Morphological and kinetic parameters of the absorption of nitrogen forms for selection of Eucalyptus clones
2021-07-15MatheusSeverodeSouzaKulmannBetaniaVahldePaulaPaulaBeatrizSeteWagnerSquizaniArrudaGabrielAlbertoSansCamilaPeligrinottiTaroucoLucianeAlmariTabaldiFernandoTeixeiraNicolosoGustavoBrunetto
Matheus Severo de Souza Kulmann·Betania Vahl de Paula·Paula Beatriz Sete·Wagner Squizani Arruda·Gabriel Alberto Sans·Camila Peligrinotti Tarouco·Luciane Almari Tabaldi·Fernando Teixeira Nicoloso·Gustavo Brunetto
Abstract Eucalyptus clones are selected according to productivity,wood quality,rooting capacity,and resistance to drought,frost and diseases.However,kinetic and morphological parameters that determine the absorption efficiency of nutrients such as nitrate (NO3−) and ammonium (NH4+)are often not considered in breeding programs.The objective of this study was to evaluate the morphological,physiological and kinetic parameters of nitrogen uptake by clones of Eucalyptus saligna (32,864) and Eucalyptus grandis (GPC 23).Morphological parameters in shoot and root systems,biomass and N concentrations in different organs,photosynthetic pigment concentrations,parameters of chlorophyll a fluorescence and photosynthetic rates were evaluated.Kinetic parameters,maximum absorption velocity (V max),Michaelis–Menten constant (K m ),minimum concentration(C min ) and influx (I) were calculated for NO3− and NH4+ in the two clones.E.grandis clone was more efficient in the uptake of NO3− and NH4+,and showed lower K m and C min values,allowing for the absorption of nitrogen at low concentrations due to the high affinity of the absorption sites of clone roots to NO3− and NH4+ .Higher root lengths,area and volume helped the E.grandis clone in absorption efficiency and consequently,resulted in higher root and shoot biomass.The E.saligna clone had higher K m and C min for NO3− and NH4+,indicating adaptation to environments with higher N availability.The results of NO3− and NH4+ kinetic parameters indicate that they can be used in Eucalyptus clone selection and breeding programs as they can predict the ability of clones to absorb NO3− and NH4+ at different concentrations.
Keywords Ammonium and nitrate·Eucalyptus saligna·Eucalyptus grandis·Root system architecture·Nitrogen influx·Maximum absorption velocity (V max) ,Michaelis–Menten constant (K m) and Minimum concentration (C min)
Introduction
The global area of plantations continues to increase rapidly to address consumption demands for forest products.However,plantations provided only 39% of the world’s wood requirements in 2015 (FAO 2016),therefore highlighting the opportunity for plantations to satisfy current and future wood demands (Paquette and Messier 2010).Forest plantations occupy approximately 290.4 million ha worldwide(FAO 2016),of which 20 million ha areEucalyptus(Booth 2013).Among the eucalypt species used for reforestation,Eucalyptus grandisW.Hill andEucalyptus salignaSm.are economically important because they are tolerant to cold temperatures and mild frosts,as well as being fast growing and having high quality wood (Gonçalves et al.2013).
Eucalyptusclone plantations are commonly established on sandy-textured soils (Iglesias and Wilstermann 2008)with low organic matter and low levels of natural nitrogen(N).However,Eucalyptusclones used in commercial plantations are selected for their productivity,desirable wood qualities,rooting ability,and resistance to drought,cold,frost and diseases (Gonçalves et al.2013).However,kinetic parameters related to nutrient uptake efficiency such as nitrogen in the form of nitrate (NO3−) and ammonium (NH4+)are not generally considered,although N is a major nutrient that affects the growth and development ofEucalyptus.Nitrogen is a primary constituent of important plant components such as proteins,nucleic acids,adenosine 5-triphosphate (ATP),nicotinamide adenine dinucleotide (NADH),nicotinamide adenine dinucleotide phosphate (NADPH),chlorophyll,enzymatic cofactors,phytohormones,and secondary metabolites (Marschner 2012;Tomasi et al.2015).Based on the importance of nitrogen,Eucalyptusclones with higher N uptake efficiencies will be important contributors to breeding programs and will impact positively on higher wood yields.
N uptake by plants is generally in the mineral forms NO3−and NH4+,mediated by transport proteins or transporters in the plasma membranes of the epidermis and root cortex cells (Marschner 2012).The functioning of the transporters varies according to the affinity for NO3−and NH4+,and can be classified as high affinity (HATS),low affinity (LATS) or double affinity.In general,HATS proteins are activated at low concentrations as ions in solution(< 0.5 mmol L−1),while LATS act at higher concentrations(> 0.5 mmol L−1).The molecular basis of these absorption systems has been described forArabidopsis(Dechorgnat et al.2010) and identified in forest species (Kronzucker et al.1995) and fruit species (Pii et al.2014;Tomasi et al.2015).Based on this,NO3−-LATS and NO3−-HATS were encoded in two gene families,NRT1 and NRT2,respectively,except for NRT1.1,a dual affinity transporter (Pii et al.2014 ;Tomasi et al.2015),and for NH4+-LATSs and NH4+-HATS,belonging to the subfamily AMT2 and AMT1,respectively (Couturier et al.2007).Therefore,it is expected that plants will adapt to conditions of low nutrient availability in order to trigger high affinity systems,especially with nitrogen (Castro-Rodríguez et al.2017;Xuan et al.2017).
Kinetic parameters of nutrient absorption are the maximum absorption rate (V max),the minimum concentration(C min),the Michaelis–Menten constant (K m),and the influx(I) (Yang et al.2007;Martinez et al.2015).Vmax is the saturation point of root cell membrane transport sites by absorbed ions;C minis the minimum nutrient concentration in the solution for roots to initiate absorption;and,K mis a parameter that describes the affinity of the ions to the transporter system.A smallerKmdemonstrates greater ion affinity with the transport sites.Iis the inflow or velocity of ion absorption in a concentration solution (Martinez et al.2015;Alves et al.2016).
Nitrogen absorption by plants is important for many physiological processes,especially in the biosynthesis of essential proteins and enzymes involved in photosynthesis such as the enzyme Rubisco which results in higher CO2assimilation and contributes to the efficient use of water and nutrients (Tcherkez et al.2017;Nadal and Flexas 2018).Under high light conditions,the enzyme responsible for reducing NO3−in the assimilation process is activated,stimulating absorption (Marschner 2012).This occurs by the acquisition of light signals through the leaf and their transmission to other organs to contribute to the development of the root system and increases the absorption of water and nutrients such as nitrogen (Lee et al.2016).Light signals received by the shoots also regulate root development through the transfer of signaling molecules from shoots to roots.Activation of phytochrome A(phyA) and phytochrome B (phyB) acts as photoreceptors and transduces light signals from shoots to roots,resulting in auxin biosynthesis or redistribution in the root system,thereby stimulating root development,especially the production of lateral roots (Lee et al.2016).PhyB induces the expression ofELONGATED HYPOCOTYL5 (HY5) and promotes stabilization of the HY5 protein which moves from the shoots to the roots where it activates gene-encoding NO3−transporters,increasing its uptake (Lee et al.2016;Xuan et al.2017).
Kinetic parameters may assist in the identification of plants that are well-adapted to different edaphoclimatic conditions.Rates of N absorption have been reported for annual crops such as rice (Araújo et al.2015),corn (Horn et al.2006),barley (Glass 2003) and Chinese cabbage (Song et al.2016),and for fruit species such as grapevine (Tomasi et al.2015) and peach (de Paula et al.2018).However,for tree species,in particularEucalyptusclones,little is known about NO3−and NH4+absorption kinetic parameters.Therefore,it is expected thatEucalyptusclones possess different abilities to absorb NO3−and NH4+,and this will be reflected in the absorption efficiency and nutrient use and,consequently,in the physiological responses during growth and production.The selection of the most efficientEucalyptusclones for NO3−and NH4+uptake is recommended for low nitrogen soils,while the least efficient NO3−and NH4+uptake clone but with important wood characteristics for the consumer market is recommended for soils with higher N levels (Clough et al.2013;Rocha et al.2014).As a result,the idealEucalyptusclone for plantations in low N soils has lowC minandK mvalues and highV maxvalues,and consequently higherI(Martinez et al.2015).The results from this study may contribute to the selection ofEucalyptusclones with greater nutrient absorption capacities and zoning of clones best adapted to the soil conditions of each region,thereby contributing to increasing productivity.The objective is to selectEucalyptusclones according to their efficiency of N absorption using kinetic,physiological and morphological parameters.
Material and methods
Plant material and treatments
The experiment was conducted from September to October 2017 in the greenhouse at the Department of Soils of the Federal University of Santa Maria (UFSM),Santa Maria,Rio Grande do Sul,southern Brazil.Throughout the experiment,average temperatures of 25 °C and average relative humidity of 60% were maintained.Seedlings ofE.saligna(32,864) andE.grandis(GPC 23) clones were produced from shoots from cut matrices.Mini-cuttings of shoot branches were collected and rooted in the greenhouse.The mini-cuttings were 12-cm long with three superior buds;leaves were cut to the center of the leaf midrib,leaving 50%of the photosynthetic area and reducing the amount of transpiration.Cultivation containers were nontoxic polypropylene plastic tubes with a volume of 180 cm3,containing substrate (1:1:1 v:v:v) of carbonized rice husks,vermiculite and a commercial substrate of pine bark.In August 2017,60-d clones approximately 20-cm high were transferred to polyethylene bags and stored in a greenhouse at 10 cm × 15 cm spacing.At 90 days,fiveE.saligna(32,864) andE.grandis(GPC 23) plants approximately 40-cm in height and with 10 to 15 leaves were removed from the plastic bags,their roots washed and transferred to 8-L pots with 5 L of 25%full strength Hoagland nutrient solution (Jones 1983) where they remained for seven days until the first acclimatization step was accomplished.The 100% Hoagland nutrient solution contained (in mg L−1) NO3‒=196;NH4+=14;P=31;K=234;Ca=160;Mg=48.6;S=70;Fe-EDTA=5;Cu=0.02;Zn=0.15;Mn=0.5;B=0.5;and Mo=0.01.
The pots were placed in the greenhouse in a completely randomized design with five replications per treatment,each plant considered a repetition.A Styrofoam slice was fixed on the surface of each pot to fasten the plants,preventing the entry of solar radiation and reducing the evaporation of the solution.The Styrofoam blade had a central hole for theEucalyptusclone stem pass through and a second hole for the entrance of a PVC (polyvinyl chloride) tube connected to an oil-free air compressor for aeration.
After 7-d acclimatization of the clones in Hoagland solution,the solution was exchanged and the plants remained for 21 d in 50% full strength Hoagland nutrient solution,finishing the second acclimatization period.The solution was renewed every five days with the pH adjusted to 6.0 ± 0.2 through the addition of 1 mol L−1HCl or 1 mol L−1NaOH every two days.After the periods of acclimatization,the clones were induced to exhaustion nutrient reserves in a 0.1 mol L−1CaSO4solution for 30 d.Where Ca and S were used to maintain the electrochemical potential of cell membranes and preserve cell wall integrity (de Paula et al.2018).
Net absorption kinetics of NO3− and NH4+
After 30-d the exhaustion of nutrient reserves in a CaSO4solution (0.1 mol L‒1),the clones were returned to the Hoagland solution at 50% full strength and kept in this solution for 1 h for the system to reach steady absorption state conditions for the application of the kinetic model by Claassen and Barber (1974).Following this,the solution was replaced again,containing the same concentration of nutrients of 50% Hoagland solution to collect the first aliquots of the solution itself.Every six hours a 10-mL solution was collected from each 5 L pot at time zero before adjusting the plants in the pots with an initial solution.Aliquots of 10 mL were collected every six hours beginning at the first 30 h,every three hours between 30 and 54 h,and every hour between 54 and 65 h.The solutions were frozen at −10 °C and stored for further analysis of N compounds.
Photosynthetic parameters
The evaluation of these parameters was carried out on the third fully expanded leaf using an infra-red gas analyzer(IRGA) portable meter (Li-Cor,LI-6400 XT,United States)and 1,500 μmol m−2s−1photosynthetic active radiation and a CO2concentration of 400 μmol mol‒1.Measurements were taken between 8:00 and 10:00 am to obtain net photosynthetic rate (A-μmol CO2m−2s−1),stomatal conductance of water vapor (Gs-μmol H2O m−2s−1),intercellular concentration of CO2(Ci-μmol CO2mol−1),the transpiratory rate(E-mmol H2O m−2s−1) and instantaneous water use effi-ciency (WUE-μmol CO2mol−1H2O).These were recorded as the ratio between the CO2fixed by photosynthesis and the amount of transpired water and the efficiency of rubisco carboxylation (A/Ci-mol CO2m−2s−1).The ratio between the CO2fixed by photosynthesis and the internal concentration of CO2.
Evaluation of chlorophyll a fluorescence
Chlorophyllafluorescence was analyzed on the first fully expanded leaf of three plants per treatment on sunny mornings between 8:00 and 9:30 am (Souza et al.2013) using a portable fluorometer of modulated light (Junior-Pam Chlorophyll Fluorometer Walz Mess-und-Regeltechnik,Germany).Prior to measurements,the leaves were pre-adapted to darkness for 30 min to measure initial fluorescence (Fo).Subsequently the samples were subjected to a saturating light pulse (10,000 μmol m−2s−1) for 0.6 s.The maximum quantum yield of PSII (Fv/Fm) was obtained as the ratio between variable fluorescence (Fv=Fm‒Fo) and maximum fluorescence.The photochemical quenching coefficient (qP) was calculated as(Schreiber et al.1995).The electron transport rate (ETRm) was evaluated using induction curve fluorescence (1,500 mmol m−2s−1).
Photosynthetic pigment determination
Leaves used to evaluate chlorophyllafluorescence were collected and frozen in liquid N2for photosynthetic pigment analysis.Chlorophylla(Chla),chlorophyllb(Chlb) and carotenoid contents were analyzed according Hiscox and Israeslstam (1979),and estimated with Lichtenthaler’s formula (Lichtenthaler 1987).Fresh 0.05 gm leaf samples were incubated in 7.0 mL of dimethyl sulfoxide (DMSO)at 65 °C for two hours until the tissues were completely bleached.Pigment concentrations were calculated after absorbance reading on a Celm E-205D spectrophotometer(Bel Engineering,Italy) at 645 and 663 nm for Chlaand Chlb,respectively,and 470 nm for carotenoids.Chlorophyll and carotenoid concentrations were expressed as mg g−1fresh weight.
Plant collection and analysis of N concentration in tissues and solution
After 65-h kinetic gait evaluation,the plants were removed from the nutrient solution and fractionated into leaves,stem and roots.Height was measured and stem diameter determined using a manual caliper.Fresh root and shoot matter were weighed and the volume of nutrient solution remaining in each pot was measured.The materials were dried in an air-forced ventilation oven at 65 °C until constant mass,and then weighed to determine dry matter.
The plant tissues were ground in a 2-mm Willey mill,weighed and nitrogen concentrations determined using an elemental analyzer (FlashEA 1112,Thermo Electron Corporation,Italy).NO3−and NH4+of the 65-h solutions were determined colorimetrically using a Segmented Flow Analyzer System (SAN ++System,Skalar,Netherlands).
Root system morphology
The characterization of root morphology was obtained from digitized images using an EPSON Expression 11,000 scanner equipped with additional light (TPU) with a 600 dpi resolution.The scanned images were used to determine root morphological traits using the WinRHIZO Pro software (Regent Instrument Inc.,Canada).Total root length(cm ind.−1),surface area (cm2ind.−1),volume (cm3ind.−1),average diameter (mm) and percent distribution of fine root length (L) for each diameter class (%) of 0 <L≤ 0.2;0.2 <L≤ 0.45;0.45 <L≤ 0.75;0.75 <L≤ 1.5;L> 1.5,were obtained.
Statistical analysis
Kinetic parameter (V maxandK m) values were calculated according to the NO3‒and NH4+concentrations in the Hoagland solution,the initial and final solution volumes in the pots,and root fresh matter values using the software Influx.C minwas determined according to the concentrations of NO3−and NH4+in the nutrient solution corresponding to the 65 h of evaluation time.Influx (I) was calculated using Eq.1 by Michaelis–Menten and modified by Nielsen and Barber (1978).

where,V maxis membrane transporters’ maximum absorption rate;Cis the concentration in solution at collection time;C minis the minimum concentration at the 65-h period andK mis transporter affinity coefficient per solute.
The results from the morphological and physiological parameters were submitted to homogeneity and normality tests and subsequently,the data were processed and statistically analyzed using R statistical software (R Development Core Team 2019).When the effects of the treatments were considered significant,the results ofV max,K m,C minandIfor eachEucalyptusclone were compared by Student’st-test(P< 0.05).The difference in NH4+and NO3−concentrations over 65 h for each clone was compared by the Scott Knott test (P< 0.05),as this test is better suited to 25 times of solution collection.
Additionally,the data were also subjected to principal component analysis (PCA) using Canoco software version 4.5 (Ter Braak and Smilauer 2002).PCA is generally used to find the weight of each variable to maximize the variance among sampling points (Ortega et al.1999).Principal componsent analysis is performed according to a set of principal components (PC1 and PC2) which are composed of standardized orthogonal linear combinations that together explain the variance of the original data.This type of analysis allows for the identification of more complex interactions between the evaluated variables and those with greater contribution to the differences among treatments.
Results and discussion
Morphological parameters
TheE.grandisclone had higher values for height,dry matter production of leaves,roots,and total dry matter and the highest levels of N in leaves and roots compared with theE.salignaclone (Table 1),suggesting that the higher dry matter production of theE.grandisshoots may have increased transpiration,generating a higher water gradient between the solution and the plant,stimulating N uptake by roots (Zufferey et al.2015;Lee et al.2016).Lower values of accumulated N and dry matter in leaves and roots ofE.salignamay be attributed to lower efficiency of the clone in absorbing N which reduces nitrogen translocation to growing organs,resulting in lower heights and biomass production.Lower biomass production results from a reduction in light interception capacity,fixing less carbon which reduces the concentration of photoassimilates and absorbing less nutrients from the roots (Marschner 2012;Lee et al.2016;Canarini et al.2019).
TheE.grandisclone had greater length,surface area,root volume and percentage distribution of fine roots (< 1.5 mm in diameter) thanE.saligna(Fig.1 a,b,d,e),variables that determine the absorption rate of water and nutrients by the roots (Batista et al.2016).The higher fine root production of the clone may have been the result of the greater reception of light signals by the shoots and transfer via signaling molecules to the roots (Lee et al.2016),since the clone showed greater shoot growth (Table 1).Reception of these light signals activates the production of fine roots,increasing contact with water (Skaggs and Shouse 2008) and nutrients(Lambers et al.2006).The greater fine root production by the clone may be a strategy to increase the area of soil/solution with lower carbon investment.In addition,the accumulation of N in the roots promotes the growth ofEucalyptusclones,since roots are sinks of carbohydrates and aminoacids used to overcome nutrient deficiency conditions and thus be redistributed to growing organs (Centinari et al.2016;Klodd et al.2016).

Table 1 Morphological parameters,accumulation and total N content in tissues of E.grandis and E.saligna clones after 30-d reduced internal nutrient reserves
Kinetic parameters of NO3− and NH4+ absorption
TheE.grandisclone had lowerK mandC minvalues for NO3−compared to theE.salignaclone (Fig.2 a).TheC minresults suggest thatE.grandishas NO3−transporter proteins that are activated at lower ion concentrations in solution,and the low Kmvalues show high affinity for NO3−.In addition,theE.grandisclone had the largest length,surface area and root volume compared to theE.salignaclone (Fig.1 a,b,d),which contributed to the higher number of transporter proteins of NO3−and N absorption efficiency (Lambers et al.2006;Raven et al.2018;Canarini et al.2019).The smallerC minvalues suggest that theE.grandisclone has a higher absorption ability even in small concentrations of NO3−in the environment (solution or soil),and can access NO3−on a larger number of absorption sites per root unit in different environments relative to theE.salignaclone (Tomasi et al.2015;Batista et al.2016).This suggests that theE.grandisclone may be grown in solution or in soils with lower NO3−availability,which may occur in soils with less history of fertilizer application or in soils with low organic matter(Clough et al.2013).As a result,the risk of contamination of surface and subsurface waters by NO3−adjacent to areas cultivated withEucalyptusis also reduced (Bindraban et al.2015;Bednorz et al.2016).On the other hand,E.grandisandE.salignaclones did not differ statistically between Vmaxvalues for NO3‒,indicating that these clones have similar nutrient absorption properties in solution when all loader sites in the root cell membranes are saturated (Yang et al.2007;Martinez et al.2015).

Fig.1 a surface area,b root volume,c average diameter,d length,e percent distribution of fine roots for each diameter classes of E.grandis and E.saligna clones;Means followed by the same letter did not differ by Student’s t-test (P < 0.05)
NO3−absorption kinetics shows the differentiation in absorption between the clones as illustrated by the clone influx curve (Fig.2 a).E.grandisclone initiates absorption of NO3−in solution even at low concentrations and continues absorption even at higher levels compared toE.saligna,and therefore has lowerC minvalues (Fig.2 a).This shows that differentEucalyptusclones absorb NO3−through distinct transport systems.Thus,theE.grandisclone possibly activates a high affinity system(HATS),while theE.salignaclone may activate a low affinity system (LATS),each mediated by more than one membrane protein with different enzymatic kinetics.The molecular basis of these high and low affinity absorption systems has been identified (Dechorgnat et al.2010),mainly in theArabidopsismodel plant (Doddema and Telkamp 1979),demonstrating that a LATS belongs to the NRT1 transporter family and a HATS to the NRT2 family,except for dual affinity transporter NRT1.1 (Tomasi et al.2015).Studies indicate that a LATS linearly contributes tothe absorption of NO3−at concentrations above 250 μmol L−1and thereafter,the absorption sites become saturated at concentrations close to 50 mmol L−1inArabidopsisplants (Glass 2003).At low concentrations of NO3−in solution,two high-affinity transport systems are activated,one of them constitutive (cHATS),withK min a range of 6‒20 μmol L−1,while another induced system (iHATS)of lower affinity,occurs with Kmbetween 20–100 μmol L−1(Tomasi et al.2015).Responses similar to these were reported by de Paula et al.(2018) who observed that peach rootstocks have different NO3−transport systems (HATS and LATS),showing that the same cultivar can act in different NO3−transport systems.
TheE.grandisclone had lower values forK mandC minof NH4+(Fig.2 b) and higher values for length,surface area and volume of roots compared to theE.salignaclone (Fig.1).These results suggest that theE.grandisclone may possibly have a larger number of NH4+absorption sites per root unit(Pii et al.2014;Tomasi et al.2015).Therefore,root morphological parameters are crucial when access to nutrients,including NO3−and NH4+,is a limiting factor,revealing the adaptability ofEucalyptusclone root architecture (Gonçalves et al.2013).
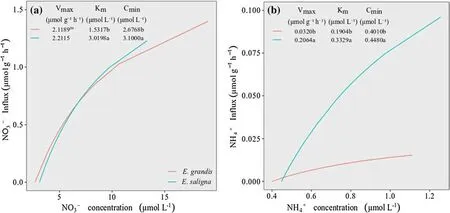
Fig.2 Influx rates and kinetic parameters of a NO3− and b NH4+ uptake of E.grandis and E.saligna clones;Means followed by the same letter did not differ by Student’s t-test (P < 0.05)
Accordingly,root morphology contributed to the kinetic NH4+absorption parameters illustrated by theE.grandisclone and possibly provided higher NH4+uptake due solely to the greater affinity of NH4+transporters,reflected in lower Kmvalues compared to theE.salignaclone(Fig.2 b).TheE.grandisclone possibly operates in a high affinity transport system,allowing NH4+absorption even when the nutrient is in low concentrations (Couturier et al.2007;Li et al.2012).This explains the greater efficiency of theE.grandisclone in absorbing NH4+being able to activate NH4+absorption sites,even though the ions are in very low concentrations in solution or the soil,allowing it to reach lowerC minvalues (Fig.2 b).In contrast,higherV maxvalues were observed in theE.salignaclone,suggesting that this clone may activate a NH4+low affinity transport system,resulting in higherK mandC minvalues.Some studies have reported that ion uptake may be mediated by high and low affinity transporters of the AMT/MEP/Rh (AMT) protein subfamily.The subfamily AMT1 is responsible for the transport of high affinity NH4+and the subfamily AMT2 for the transport of low affinity NH4+.AMTs are proteins that activate the transport of NH4+through the plasma membranes,providing the principal path for NH4+influx into the roots (Castro-Rodríguez et al.2017;Xuan et al.2017).Another factor that contributed to the absorption of NH4+from theE.grandisclone was the higher production of leaf dry matter.This increased the transpiration rate of the plants and the water gradient between the solution and the plants,allowing the nearness of NH4+to the external surface of the roots.This favors its absorption and transport,can accumulate of N in leaves and roots (Table 1) (El-Jendoubi et al.2013;Jordan et al.2014;Rivera et al.2016).
Evaluation of NO3− and NH4+ absorption over the kinetic gait period
The NO3−absorption kinetic gait demonstrated that,initially,the two clones absorbed NO3−intensely (Fig.3),possibly due to low N reserves and high nutrient demand(Tomasi et al.2015).This NO3−decay behavior in solution occurred in a circuitous pattern up to 24-h evaluation,and thereafter the decay occurred smoothly.Similar responses have been reported in NO3−absorption studies (Yang et al.2007;Pii et al.2014).The sinuous decay of NO3−in solution for the clones occurred within 24-h evaluation.This shows that the initial uptake by theEucalyptusclones was similar,and that uptake occurred through a low affinity transport system (LATS).However,the differentiation of root morphological characteristics between clones provided different mechanisms of NO3−absorption.After 24 h,the clones possibly initiated the absorption of NO3−through another transport system (HATS) until the absorption of NO3−decreases,reaching Cmin,showing the highest absorption efficiency of theE.grandisclone.This allowed for the absorption of NO3−in low concentrations in solution.As a result,lower values ofK mandC minwere achieved.TheE.salignaclone absorbed NO3‒ less intensely,reachingC minonly after 65-h appraisal (Fig.3).However,theE.grandisclone absorbed NO3−more continuously than the other clone,reachingC minat 64-h evaluation.The results show thatEucalyptusclones differ in NO3−absorption intensity,and this correlates with the genetic characteristics of each clone (Tomasi et al.2015;Kiba and Krapp 2016).

Fig.3 Concentration of NO3−in nutrient solution with E.grandis and E.saligna clones after 30-d internal nutrient depletion;Average NO3− concentrations in blue and green differ significantly from averages of concentrations in red α=0.05 (Scott Knot’s test).a Time reaching lowest concentration P < 0.05
The NH4+absorption kinetic gait showed that initially absorption occurs similarly in both clones.Initial uptakeof NH4+is intense by the roots,with a more winding NH4+decay behavior in solution up to 12-h evaluation,and then the decay occurs less intensely until 42-h evaluation (Fig.4).This possibly was caused by the low induction of proteins that act on the transport of NH4+in the root plasma membranes where the main route of NH4+influx is mediated by NH4+transporter proteins (AMTs).This may occur due to NH4+saturation at these absorption sites,differentiating an initial NH4+depletion stage in the solution between two absorption mechanisms.A low affinity NH4+uptake mechanism possibly occurs up to 12 h in the two clones,which are saturated until near 42-h evaluation.Low affinity NH4+transport is mediated by AMT2 proteins and then another high affinity NH4+uptake mechanism activates,thus activating NH4+transporter proteins (AMT1) (Castro-Rodríguez et al.2017).As a result,the absorption of NH4+by the transporters decreases until reachingC minat 48-h evaluation(Fig.4) but with a difference in the NH4+concentration in solution which shows the differentiation between the clones regarding ion extraction capacity.This justifies lower values ofK mforE.grandiswhen compared toE.saligna.
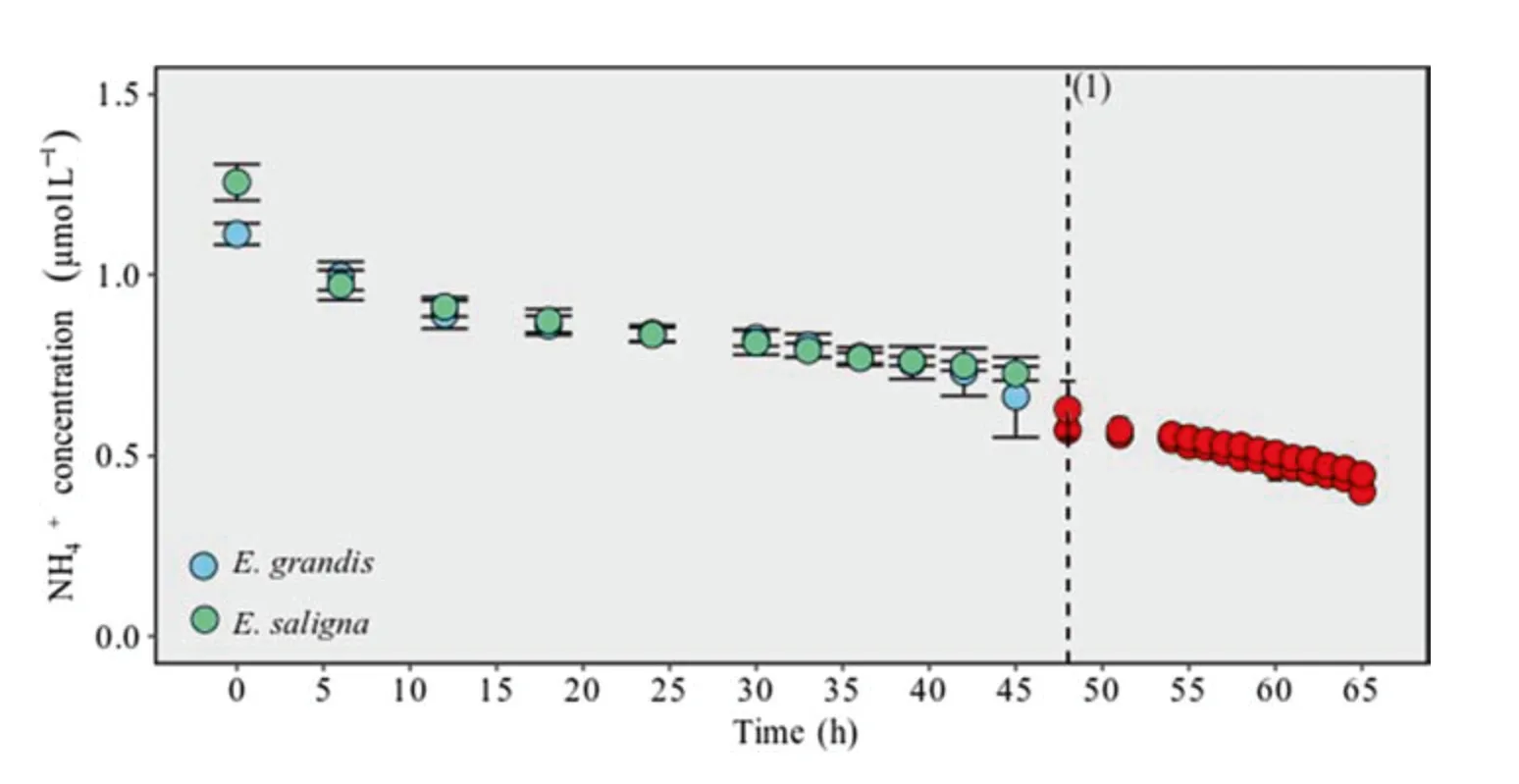
Fig.4 Concentration of NH4+in nutrient solution with E.grandis and E.saligna clones after 30-d reduced internal nutrient reserves;Average NH4+concentrations in blue and green differ significantly from averages in red α=0.05 (Scott Knot´s test).a Time reaching lowest concentration P < 0.05
It should be noted that the clones continued to absorb NO3−and NH4+over the 65-h evaluation period and only after 65 h reachedC minfor NO3−and 48 h for NH4+.This shows the importance of the collecting solutions at more spaced out periods in the initial hours of absorption,compared with the more 5 h of evaluation as used for cabbage(Song et al.,2016),8 h for rice (Araújo et al.2015),24 h for corn (Horn et al.2006),and grapes (Tomasi et al.2015).In addition,at the end of the evaluation period,sampling should be performed in shorter periods so that it is possible to note with more accuracy the actual moment plants reachC min.
Gas exchange parameters
Significantly higher levels of intercellular CO2concentration were observed inE.grandis(Fig.5 c).This was because the clone had the highest values of accumulated N in leaves and shoot dry matter production.Therefore,the availability of CO2was maximized and the assimilation of C from photosynthesis was assisted.This contributed to greater CO2fixation in leaf tissues (Martim et al.2009;Tcherkez et al.2017).These results corroborate studies with perennial crops which show that the increase in leaf N content correlates with an increase in the CO2absorption rate (Jennings et al.2016;Greer 2018).The results suggest that theE.grandisclone converted greater amounts of CO2per leaf tissue area,highlighting the importance of clone genotype on photosynthetic parameters (Nadal and Flexas 2018).However,no significant difference was observed between the two clones for net photosynthetic rate (Fig.5 a).However,this higher intercellular CO2concentration may be the result of greater respiration (Tcherkez et al.2017).This confirms the greater efficiency of theE.grandisclone in the absorption of NO3−and NH4+and,consequently,greater accumulation of N in the leaves and assisting in photosynthesis by contributing important chloroplast proteins (Blank et al.2018;Hu et al.2019;Moriwaki et al.2019).
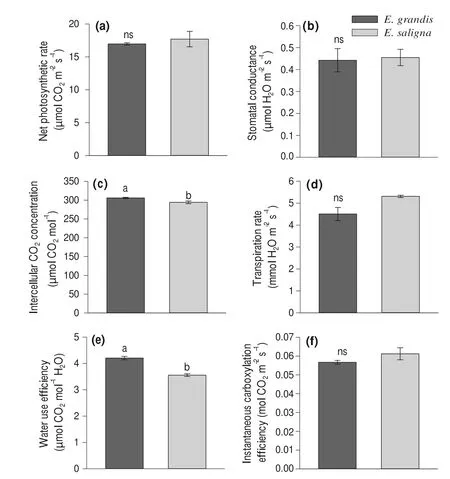
Fig.5 a net photosynthetic rate,b stomatal conductance,c intercellular CO2 concentration,d transpiration rate,e water use efficiency and finstantaneous carboxylation efficiency in leaves of E.grandis and E.saligna clones;Means followed by the same letter did not differ by Student’s t-test (P < 0.05)
TheE.grandisclone showed significantly higher water use efficiency (WUE) than theE.salignaclone (Fig.5 e).Although there was no statistical difference in rate of transpiration (Fig.5 d),theE.grandisclone lost less H2O per unit area because transpiration is expressed as mmol H2O m−2s−1.This may be related to internal CO2concentration,resulting in greater leaf vigor which may result in greater WUE (Fig.5 e).In addition,water use efficiency is an important metric for indicating plant stress and demonstrating crop suitability under different edaphoclimatic conditions (Wu et al.2018).However,theEucalyptusclones did not differ statistically in net photosynthetic rate,stomatal conductance of water vapor,rate of transpiration and instantaneous rubisco carboxylation efficiency (Fig.5 a,b,d,f).
TheE.grandisclone had the lowest photochemical quenching coefficient (qP) compared to theE.salignaclone (Fig.6 a),and consequently had the highest maximum quantum yield of photosystem II (Fv/Fm) and effective quantum efficiency of PSII (Y(II)) (Fig.6 b,d).These results indicate that theE.grandisclone transfers more excitation energy from the light collecting system to the reaction center,and more energy directed to the photochemical reaction (Wang et al.2019).This illustratesthat plants with less energy loss reflect higher shoot dry matter production (Table 1).TheE.grandisclone uses more energy directed to the photochemical stage of photosynthesis,converting more light energy into chemical energy.Therefore,the lower the dissipation of energy in the form of fluorescence,the greater the formation of ATP and NADPH and,consequently,the greater the photosynthetic C assimilation.

Fig.6 a photochemical quenching coefficient (qP),b maximum quantum yield of PSII (F v/ F m),c electron transport rate (ETR m) and d effective quantum efficiency of PSII (Y(II) in E.grandis and E.saligna clone leaves;Means followed by the same letter did not differ by Student’s t-test(P < 0.05)
TheE.grandisclone had the highest carotenoid content(Fig.7),indicating higher levels per leaf area,allowing greater energy absorption and transfer in photosynthesis as carotenoids are responsible for light absorption in different regions of the spectrum in early stages of photosynthesis.In addition,the photochemical phase is only accomplished if there are sufficient pigments to interact with photosynthetic radiation.The higher carotenoid levels in theE.grandismay also protect against excess light,as carotenoids,besides acting as accessory pigments,are also photoprotective agents (Marschner 2012).Levels of chlorophylla,band total chlorophyll did not differ statistically between clones (Fig.7).However,the concentration of photosynthetic pigment contents in theE.salignaleaves may be attributed to lower dry matter production (Table 1).Thus,larger amounts of pigments are visualized per unit of mass,resulting in a concentration of photosynthetic pigments,allowing close values between the clones.

Fig.7 Pigments concentration of chlorophyll a (Chl a),chlorophyll b (Chl b),total chlorophyll (Chl total) and carotenoids in leaves of E.grandis and E.saligna clones;Means followed by the same letter did not differ by Student’s t-test (P < 0.05)
Principal component analysis (PCA)
PCA was carried out by extracting only the first two components,PC1 and PC2,in which their sum explained 72.95%of the original data variability (Fig.8).Of this,53.72% were explained by PC1 and 19.23% by PC2.The PCA results show two clusters of data,highlighting the differentiation ofE.salignaandE.grandisclones.The variables with the greatest influence on the group formed by theE.grandisrepetitions were height (h);leaf accumulated N (LAN),stem accumulated nitrogen (SAN) and root accumulated nitrogen(RAN);leaf dry matter (LDM),stem dry matter (SDM),root dry matter (RDM) and total dry matter (TDM);root surface area (RSA),root length (RL),root volume (RV) and root diameter (RD);photosystem II quantum yield (Fv/Fm),maximum fluorescence (Fm),water use efficiency (WUE),CO2intercellular concentration (Ci) and carotenoids (carot).In contrast,theE.salignaclone was influenced by the variables Cminof NO3−and NH4+,Kmof NH4+,V maxof NH4+,minimum fluorescence (Fo),electron transport rate (ETRm),net photosynthetic rate (E),stomatal conductance (Gs) and instantaneous carboxylation efficiency (A/Ci).
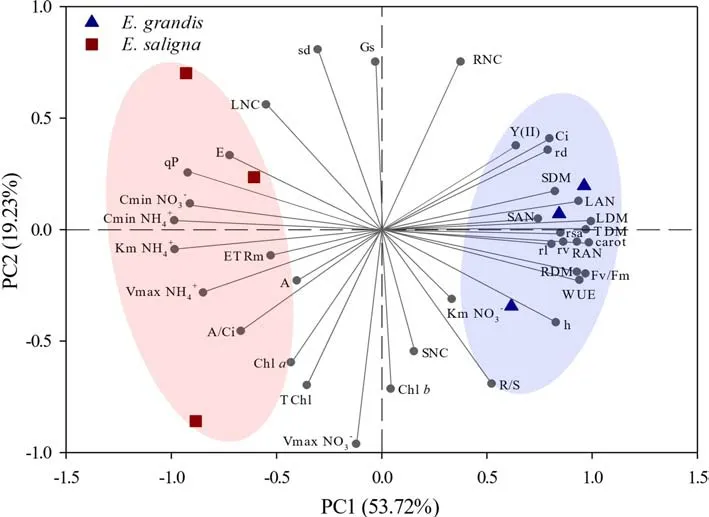
Fig.8 Scatter plot of principal component analysis (PCA) of kinetic parameters of NO3− and NH4+ (V max ;K m ;C min),morphological(height (h);stem diameter (sd);dry matter in leaves (LDM),in stem(SDM),in roots (RDM),in total (TDM);root/shoot ratio (R/S);total N in leaves (LNC),in stems (SNC),in roots (RNC);N accumulated in leaves (NLA),in stem (NSA),in roots (NRA)),root morphological parameters (surface area (rsa);volume (rv);diameter (rd);length (rl))and physiological parameters (photochemical quenching coefficient(qP);effective quantum efficiency of PSII (Y(II));electron transport rate (ETRm);maximum quantum yield of PSII (Fv/Fm);net photosynthetic rate (E);stomatal conductance (Gs);intercellular CO2 concentration (Ci);transpiration rate (A);water use efficiency (WUE);instantaneous carboxylation efficiency (A/Ci);concentration of chlorophyll a (Chl a),b (Chl b),total (Chl total);carotenoids in leaves) in E.grandis and E.saligna clones
Kinetic parametersC minof NO3−and NH4+,Kmof NH4+,V maxof NH4+,were negatively correlated with root morphology,length,diameter,area and volume,and correlated positively with theE.grandisclone.This suggests that the higher the development of theEucalyptusclone root system,the lower theC minandV maxof NO3−and NH4+,and Kmof NH4+.This is important because the lower their values,the greater the absorption efficiency of NO3−and NH4+and the lower the concentration at which roots will be able to extract the nutrient from the solution.In addition,these results demonstrate that as the plant invests photoassimilates in the roots,the likelihood of water and nutrient absorption increases and this results in lowerC minvalues,as plants will be able to access more restricted areas and lower concentrations of elements.Combined with this,in this grouping there is a positive correlation with the increase of nitrogen in the leaves,stem and roots.This confirms the positive correlation carotenoid pigment levels per leaf area which helps the assimilation of intercellular CO2provided by quantum yield of photosystem II,thereby assisting the development of theE.grandisclone.Thus,higher heights,stem diameters,and leaf,stem and root dry matter production were observed.
Another grouping differentiates theE.salignaclone and shows the strong influence of kinetic parameters on the absorption efficiency of different forms of nitrogen and,consequently,the accumulation of N in organs.TheE.salignaclone was positively correlated with theC minvariables of NO3−and NH4+,K m of NH4+,which is not desirable as it shows a lower affinity for NO3−and NH4+absorption.This confirms the positive correlations with physiological parameters such as photosynthetic stress.This suggest that the clone may have suffered damage to the PSII reactive center,decreasing the efficiency of excitation energy transfer from the light collecting system to the reaction center.This result in lower development of leaves,justifying the inverse correlation with dry matter production (LDM).
Conclusions
TheE.grandisclone was more efficient in the absorption of NO3−and NH4+and had kinetic parameters with lower values ofC minandK mcompared to theE.salignaclone.Root morphological parameters such as area,volume and length are positively related with kinetic absorption parameters such as lowerK mandC minand can be used in selection and breeding programs ofEucalyptus.However,the minimum time for kinetic gait assessment to reachC minforEucalyptusclones should be 65 h for NO3−and 48 h for NH4+.Kinetic gait studies help in understanding nutrient absorption,and the results of this study may contribute to the selection of more efficientEucalyptusclones in absorbing forms of nitrogen and assist in nitrogen fertilization strategies.
杂志排行

Journal of Forestry Research的其它文章
- Flexible transparent wood enabled by epoxy resin and ethylene glycol diglycidyl ether
- Diversity and surge in abundance of native parasitoid communities prior to the onset of Torymus sinensis on the Asian chestnut gall wasp (Dryocosmus kuriphilus) in Slovenia,Croatia and Hungary
- Ozone disrupts the communication between plants and insects in urban and suburban areas:an updated insight on plant volatiles
- Testing visible ozone injury within aLight Exposed Sampling Site as aproxy for ozone risk assessment for European forests
- Logging and topographic effects on tree community structure and habitat associations in a tropical upland evergreen forest,Ghana
- Spatial pattern dynamics among co-dominant populations in early secondary forests in Southwest China