Resistance genes mediate differential resistance to pine defensive substances α-Pinene and H2 O2 in Bursaphelenchus xylophilus with different levels of virulence
2021-07-15LinRuiHongbinLiuRuiLiangXiaoqinWu
Lin Rui·Hongbin Liu·Rui Liang·Xiaoqin Wu
Abstract The pine wood nematode (PWN),Bursaphelenchus xylophilus (Steiner &Buhrer) Nickle,is the pathogen of pine wilt disease (PWD) which can devastate forests.PWN can be of high or low severity and the mechanisms underlying the differences in virulence are unclear.Therefore,it is necessary to study the relationship between differentiation of PWN severity and its resistance to the main defensive substances of pine species (i.e.,α-pinene and H2 O2).The feeding rate and fecundity of PWN was examined at different levels of virulence under conditions of α-pinene and H2 O2 stress.Moreover,the expression patterns of the main resistance genes of PWN with different virulence were determined under conditions of α-pinene and H2 O2 stress.The feeding rate and fecundity of the high virulence strain AMA3 were higher than those of the low virulence strain YW4.The expression levels of the autophagy gene BxATG5,cytochrome P450 gene BxCYP33D3,and glutathione S-transferase genes BxGST1 and BxGST3 in AMA3 increased significantly upon exposure to α-pinene for 2 h,while these genes showed smaller degrees of upregulation in YW4.Under conditions of H2 O2 stress,the expression levels of BxATG5,catalase genes Bxy-ctl-1 and Bxy-ctl-2,and the 2-cysteine peroxiredoxin gene BxPrx in AMA3 were higher than those in YW4.These findings suggest that high virulence PWN has greater resistance to pine defensive substances α-pinene and H2 O2 than low virulence PWN,and resistance genes mediate the differential resistance of PWN strains.This study will contribute to the clarification of the mechanism underlying virulence differentiation of PWN and will advance understanding of the pathogenic mechanism of PWD.
Keywords Pine wood nematode·Virulence·Defensive substances·Autophagy gene·Detoxification gene·Antioxidant gene
Introduction
Pine wilt disease (PWD) is caused by the well-known quarantine pest,the pine wood nematode (PWN),Bursaphelenchus xylophilus(Steiner &Buhrer) Nickle,which belongs to the family Aphelenchoididae,and has a devastating effect onPinusspecies (Zhou et al.2017).PWN infects 58 species ofPinusand 13 other coniferous species (He et al.2014).The disease originated in North America and is found in Canada,the United States,and Mexico.It also occurs in Spain and Portugal in Europe,and in China,Japan,South and North Korea in Asia (He et al.2014;Ye 2019).PWD was first discovered in 1982 at the Sun Yat-sen Mausoleum in Nanjing,China,and has since spread rapidly (Cheng et al.1983).China is currently the country most threatened by pine wilt disease (Huang et al.2019;Ye 2019).However,the pathogenic mechanism of PWD is not well understood and it remains difficult to prevent and control (Li et al.2018).
The life cycle of the disease involves the dispersal phase and the propagative phase (Futai 2013).When encountering adverse environments such as food scarcity or low temperatures,the second stage propagative larvae molt and develop into third-stage dispersal larvae,and then grow into a specific dispersal stage or alternate developmental stage,of dauer larvae (Ogura and Nakajima 2002;Kikuchi et al.2011).Following dispersal from dead to healthy pines viaMonochamusvector beetles (Zhao et al.2016;Wu et al.2019),the dauer larvae infect the pine host to feed (Vicente et al.2013).In the infection process,the pines secrete various defensive substances to attack the nematodes.Pine resin consisting of turpentine (monoterpenes and sesquiterpenes)and rosin (diterpenes) is the first line of defense (Zulak and Bohlmann 2010).Monoterpenes,the most essential defensive volatile substances,can help plants resist relevant pathogen and herbivore invasions (Lewinsohn et al.1993).The monoterpenes secreted by conifers are mainly α-Pinene(Tiao et al.2006).Li et al.(2019) reported that the reproduction of PWN was dramatically inhibited by 56.3 mg mL ‒1of α-pinene.In addition,plants can also produce reactive oxygen species (ROS) to damage organelles and inhibit cell functions in pathogens (Baker and Orlandi 1995).The most abundant and stable ROS in pines is hydrogen peroxide,H2O2(Vellosillo et al.2010).The resin and ROS levels in pine trees increase markedly during the early stages of the disease in response to invasion by the nematodes (Yang et al.2003;He et al.2010;Yu et al.2013).On the other hand,resistance genes protect the nematodes from damage by pine defensive substances.Autophagy is a subcellular degradation pathway which helps to degrade and cycle aging and damaged organelles and proteins in eukaryotes (Ktistakis et al.2016).In addition,the autophagy geneBxATG5shows strong affiliations with the growth and development,stress resistance,and virulence of PWN (Liu et al.2019a;2020).The degradation of pine defensive substances by the pine wood nematodes is vital for successful infection.The cytochrome P450 geneBxCYP33D3is responsible for the detoxification of α-pinene (Xu et al.2015).PWN responds to the accumulation of α-pinene by upregulating cytochrome P450 gene expression (Wang et al.2017).Glutathione S-transferase (GST) is an important detoxifying enzyme in eukaryotes.The GST genes,BxGST1andBxGST3,are closely associated with feeding rate,reproduction,and pathogenicity of the nematodes (Nian 2017).The catalase(CAT) genes,Bxy-ctl-1andBxy-ctl-2,and the 2-cysteine peroxiredoxin geneBxPrxwere reported to be the main antioxidant genes in PWN,which can convert H2O2to water by regulating the activities of CAT and peroxiredoxin (Li et al.2011;Vicente et al.2015;Xu et al.2019).
PWN can have high or low virulence (Shinya et al.2012;Ding et al.2016),and its severity and adaptability to unfavorable environments are strongly correlated with the spread and development of pine wilt disease (Liu et al.2019b).The degree of virulence of PWN is positively correlated with its reproductive ability in pines and onBotrytis cinereaPers.or grey mould (Wang et al.2005;Aikawa and Kikuchi 2007).Eo et al.(2011) reported that the virulence of pine wood nematodes was not correlated with its migration rate.However,the mechanism underlying differences in PWN virulence are unknown.Therefore,in this study,we evaluated the effectiveness of the pine defensive substances α-pinene and H2O2on the feeding,reproduction,and resistance gene expression patterns of PWN at different levels of virulence to explore the relationship between nematode resistance and its virulence differentiation.The results of this study should further our understanding of the mechanism underlying the virulence differentiation of pine wood nematodes and the sophisticated pathogenesis of pine wilt disease.
Materials and methods
Culturing of PWN
The nematode strain AMA3 that exhibited high virulence was isolated from diseasedPinus thunbergiiParl.from Anhui,China;the strain YW4 that showed low virulence was isolated from diseasedPinus kesiyavar.langbianensis(A.Chev.) Gaussen ex Bui from Yunnan,China.Both strains were supplied by the Forest Protection Laboratory,Nanjing Forestry University.Experimental nematodes were cultivated on mats ofB.cinereagrown on potato dextrose agar (PDA) plates of 7 g of fresh potatoes,0.7 g of dextrose,0.7 g of agar,and 30 mL of sterile water at 25 °C.When the hypha was totally consumed,nematodes at various stages were extracted by a Baermann funnel (Viglierchio and Schmitt 1983) and washed three times with M9 buffer (30 g L‒1KH2PO4,60 g L‒1K2HPO4,and 50 g L‒1NaCl).
Assay of feeding rate and reproduction of PWN under H2 O2 and α-pinene stress
Five hundred samples of AMA3 and YW4 treated with double distilled water (ddH2O),0.5% Triton X-100,56.3 mg mL‒1α-pinene,or 15 mM H2O2,were inoculated on fungal mats ofB.cinereaand incubated at 25 °C in three replicates and photographed periodically.Subsequently,the nematodes were isolated from the plates by the Baermann funnel method and counted under a stereomicroscope (Leica DM500;Leica Microsystems,Heerbrugg,Switzerland)when at least one of the treatments had completely consumed the hypha.
Collection of PWN treated with α-pinene and H2 O2
Organisms usually respond quickly by regulating defense genes after being stressed (Tsyglakova et al.2019).Therefore hourly treatments time up to five hours were selected.Samples of 7000 AMA3 and 7000 YW4 were soaked in 1 mL of 56.33 mg mL‒1α-pinene or 15 mM H2O2hourly up to five hours in three replicates.The nematodes were washed and stored at ‒80 °C after rapid freezing by liquid nitrogen.
RNA extraction and cDNA synthesis
Samples were ground into powder by a high-speed tissue grinder and TRIzol reagent (Invitrogen,Carlsbad,CA,USA)added.After shaking and mixing,the samples stood for 5 min,then centrifuged (12,000 rpm),and the supernatants transferred to new tubes.After extraction with chloroform and isopropanol,the samples were washed with 75% ethanol and dried.Total RNA was extracted and dissolved in a volume of 20 μL diethyl pyrocarbonate-treated water at 50 °C for 2 min.The purity of total RNA was electrophoresed and measured on a Nanodrop 2000C spectrophotometer (Thermo Fisher Scientific,Waltham,MA,USA).cDNA was synthesized from 1 μg of total RNA according to HiScript® II Q RT SuperMix for qPCR (+gDNA wiper;Vazyme,Nanjing,China).
Quantitative reverse transcription-polymerase chain reaction
The qRT-PCR primers forBxATG5(MK387710) andBxCYP33D3(KM973212) were designed according to their coding sequences (https ://www.oligo archi tect.com);the products were 80–150 bp.Primers forBxGST1,BxGST3,Bxy-ctl-1,Bxy-ctl-2,andBxPrxwere from previous studies(Vicente et al.2015;Li et al.2016;Nian 2017).TheActingene (EU100952) of the pine woody nematode served as an internal control,with the primers listed in Table 1 (Hu et al.2020).

Table 1 PCR primers used in the study
SYBR Green Master Mix (Vazyme) and a two-step method were used for qRT-PCR;the conditions were as follows:95 °C for 5 min for initial denaturation,followed by 40 cycles of 95 °C for 10 s and 60 °C for 34 s.Melting curve analysis was performed using the default program:95 °C for 15 s,60 °C for 1 min,and 95 °C for 15 s.Relative expression levels were calculated using the 2 −ΔΔCt method of ABI Prism 7500 software (Applied Biosystems,Foster City,CA,USA)(Livak and Schmittgen 2001).qRT-PCR was conducted using three biological and technical replicates.
Statistical analysis
All assays had three biological replicates and three technical replicates.Microsoft Excel software (Microsoft Corp.,Redmond,WA,USA) was used for the calculation of mean and standard deviation of the three biological replicates.SPSS software (ver.17.0;IBM China Company Ltd.,Beijing,China) calculated statistical significance by conducting the paired Student’st-test or Tukey’s HSD test.In all analyses,P< 0.05 indicated statistical significance.
Results
Feeding rate of High and low virulence PWN under conditions of α-pinene and H2 O2 stress
The feeding rates of high virulence AMA3 and low virulence YW4 were first evaluated in the absence of stress.The dark area of the culture medium indicated that the hypha had been consumed.And the results show that both AMA3 and YW4 cultured onB.cinereaconsumed all hyphae by day 5 (Fig.1 a).AMA3 and YW4 were next treated with Triton X-100,α-pinene,and H2O2to evaluate their feeding rates.The results show that feeding rates did not differ between AMA3 and YW4 treated with Triton X-100 as a negative control for α-pinene.Both AMA3 and YW4 treated with α-pinene fed more slowly than those treated with Triton X-100,and α-pinene had a stronger inhibitory effect on YW4 than on AMA3.Under conditions of α-pinene stress,AMA3 began to feed on day 6,while YW4 had not begun.On day 7,AMA3 had almostfinished feeding whereas YW4 had just begun (Fig.1 b).Under conditions of H2O2stress,AMA3 had consumed a small portion of hyphae on day 5,while YW4 had not begun to feed.By day 6,AMA3 had finished feeding,while YW4 had consumed one quarter of the hyphae(Fig.1 c).Therefore,high virulence pine woody nematodes fed more rapidly than low virulence PWN under conditions of α-pinene and H2O2stress.

Fig.1 Feeding rates of pine wood nematodes with different levels of virulence inoculated onto Botrytis cinerea with different treatments;dark areas of the culture medium indicates that the hypha has been consumed.a ddH2O .b α-pinene.Triton X-100 served as negative control.c H2O2
Fecundity of the high virulence PWN compared with the low virulence PWN under conditions of α-pinene and H2 O2 stress
Levels of reproduction were similar for high virulence AMA3 and low virulence YW4 in the absence of stress(Fig.2 a).AMA3 and YW4 were then treated with Triton X-100,α-pinene,or H2O2.The results show that there was no difference in reproduction between AMA3 and YW4 treated with Triton X-100 as a negative control for α-pinene.The α-Pinene had no effect on the reproduction of AMA3 but markedly reduced the reproduction of YW4.The reproduction of AMA3 was more than three times higher than YW4 under conditions of α-pinene stress (Fig.2 b).Under H2O2stress,the reproduction of AMA3 was normal while that of YW4 was inhibited.The reproduction of AMA3 was almost two-fold that of YW4 under conditions of H2O2stress(Fig.2 c).Therefore,the fecundity of high virulence PWN was greater than that of low virulence nematodes under conditions of α-pinene and H2O2stress.
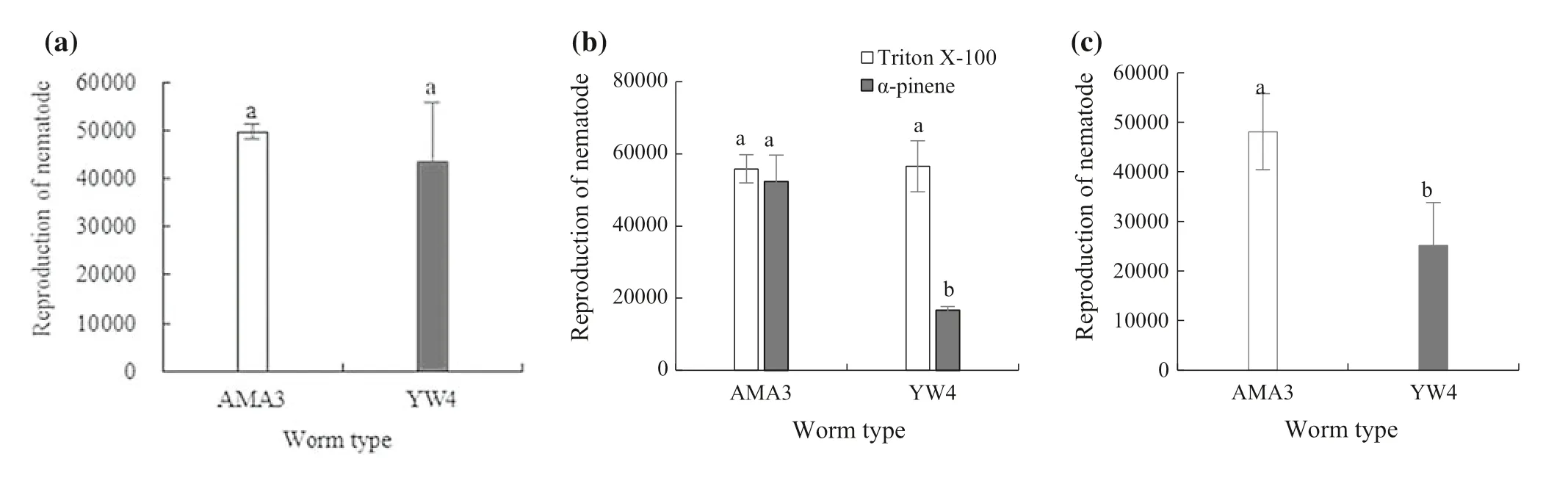
Fig.2 Reproduction of pine wood nematodes with different levels of virulence;AMA3 and YW4 treated with ddH2 O,α-pinene,Triton X-100,and H2 O2 .a ddH2 O.b α-pinene.Triton X-100 served as negative control.c H2 O2 .Error bars indicate SE (n=3);letters indicate significant differences at the 5% level by paired Student’s t-test or Tukey’s HSD test
Expression levels of BxATG5 in PWN with different virulence levels under conditions of α-pinene and H2 O2 stress
The autophagy geneBxATG5was reported to be related to the stress resistance of pine wood nematodes (Liu et al.2019a).The expression ofBxATG5was examined in high virulence AMA3 and low virulence YW4 exposed to α-pinene and H2O2hourly up to 5 h.TheActingene(EU100952) of PWN served as an internal control.Under conditions of α-pinene stress,the expression level ofBxATG5in AMA3 showed two peaks at 2 and 4 h,with the first peak being much higher than the second.The expression level ofBxATG5in YW4 peaked at 4 h and was lower than the first peak of AMA3.The expression ofBxATG5in AMA3 was significantly upregulated more than five-fold by exposure to α-pinene for 2 h,but in YW4 it was downregulated almost 0.7-fold (Fig.3 a).These results indicate that the expression variation response to α-pinene ofBxATG5in AMA3 was greater than in YW4.Under conditions of H2O2stress,the expression ofBxATG5showed significant upregulation at 2 h and remained on high level in AMA3.However,the expression ofBxATG5in YW4 increased slightly at 2 h,decreased immediately at 3 hand slowly returned to normal.AMA3 showed upregulated expression ofBxATG5by almost four-fold upon exposure to H2O2for 2 h,while the expression ofBxATG5in YW4 showed less than two-fold increase at the same time point.Moreover,AMA3 showed a higher level of expression ofBxATG5at all time periods compared to YW4 (Fig.3 b).Therefore,the response ofBxATG5to H2O2stress was stronger and of longer duration in high virulence PWN.
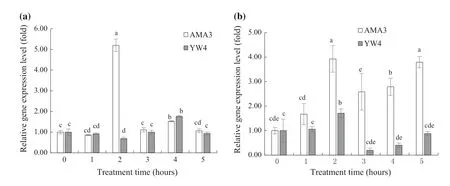
Fig.3 Expression patterns of the autophagy gene BxATG5 with different levels of virulence under conditions of α-pinene and H2 O2 stress;a α-pinene,b H2 O2; Error bars indicate SE (n =3),letters indicate significant differences at the 5% level by Tukey’s HSD test
Detoxification gene response to α-pinene in high virulence and low virulence PWN
We examined the expression patterns of the cytochrome P450 geneBxCYP33D3and the GST genesBxGST1andBxGST3in high virulence AMA3 and low virulence YW4 exposed to α-pinene by qRT-PCR.Under conditions of α-pinene stress,the expression level ofBxCYP33D3in AMA3 peaked at 2 h and then decreased to the normal level by 5 h.In YW4,a smaller peak was observed at 2 h and then returned to normal by 3 h.AMA3 showed rapid upregulation ofBxGST1expression to a peak at 2 h followed by a decrease to a low level,while the expression ofBxGST1in YW4 remained normal throughout.There was a surge in the expression level ofBxGST3in AMA3 at 2 h,and the expression remained normal at other time points.The expression ofBxGST3in YW4 increased slightly up to 4 h.In AMA3,the expression levels of the detoxification genesBxCYP33D3,BxGST1,andBxGST3peaked upon exposure to α-pinene for 2 h,showing 1.5-,2.8-,and 12.7-fold increases,respectively,whereas no such upregulation was detected in YW4(Fig.4).These results indicate that AMA3 is more efficient at upregulating detoxification genes to reduce damage by α-pinene in comparison with YW4.Therefore,the virulence of PWN was dependent on its ability to upregulate genes related to the detoxification of α-pinene.

Fig.4 Expression patterns of the cytochrome P450 gene BxCYP33D3 and glutathione S-transferases genes BxGST1 and BxGST3 in pine wood nematodes with different levels of virulence under conditions of α-pinene stress;a BxCYP33D3,b BxGST1,c BxGST3;error bars indicate SE (n=3),letters indicate significant differences at the 5% level by Tukey’s HSD test
Effects of H2 O2 on the expression of antioxidant genes in high virulence and low virulence PWN
The expression of the CAT genesBxy-ctl-1andBxy-ctl-2and the peroxidase geneBxPrxin high virulence AMA3 and low virulence YW4 exposed to H2O2by qRT-PCR were examined.Bxy-ctl-1expression patterns were similar in AMA3 and YW4,increasing slowly to peak with H2O2treatment for 4 h followed by a decrease by 5 h under conditions of H2O2stress.However,the overall expression level ofBxy-ctl-1in AMA3 was higher than in YW4;the peak ofBxy-ctl-1in AMA3 was 1.4-fold higher than in YW4(Fig.5 a).The expression ofBxy-ctl-2in AMA3 increased significantly after exposure to H2O2for 1 h,returned to normal by 4 h,and increased again at 5 h.YW4 showed downregulation ofBxy-ctl-2expression at 4 h and upregulation at 5 h under conditions of H2O2stress.Importantly,AMA3 showed a higher expression level ofBxy-ctl-2than YW4 throughout the period of hydrogen peroxide treatment (Fig.5 b).The trend of expression byBxPrxin AMA3 was consistent with that in YW4 under conditions of H2O2stress,showing a gradual increased to a peak,followed by a decrease,and then increasing again.Importantly,AMA3 also showed a higher expression level ofBxPrxthan YW4 during the period of H2O2treatment.(Fig.5 c),according to the expression of antioxidant genes,suggesting that the antioxidant capacity of high virulence PWN is stronger than that of low virulence PWN.

Fig.5 Expression patterns of the catalase genes Bxy-ctl-1 and Bxyctl-2 and peroxidase gene BxPrx of pine wood nematodes with different levels of virulence under conditions of H2 O2 stress;a Bxy-ctl-1,b Bxy-ctl-2,c BxPrx;error bars indicate SE (n=3);letters indicate significant differences at the 5% level by Tukey’s HSD test
Discussion
Hydrogen peroxide and α-Pinene are closely related to the resistance of pine species to pine wood nematodes.Therefore,the differences in virulence of PWN are likely related to the responses to these defense molecules.Tan et al.(2009)reported that α-pinene had no effect on the fertility of PWN.However,Li et al.(2019) noted that the reproduction of the nematodes was severely inhibited by 56.3 mg mL ‒1of α-pinene.In this study,we examined the impact of α-pinene and H2O2on the feeding rate and fecundity of PWN at different levels of virulence and found that both compounds markedly reduced feeding rates and reproduction of YW4,but not of AMA3.These results show that the differential resistance of the nematodes to pine defensive substances is a key factor in their pathogenic differentiation.
The autophagy geneATG5is a central factor involved in autophagosome assembly inSaccharomyces cerevisiaeDesm.,and its protein product has an important role in ubiquitin-like modifications in the process of autophagy (Matsushita et al.2007).Miller et al.(2008) noted thatATG5has major roles in B cell survival during development.Previous studies showed that α-pinene and ROS production in pine species were induced after infection with PWN (Liu et al.2008).H2O2and α-Pinene induced significant upregulation ofBxATG5expression in PWN (Liu et al.2019a).We treated the nematodes with different virulence levels with α-pinene and H2O2in vitro.The expression ofBxATG5in high virulence PWN peaked at 2 h,while in low virulence PWN decreased under conditions of α-pinene stress.High virulence PWN retained a higher expression level ofBxATG5than low virulence PWN when stressed with H2O2.Moreover,autophagy assists eukaryotes to degrade aberrant cytoplasmic components for cellular homeostasis (Boya et al.2013;Liu et al.2019c).High virulence pine wood nematodes showed significant upregulation ofBxATG5expression earlier than low virulence PWN under conditions of α-pinene and H2O2stress.Therefore,high virulence PWN may have a greater ability to remove the deleterious effects and recycle α-pinene and H2O2by activating autophagy genes earlier.
The cytochrome P450s of insects can help detoxify pesticides (Yue et al.2007;Xu et al.2009).The Ce-GST-p24 of the nematode,Caenorhabditis elegansMaupas was closely related to xenobiotic detoxification (Leiers et al.2003).Tsai et al.(2016) reported transcriptomic differences in PWN fromB.cinereaand pine species,and Li et al.(2019) conducted a comparative transcriptome analysis of PWN treated with sterile water and α-pinene.Both demonstrated upregulated expression of cytochrome P450 and GST family genes.These genes of PWN show significant roles in responding to α-pinene stress.Moreover,the expression of cytochrome P450 genes,related to the vitality,transmissibility,reproduction,pathogenicity,and metabolism of toxic substances of PWN,were shown to be involved in the response of PWN to the accumulation of α-pinene in pine species (Xu et al.2015;Wang et al.2017).Silencing ofBxGST1andBxGST3inhibited the feeding,reproduction,and pathogenicity of PWN (Nian 2017).We further found that high virulence pine wood nematodes showed rapid upregulation ofBxCYP33D3,BxGST1,andBxGST3expression upon exposure to α-pinene,while low virulence PWN responded considerably more slowly,suggesting that differential responses to α-pinene in PWN with different virulence are mediated by detoxification genes.
CATs,which are widespread among Animalia,Plantae,and Fungi,are able to reduce H2O2to water and to oxidize it to molecular oxygen (Zamocky et al.2008).The regulation ofCATgene expression shows significant role in determining the levels of CAT proteins (Kodydkova et al.2014).In pengze crucian carp (Carassius auratusL.),the content of active oxygen in the liver increases on stimulation with cadmium ions and the expression ofCATis also upregulated,indicating thatCATshows significant role in the antioxidant defense system (Li et al.2013).InArabidopsis thaliana(L.)Heynh.,CAT1helps to remove hydrogen peroxide from environmental stresses.CAT2andCAT3separately remove H2O2under light and dark conditions to maintain the organism’s balance (Du et al.2008).The CAT protein family of the fungal entomopathogenBeauveria bassiana(Bals.-Criv.)Vuill.consists of CAT-A (spore-specific),CAT-B (secreted),CAT-P (peroxisomal),CAT-C (cytoplasmic),and CAT-D(secreted peroxidase/catalase).These five CAT proteins function redundantly in regulating the tolerance of fungi to hyperosmotic or fungicil stress (Wang et al.2013).Kim et al.(1989) discovered that oxidative stress can activate the expression of Peroxiredoxin inS.cerevisiaeDesm..InHaemonchus contortusRudolphi,HcPrx2 can protect the nematode against oxidative damage (Hudson et al.2011).Previous studies cloned theCATgenesBxy-ctl-1andBxyctl-2and the peroxidase geneBxPrxfrom PWN,which are related to the elimination of H2O2produced by the host tree(Li et al.2011;Vicente et al.2015).We found upregulation ofBxy-ctl-1,Bxy-ctl-2,andBxPrxexpression in PWN within 5 h of H2O2stress,and the magnitude of upregulation was greater with high virulence than with low virulence PWN.Vicente et al.(2015) also found that the changes in expression levels ofBxy-ctl-1andBxy-ctl-2in response to different concentrations of H2O2were greater in high virulence than low virulence pine wood nematodes.Therefore,the antioxidant activity of high virulence PWN was stronger than that of low virulence PWN,which may have a significant role in the virulence differentiation mechanism of PWN.The resistance mechanisms of nematodes with different degrees of virulence should be investigated further to better understand virulence differentiation in PWN.
Conclusions
Feeding rate and fecundity of high virulence pine wood nematodes were significantly higher than for low virulence nematodes under conditions of α-pinene and H2O2stress,suggesting that high virulence pine wood nematodes have greater resistance than low virulence PWN.Moreover,the expression levels of the main resistance genes,including autophagy genes,detoxification genes,and antioxidant genes,in high virulence pine wood nematodes were all greater than in low virulence PWN under conditions of α-pinene and H2O2stress.These results suggest that the differential resistance of PWN with different degrees of virulence to α-pinene and H2O2is mediated by nematode resistance genes.This study will increase our understanding of the mechanism of virulence differentiation in PWN,and provides insight into the pathogenic mechanism of pine wood nematodes.
Author contributionsLin Rui designed the study,conducted experiments,analyzed the data and prepared the manuscript;Hong-Bin Liu designed the study and revised the manuscript;Rui Liang conducted experiments;Xiao-Qin Wu designed the study,acquired the data,contributed reagents/materials/analysis tools and approved the final manuscript.Open AccessThis article is licensed under a Creative Commons Attribution 4.0 International License,which permits use,sharing,adaptation,distribution and reproduction in any medium or format,as long as you give appropriate credit to the original author(s) and the source,provide a link to the Creative Commons licence,and indicate if changes were made.The images or other third party material in this article are included in the article’s Creative Commons licence,unless indicated otherwise in a credit line to the material.If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use,you will need to obtain permission directly from the copyright holder.To view a copy of this licence,visit http://creat iveco mmons.org/licen ses/by/4.0/.
杂志排行

Journal of Forestry Research的其它文章
- Flexible transparent wood enabled by epoxy resin and ethylene glycol diglycidyl ether
- Diversity and surge in abundance of native parasitoid communities prior to the onset of Torymus sinensis on the Asian chestnut gall wasp (Dryocosmus kuriphilus) in Slovenia,Croatia and Hungary
- Ozone disrupts the communication between plants and insects in urban and suburban areas:an updated insight on plant volatiles
- Testing visible ozone injury within aLight Exposed Sampling Site as aproxy for ozone risk assessment for European forests
- Logging and topographic effects on tree community structure and habitat associations in a tropical upland evergreen forest,Ghana
- Spatial pattern dynamics among co-dominant populations in early secondary forests in Southwest China