳性重组质粒pET-CPN60-β送至广州生工基因股份有限公司测序,以验证序列正确与否。
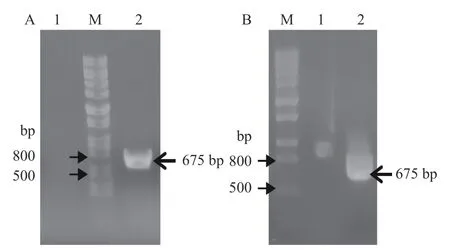
图1 重组质粒pET-CPN60-β构建Fig. 1 Construction of recombinant plasmid pET-CPN60-β
2.2 叶绿体基因CPN60-β的表达与鉴定
重组质粒pET-CPN60-β转化表达菌BL21后,为了明确CPN60-β基因是否在大肠杆菌BL21能够诱导表达,将诱导表达条件设置为:诱导剂IPTG浓度0.5 mmol·L−1、摇床温度37 ℃、转速220 r· min−1、振荡培养3.5 h进行诱导表达。SDS-PAGE电泳结果表明,在此诱导表达条件下,重组质粒表达约30 kDa的蛋白条带,而不添加IPTG的对照中并未出现特异蛋白条带(图2-A),该结果初步表明重组质粒pET-CPN60-β能够表达分子量约30 kDa的CPN60-β融合蛋白。为了进一步验证CPN60-β是否表达,以抗6×His抗体为一抗、辣根过氧化物酶HRP标记抗体为二抗进行Western blot试验,结果表明,在诱导表达组中,在30 kDa处6×His抗体能够特异结合其抗原6×His进而显色,而不添加IPTG的对照中在相应位置并未结合和显色(图2-B)。因为CPN60-β融合了6×His,所以该结果进一步表明CPN60-β融合蛋白能够成功诱导表达。
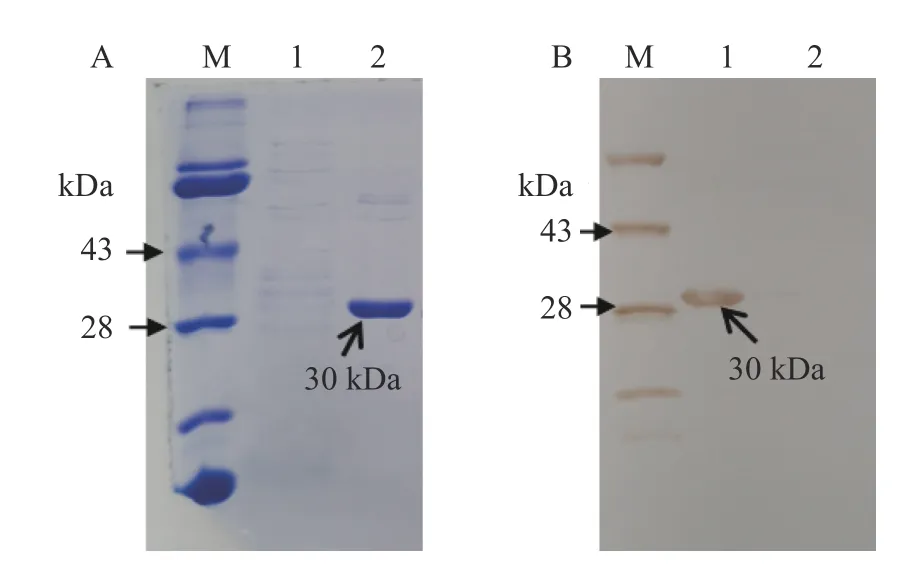
图2 CPN60-β基因的表达与鉴定Fig. 2 Expression and confirmation of CPN60-β gene
2.3 抗CPN60-β抗体的效价和浓度
为了监测免疫过程中是否产生抗CPN60-β多克隆抗体,在蛋白免疫前和第2次加强免疫后采集少量血样,收集血清,倍比稀释后采用ELISA检测免疫前和加强免疫后(未纯化)的抗血清效价。ELISA检测结果表明,与免疫前和空白对照相比,未纯化加强免疫后的抗血清随着稀释比例加大,ELISA显色越来越弱;稀释比例达到1∶1 000 000显色反应仍然可见(图3-A),表明未纯化的加强免疫后抗血清的效价为1∶1 000 000。
为了明确多克隆抗体纯化前后的效价,多克隆抗体经亲和配体纯化、倍比稀释后,以ELISA检测其抗体效价。ELISA检测结果显示,与纯化前的抗血清对照相比,在相同倍比稀释下,纯化后的抗体ELISA显色反应明显比较深,该结果说明抗体经过纯化后其浓度得到有效的提高(图3-B)。此外,多克隆抗体纯化前后在倍比稀释达到1∶1 000 000时,ELISA显色反应仍然较为明显,说明纯化后的抗体效价都达到1∶1 000 000(图3-B)。然而,洗脱液ELISA反应不显色,表明抗体大部分被纯化(图3-B)。
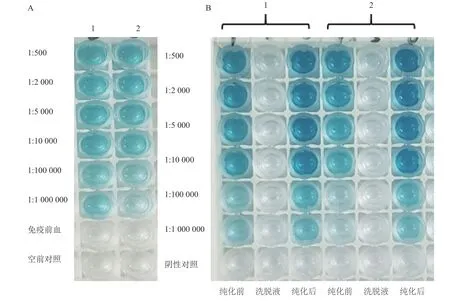
图3 抗CPN60-β抗体的效价测定Fig. 3 Titer of antibodies against CPN60-β
抗体结构中重链和轻链分子大小分别为55 kDa和25 kDa。因此,抗体通过变性或还原性SDS-PAGE分离后考马斯显色可观察抗体重链含量从而检测抗体的浓度。经SDS-PAGE电泳检测后抗体的含量约为300 μg·mL−1(图4)。
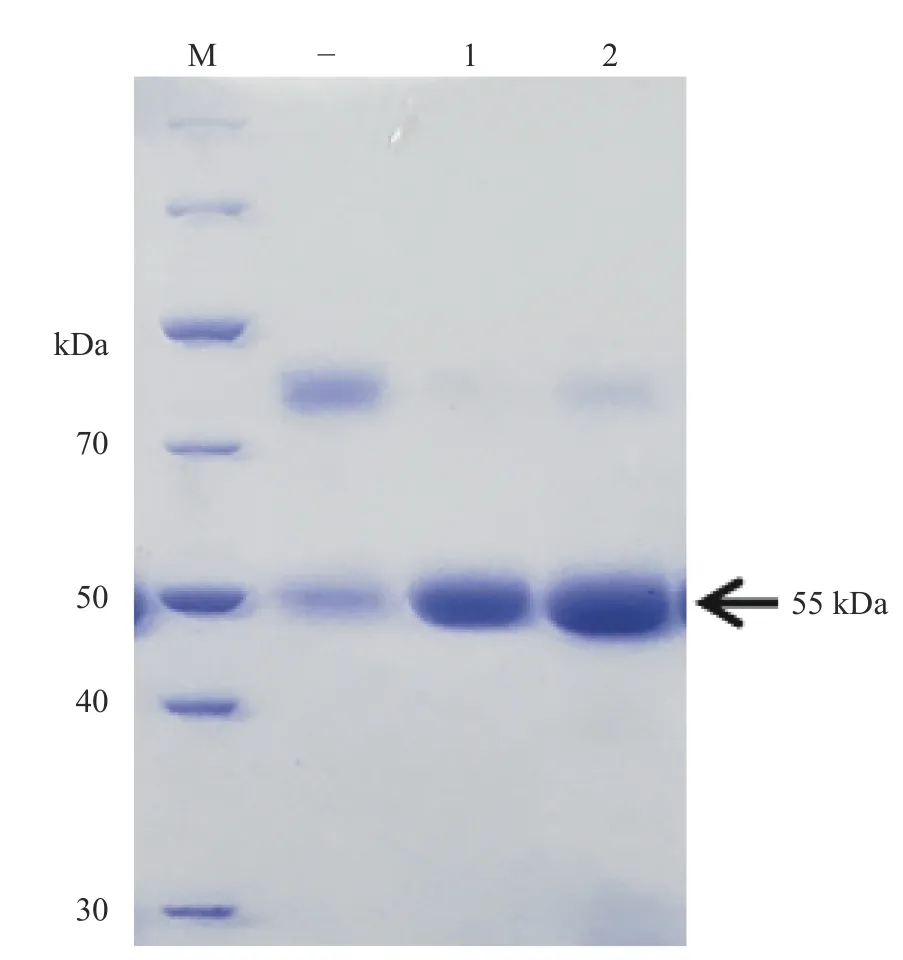
图4 变性SDS-PAGE检测抗CPN60-β抗体浓度Fig. 4 Antibody concentration estimated by denatured SDSPAGE
3 讨论
前人的研究结果表明,RSV编码的运动蛋白NSvc4在模式植物烟草中参与病毒的长距离、胞间运动,以及本氏烟感病症状的形成[8,13−14],证明NSvc4参与病毒的运动和致病等侵染过程。但是NSvc4的运动或致病机制仍未明确。因此,加深NSvc4与寄主水稻的互作机理的研究和了解,对于病毒侵染机制和病毒病害的综合绿色防治具有重要意义。病毒编码的蛋白数量有限,其必须招募或者利用寄主细胞内的蛋白来完成侵染过程。笔者在酵母双杂交试验中发现,NSvc4与水稻叶绿体相关的一个蛋白——RuBisCO亚基结合蛋白Chaperonin-60-beta(CPN60-β)能够特异互作。因此,猜测CPN60-β蛋白在NSvc4蛋白介导的RSV在寄主中的运动和致病过程中发挥重要作用。但是,目前还缺乏针对CPN60-β蛋白的特异抗体,阻碍了CPN60-β蛋白在细胞内定位、发挥作用机制等问题的研究。本试验制备的水稻CPN60-β多克隆抗体有助于研究CPN60-β蛋白在RSV病毒运动和致病过程中的功能。
目前,通过蛋白免疫新西兰大白兔制备多克隆抗体的方法具有成熟、简单,周期短等优点[15]。多克隆抗体能够制备成功的关键点为能否正确选取抗原蛋白的抗原片段[16]。本研究首先通过分析水稻CPN60-β蛋白分子的抗原指数、可及性、亲水性和疏水性等特性,选取抗原指数较高的179~422 aa区段作为抗原。然后扩增目的基因CPN60-β,且成功构建原核表达载体pET-CPN60-β并进行高效表达,获得的CPN60-β融合蛋白经Ni2+-NTA琼脂糖亲和层析纯化后作为抗原免疫雄性新西兰大白兔,成功制备获得抗水稻CPN60-β多克隆抗体,并证实制备获得的CPN60-β多克隆抗体纯度较高,其质量浓度和效价分别为300 μg mL−1和1∶1 000 000。CPN60-β抗体的制备为后续进一步研究CPN60-β在NSvc4介导的RSV运动和致病过程中发挥的具体机制提供了重要材料。稻谷等粮食是人类赖以生存和发展的重要物质基础。病毒病一直严重影响着我国水稻等粮食生产和粮食安全。粮食安全是关乎国计民生的头等大事,我国农业病虫害多发,严重威胁农业生产安全和农产品质量。CPN60-β抗体的制备将有助于加深对RSV NSvc4及其CPN60-β在病毒致病过程中的具体作用机制的理解,并可望通过设计靶位点开发出针对水稻病毒病的绿色防控技术。
