Picea species from humid continental and temperate marine climates perform better in monsoonal areas of middle latitudes of China
2021-07-15FangqunOuyangJianweiMaJunchenWangLishengKongHanguoZhangMulualemTigabuJunhuiWang
Fangqun Ouyang·Jianwei Ma·Junchen Wang·Lisheng Kong·Hanguo Zhang·Mulualem Tigabu·Junhui Wang
Abstract Picea spp.are a prominent component of the boreal,montane,and sub-alpine forests in the Northern Hemisphere,and have substantial economic importance due to their high quality fibre.However,performance and wood properties of the genus have not been systematically evaluated in China.This study aimed to examine genetic variations in growth traits,phenology,and wood properties of 17 Picea species,(three from North America,two from Europe,and 12 from China),in response to specific climate factors using a randomized complete block design in a monsoonal,middle latitude area of China.Results show that all growth traits and wood properties significantly varied among species (P < 0.00).Of the 17 species examined,P.abies and P.pungens were the tallest with heights of 2.5 and 1.9 m at 9 years old,respectively,90% and 50% greater than the average heights.Branch length,number,and angles of both P.abies and P.pungens were greater than those of the other species.Heights of P.glauca and P.omorika were 20–33% greater than the average.Fast-growing species had high quantities of first lateral branches and large top whorl branch lengths.The taller species exhibited greater tracheid lengths and average tracheid lengths to radial central diameter ratios (TL/R_D2),but smaller cell wall thicknesses to tracheid radial lumen diameter ratios (WT/R_D1),which is favorable for pulp production.Correlation analysis revealed that height and ring width had significant positive correlations with latitude but strongly negative correlations with longitude.Height was positively correlated with average annual rainfalls,but negative correlations with average yearly sunshine.Due to the increased average annual sunshine at the China test site relative to the seed source,species such as P.abies,P.pungens,P.glauca,and P.omorika from wet,humid areas performed better than native species.Considering similar temperatures,precipitation may be the main factor affecting growth,which is beneficial for predicting the extent of spruce expansion.These results should encourage further testing and provide reference information for future exotic species studies in this part of China.
Keywords Climate·Growth performance·Genetic variations·Picea ·Wood properties
Introduction
PiceaMill.(Spruce) is a genus of approximately 34 species,the third largest in the Pinaceae family,and widely distributed throughout the Northern Hemisphere.The largestPiceacenter of distribution and differentiation is in Asia (Farjon 2001),with the most numerous species,including seven endemics in China (Fu et al.1999;Farjon 2001).Piceaare prominent components of the boreal,montane,and subalpine forests in the Northern Hemisphere and have substantial economic importance due to their high quality fibre.Several studies have reported that the phylogenetic relationships of the genus show significant topological incongruencies between phylogenetic species based on morphology,DNA,mtDNA and cpDNA (Ran et al.2006;Bouillé et al.2011;Lockwood et al.2013;Jia et al.2014;Sullivan et al.2017).Despite the complex evolutionary history,infrageneric classification of the genus has clearly demarcated categories,although even this remains quite controversial (Fu et al.1999;Farjon 2001).
Some non-native spruce species,which have been recently introduced to China,show better growth than the native species such as the Korean spruce,P.KoraiensisNakai,and the Qinghai spruce,P.crassifoliaKom.(Zhang and Zhang 1995;Liu et al.2003;Li et al.2012;An et al.2018).The results of introductory and early evaluation studies showed thatP.sitchensis,P.abies,P.glauca,andP.marianahad superior performance compared with native Chinese species (Liu et al.2003;Li et al.2012).P.abies,P.glauca,andP.marianawere derived from the best provenances,latitude 49° 16′–58° 38′ N and longitude 68° 13′–118° 24′W,and are suitable for introduction and cultivation in Xiaolongshan within China’s Gansu Province (An et al.2018).However,there are few reports that systematically comparePiceawood properties.Sudo (1955) described the anatomical characteristics of 18 spruce species and one variety,and discussed the relationship between wood anatomical identification,growth characteristics,and their natural regions.Yang and Lu (1998) studied tracheid traits of 37 species and varieties and the results showed that Sitka spruce,P.sitchensis(Bong.) Carr.and Taiwan spruce,P.morrisonicolaHayata showed the highest values for earlywood and latewood tracheid lengths.However,there are few published reports on the growth performance and wood anatomy ofPiceacultivated under the same growing conditions,and this is especially so for continental climates.
Climate is the major influence on tree growth (Mäkinen et al.2002),with temperature and precipitation the primary factors (Egorov and Afonin 2018).Forests in mountainous areas are limited by temperature,whereas growth at lower altitudes is generally limited by precipitation (Rybnícek et al.2012).The northern distribution ofP.glaucain North America is limited by temperature,while the distribution of species in arid zones is determined by the hydrothermal coefficient which should not exceed 1.2 (Egorov and Afonin 2018).It has been confirmed thatP.abiestree ring widths in the central part of the Ceskomoravská Upland (Czech Republic) display a positive correlation with summer precipitation and a negative correlation with summer temperatures (Rybnícek et al.2012).At the same time,the growth and adaptation ofPiceaspecies are also affected by latitude,longitude,and elevation (Rożkowski et al.2001;Collignon et al.2002).These observations reflect a strong dependence on latitude for the growth ofP.abiesin Nordic-Baltic populations of Europe (Collignon et al.2002).Generally,flushing and growth cessation occur early at high latitudes–such as in Fenno Scandinavia,and at high altitudes–such as the central Alps (Krutzsch 1992).Usually,the species of high altitudes are not suitable for growth in low altitude areas.The only population from a high elevation,i.e.,1420 m a.s.l.,performed poorly as expected when growing at a low elevation (Rożkowski et al.2001).Good growth was noted for some low elevation populations,< 300 m a.s.l,as well as for populations at moderate elevations,600–700 m a.s.l(Rożkowski et al.2001).Conifer species have been evaluated based on combinations of growth traits and wood properties due to the negative correlations between growth and wood density as well as the fibre traits observed inPiceaand other conifer species (Hannrup et al.2004;Lenz et al.2011;Fries 2012;Chen et al.2014,2016;Ouyang et al.2018).Selecting for faster height growth in Scots pine resulted in a minor increase in fibre width but a slight decrease in fibre length (Fries 2012).Selection for increased diameter over time increased tracheid wall thickness and diameter,but decreased length and tracheid length to diameter ratio–all of which are unfavorable for papermaking (Ouyang et al.2018).Nevertheless,there are limited reports on the growth and wood properties ofPiceaspecies from different longitudes,latitudes,altitudes,and climates when grown together in the same environment in China.SeventeenPiceaspecies,of which three from North America,two from Europe,and 12 from China,were grown in the same locale to evaluate their growth and wood traits.The specific objectives were to:(1) compare the growth performance and wood properties of spruce species from different climates grown at the same site;(2) investigate the relationship between growth traits and wood tracheid traits at the interspecific level;and (3)determine the relationship between trait performance and climate/geographic factors.The results will aid in understanding the fitness of spruce species in non-native environments and provide reference for further application of exotic spruce as timber forests.
Materials and methods
Experimental materials and study design
The experiment was conducted in a nursery at the Research Institute of Forestry of Xiaolong Mountain in Gansu Province,China (34° 28′ 50″ N,105° 54′ 37″ E).The meteorological data for the experiment site and for the geographic sources of the 17Piceaspecies is shown in Table 1.The seeds were sown in 2008.After three years of cultivation,the seedlings were planted in the nursery at a spacing of 1.5 m.× 1.5 m.Altitude is 1160 m a.s.l;annual precipitation 600–800 mm;average annual temperature 10.7 °C,soil nitrogen content 62.5 mg kg−1;phosphorus 32.3 mg kg−1;potassium 186.9 mg kg−1;soil organic matter,0.64%;and pH 8.0.The experiment was a randomized complete block design with 30 individuals for each replication,with three replicates for each species.
Growth performance,phenology and wood traits
Ten trees were randomly selected from each block for a total of 30 trees.Heights,number of first lateral branches,length of the top whorl of branches,and branching angle were measured for the 30 trees at age nine.A protractor was used to measure the branch angle between the branches of the top whorl and the trunk to obtain the average branch angle.Bud burst and bud set were recorded for the same lateral branch for 10 trees of each species;the growth period for each species throughout a whole year was then calculated.Bud burst consisted of when the top buds were beginning to swell and appear green in spring (Supplemental Fig.S1).Bud set was identified by spiraling of the top needles and the needles surface developing a waxy,white coat (Supplemental Fig.S2).
Wood properties and tracheid traits were measured according to Ouyang et al.(2018).Ring width was measured using a ring width tester (Alfred J.Amsler &Co.,Schaffhausen,Switzerland) with an accuracy of 0.01 mm.Tracheid traits were determined using the nitric and chromic acid mixture segregation Tayler staining method (Heimsch and Tschabold 1972).Tracheid lengths were measured using a projection microscope (XST-2;Nanjing Jiangnan Novel Optics Co.,Ltd.,Nanjing,China) after staining with safranin.Fifty earlywood or latewood tracheids from each tree were randomly selected from the 6-year ring to measure tracheid length,and the average tracheid length calculated.The wood samples were cut into 10.00–15.00 μm crosssections with diagonal serial sections from pith to bark with a wood slicing machine (1400 Slip type;Leitz,Wetzlar,Germany).After the cross-sections were dried and gradually dehydrated with ethanol,the edges of the coverslip were sealed with rubber cement.Fifty valid tracheids were randomly selected from the annual ring and photographed with a P4 computer-aided system consisting of a Nikon 80i microscope (Nikon,Tokyo,Japan) connected to a DS-Ri1 digital photography transducer (Nikon).Cross-sectional dimensions of individual tracheids were measured using a TDY-5.2 Micro-color Image Analysis Software (Beijing Tiandiyu Science and Technology Co.,Ltd.,Beijing,China)at a magnification of 640 × 480 (the spatial calibration system corresponded to 0.10 μm/pixel).Measurements were taken at a minimum of two points.In the aggregate,four tracheid dimensions were measured from individual trees,including:length (TL),calculated by averaging the earlywood and latewood tracheid lengths;earlywood cell wall thickness (WT);earlywood radial lumen diameter (R_D1);and,earlywood radial central diameter (R_D2) at the 6-year ring.Average values for all four parameters were calculated as was the cell wall thickness to radial lumen diameter ratio(WT/R_D1) and the average tracheid length to radial central diameter ratio (TL/R_D2).
Statistical analysis
The data were collated and analyzed with a General Linear Model (GLM) using an ANOVA with species as a random effect and ‘e’ as a vector of random residual errors.Pearson product-moment correlation and multiple comparisons using the Duncan method (alpha=0.05 level) were also conducted.All statistical analyses were performed using the SAS statistics software (SAS Institute 1990).
Results
Variation of growth and branch traits
There were significant differences (P< 0.00) in height,the number of first lateral branches (BN),the length of the top whorl of branches (BL),and branching angle (BA)among the 17 species.P.abieswas tallest (2.48 m),90%greater than the average height of all species (1.24 m).The height of individualP.abiespeaked at 4.00 m (Fig.1),and the mean annual height increase was approximately 35.0 cm.The second tallest species,P.pungens,measured 1.93 m in height which was approximately 50% more than the average height of all species.Average annual height increase was approximately 25.0 cm.Heights ofP.likiangensis,P.linzhiensis,P.glauca,andP.omorikawere 20–33% above the average,with mean annual height increases of 20.0–28.0 cm.P.purpurea,P.Koraiensis,P.balfouriana,andP.marianaheights were intermediate.Piceacrassifolia,P.retroflexa,P.spinulosa,andP.wilsoniiheights were below average with an annual increase of 9.0–18.0 cm.Piceahirtella,P.meyeri,andP.schrenkianagrew slowly with a mean annual increase of 6.0–13.0 cm,resulting in their average,collective height lagging behind by 40–60% compared with the average height of the 17 species (Fig.1 a).

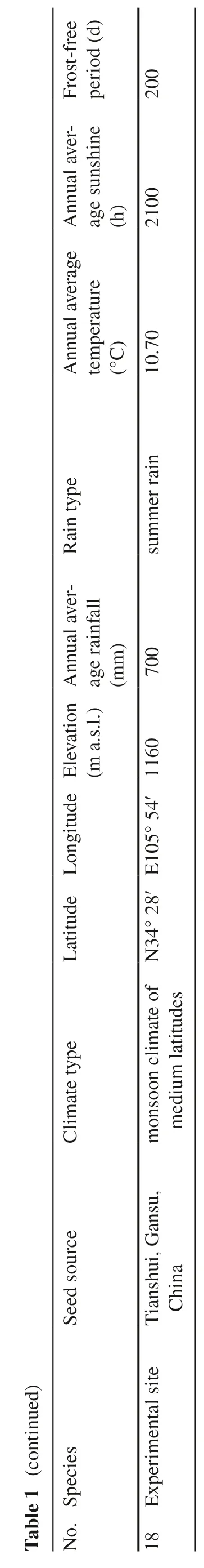
With respect to branch angles (BA) amongst the 17Piceaspecies,the two tallest produced the largest BA(> 60°);P.likiangensis,P.glauca,andP.purpureathe smallest (30–40°);the other species had intermediate BA values (50–60°) (Fig.1 b).In addition,branch numbers(BN) and branch lengths (BL) of the two tallest species were also larger (Figs.1 c,d).ForP.abies,the BN and BL were 36.0 and 58.0 cm,36.6% and 71.4% greater than the average,respectively.The BN ofP.pungens(38) was slightly larger than that ofP.abies,whereas in contrast,its BL (56.0 cm) was only 2.0 cm shorter than that ofP.abies.The maximum BN was observed inP.mariana(41),and was 55% higher than the average;P.omorikaBN was 16%higher than the average,whileP.likiangensis,P.crassifolia,P.jezoensis,P.linzhiensis,P.Koraiensis,P.schrenkinana,andP.hirtellabranch numbers were 13%–38%lower than average.Branch numbers of the other spruce were intermediate.The BL and BN value trends were not parallel.Specifically,branch lengths ofP.glauca,P.linzhiensis,P.likiangensis,andP.crassifoliawere approximately 20–60% greater than the average BL;whereas the BL value ofP.wilsonii,P.hirtella,P.spinulosa,P.retroflexa,and P.schrenkinana were about 20%–60% less than the average.
Phenology
Phenological data revealed small differences in bud burst and bud set timing among species.Bud burst ranged from early March to the third week of April,while bud setting began late May to the third week of June (Table 2).However,the growing period differed markedly among species(Table 2).P.pungensandP.abies,the two tallest species,had a short growing period,especiallyP.pungensat 38 days.P.wilsoniialso had a short growth period of 47 days.However,the growing periods ofP.glauca,P.spinulosa,P.omorika,andP.linzhiensiswere 95 days,80 days,77 days,and 68 days,respectively,46%,24%,21%,and 9% higher than the average growing period of 62 days.The other species had growth periods of 56–62 days (Table 2).
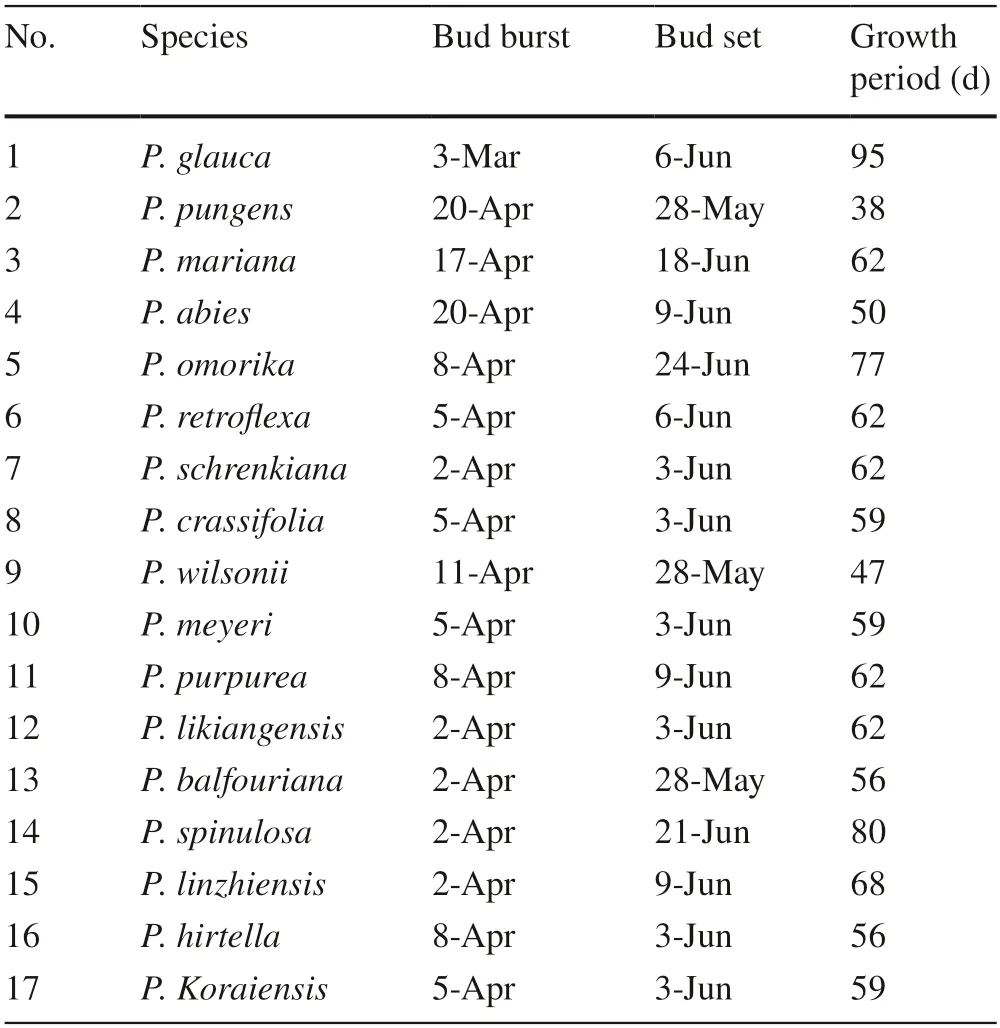
Table 2 Phenology data (bud burst,bud set,and growth period) of 17 species
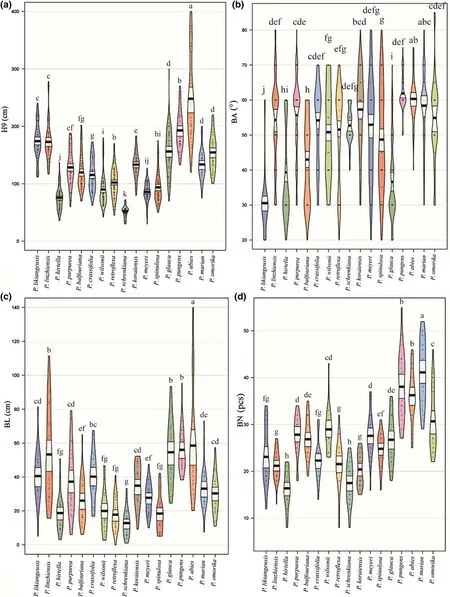
◂Fig.1 The average height at nine years old (H9),branching angle(BA),the number of first lateral branches (BN),and the length of the top whorl of branches (BL) of 17 spruce species.Note:A pirate plot is the plotted RDI (raw data,descriptive statistics,and inferential statistics) selected from R pirates that are displaying the relationship between one and three categorical independent variables,and one continuous dependent variable.The Pirate Plot includes five main elements:points that represent the raw data;vertical bars that show central tendencies;a bean that represents a smoothed density;a rectangle that depicts the inference interval;and a black line in the middle that marks the median of a raw data set
Wood proportion and tracheid trait variation
There were significant differences in ring width,tracheid length,cell wall thickness,radial lumen diameter,radial centre diameter,ratios of cell wall thickness to radial lumen diameter (WT/R_D1),and average tracheid length to radial centre diameter ratio (TL/R_D2) (P< 0.01,Fig.2 a–g).The average ring width was 1.7 mm,with a 33% coefficient of variation.Piceaomorika had the largest ring width (2.6 mm),withP.glaucawith the second largest (2.3 mm).No significant differences were observed in ring widths amongP.abies(2.0) andP.purpurea(2.0).Piceawilsonii andP.schrenkianahad the narrowest ring widths at 1.1 (Fig.2 a).The average values of the tracheid traits,length,cell wall thickness,lumen diameter,radial central diameter,WT/R_D1,and TL/R_D2,were 1.3 mm,4.6 μm,16.4 μm,21.0 μm,1.4 mm,0.3,and 0.1,respectively.For these traits,the CV was 10–20%.There were no significant differences in tracheid lengths ofP.abies(1.5 mm) andP.purpurea(1.5 mm),which had the longest tracheids.Tracheid lengths were shortest inP.schrenkiana(1.0 mm).The other species had intermediate tracheid lengths,fluctuating between 1.1 and 1.4 mm(Fig.2 b).Piceaabies had the largest radial lumen diameter(19.6 μm) and radial central diameter (24.2 μm),which did not differ significantly fromP.likiangensis,P.omorika,P.purpurea,orP.balfouriana(Fig.2 d,e).Piceaspinulosa had the highest TL/R_D2 ratio which did not differ significantly from those ofP.abies,P.pungens,P.mariana,P.Koraiensis,P.purpurea,andP.retroflexa(Fig.2 g).In contrast,the WT/R_D1 ratios ofP.purpureaandP.balfourianawere the lowest with no significant difference fromP.abies,P.pungens,P.likiangensis,P.Koraiensis,P.crassifolia,andP.wilsonii(Fig.2 f).P.schrenkianathe highest WT/R_D1 (tracheid radial lumen diameter) ratios,even though it had the smallest tracheid length and TL/R_D2(tracheid length to radial central diameter) ratios.WT/R_D1 and TL/R_D2 exhibited a linear relationship with the average height at nine years at the species level.To be more specific,the WT/R_D1 showed a significant negative relationship (r2=0.34),while the TL/R_D2 showed a positive relationship (r2=0.11) (Fig.3).Regression analysis can be used to predict fiber character WT/R_D1 and TL/R_D2 by tree height for spruce species.

◂Fig.2 The average value of 17 spruce species for the traits ring width (RW,mm),tracheid length (TL,μm),cell wall thickness (WT,μm),radial lumen diameter (R_D1,μm),and radial central diameter(R_D2,μm).The traits were measured from individual trees and average values were calculated.The ratio of cell wall thickness and radial lumen diameter (WT/R_D1) and the ratio of average tracheid length and radial central diameter (TL/R_D2) are also given
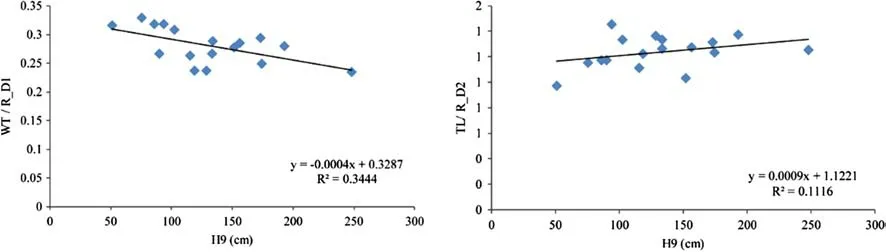
Fig.3 Regression analysis for height at nine years old (H9) and the ratio of cell wall thickness and radial lumen diameter (WT/R_D1) and the ratio of average tracheid length and radial central diameter (TL/R_D2) at the species level
Pearson correlation among growth traits,wood traits,and climate factors
There were positive correlations between height with branch number and length (Fig.4).Positive correlations were found between tree height and ring width,radial lumen diameter,radial centre diameter,and TL/R_D2.A strong positive correlation was also observed between tree height and tracheid length.Similarly,branch length showed positive correlations with ring width,tracheid lengths,radial lumen diameters,and radial central diameter.Both height and branch length were negatively correlated with WT/R_D1.The length of the growing period was strongly negatively correlated with branch angle,and positively with ring width (Fig.4).
With regards to the correlation of geoclimatic factors with growth and wood properties,height,branch number and ring width were negatively correlated with latitude but positively with longitude.In addition,height and branch number were positively correlated with annual rainfall,while height and branch length were inversely correlated with annual average sunshine (Fig.4).
Discussion
According to the results of this study,the five introduced species (three from North America and two from Europe)grew better than native Chinese species,especiallyP.abiesandP.pungens.At nine years,these two species were 2.5 and 1.9 m tall,respectively,and 90% and 50% taller than the average height of the 17 species.In addition,these fast-growing species exhibited larger tracheid lengths and smaller WT/R_D1 ratios,which are advantageous for pulp production.The best-quality fiber for paper-making are those that are thin and long,and have small wall thickness to fiber lumen diameter ratios.According to Runkel’s index of raw paper material,if the ratio of wall thickness to lumen diameter is > 1,then the raw material is of inferior quality.When the ratio is ≤ 1,the raw material is considered to be good quality.It is generally believed that fibers with smaller wall thickness to lumen diameter ratios are soft and easily crushed,enabling large fiber combinations,tight configurations,and smooth paper.Excepting tear strength,all other properties such as tensile strength,bursting strength,and folding endurance are high.Smaller ratios are also favorable for pulping which is beneficial for improving paper strength and making sheet paper (Cui 2006).
Relationship of trait performance with climate factors of test species
Piceagrowth can be affected by different climatic factors in different geographic regions (Wimmer and Grabner 2000;Koprowski 2013;Singh et al.2016).P.abiesgrowth is often influenced by air temperature and precipitation (Mäkinen et al.2002;Rybnícek et al.2012).The limiting effect of low temperatures was more significant in northern regions and high-altitude sites,while the importance of precipitation increased in the southern regions and low altitude sites(Mäkinen et al.2002).Temperature functions as a predominant selective force for the geographic distribution ofP.abies(Bergmann 1978);however,in this study,temperature weakly correlated withPiceaspp.growth since the site temperature was ultimately higher than or similar to that of the introduced species origin.Warm winters and cool summers with fairly high precipitation are the favored conditions for most of the spruce species (Li et al.2016).The correlation results also showed that spruce growth had positive correlations with annual average rainfall.In addition,height,the number of first lateral branches,and ring width had strong negative correlations with longitude.Longitude mainly affected land and sea locations.Specifically,species from Canada,the United States and Norway (P.glauca,P.mariana,P.pungens,P.abies) were located near the ocean.Thus higher precipitation is beneficial for their growth.Despite its northerly latitude,Norway (the source ofP.abiesseed) is influenced by the North Atlantic current and has a generally temperate maritime climate which is mild and humid all year round (Table 1).The two spruce (P.glaucaandP.mariana)both showed superior growth to the other species in the area in China where this research was conducted.Climate conditions in the Gansu Province are similar to the temperate zone of North America.Thus trees from North America,such asP.glaucaandP.mariana,exhibited excellent growth performance in China (Jia et al.2002;Dong et al.2007).P.marianacomes from New Brunswick,Canada,which has a temperate marine climate,also exhibited better growth than most of the species originating from China.

◂Fig.4 Pearson correlation among growth traits,wood traits,and climate factors.The darker the color,the stronger the correlation.(average height at nine years old (H9),branching angle (BA),the number of first lateral branches (BN),the length of the top whorl of branches(BL),growth period (GP),ring width (RW),tracheid length (TL),cell wall thickness (WT),radial lumen diameter (R_D1),radial central diameter (R_D2),the ratio of cell wall thickness and radial lumen diameter (WT/R_D1),the ratio of average tracheid length and radial central diameter (TL/R_D2),Latitude,Longitude,Elevation,annual average rainfall (Rainfall),annual average temperature (Temperature),annual average sunshine (Sunshine),frost-free period (Period)
Piceapungens was the second tallest among the tested species.The average height ofP.pungensin Coconino County,Arizona,US,which has a semi-arid climate,was only 1.43 m at age 13 (Townsend and Douglass 1990).This is 26% shorter than the average height ofP.pungensat age 9 in this study.Of the native species,onlyP.linzhiensis,which originates in the tropical humid and sub-humid climate of Linzhi,Tibet,andP.likiangensis,which originates in the cold temperate monsoon climate of Shangri-La,Yunnan,grew relatively better than the other spruce from China.Thus,if the trees’ native regions are humid with high precipitation,spruce may show favorable growth on this site in Gansu province.
Relationship of trait performance with geographic factors
The wide range ofPiceaspecies may also trigger their adaptability to different climates;thus,species with a broad geographical distribution may have better growth performance.The natural distribution ofP.abiescovers 31 degrees of latitude,spanning from the Balkan Peninsula (latitude 41° 27′ N) to its northernmost extension near the Chatanga River,Siberia (latitude 72° 15′ N).The longitudinal range is from 5 °27′ E in the French Alps to 154° E at the Sea of Okhotsk in eastern Siberia (Schmidt-Vogt 1977).Altitudinally,it ranges from sea level to above 2300 m in the Italian Alps.Piceaspp.have been widely planted outside of this range,particularly in central Europe (Ellenberg and Ellenberg 1988) and Scandinavia.Similar toP.abies,P.glaucaandP.marianahave a transcontinental distribution and form the backbone of the forest products industry in boreal North America (Lenz et al.2011).Species with limited distribution grow relatively slowly,especiallyP.hirtella,a variety ofP.likiangensisvar.hirtella (Rehd.et Wils.) Cheng ex Chen,which is mainly found west of Sichuan and east of Tibet at altitudes of 3000–4000 m a.s.l.(Fu and Zheng 1978).Piceahirtella grew slowly in this study,and was 40%–60%smaller than the average height of all species.Similarly,P.schrenkianaalso grew slowly.Piceaschrenkiana is one of the most important zonal species in the arid lands of the Xinjiang Uygur Autonomous Region in northwestern China,and is distributed only in valleys and on humid slopes at altitudes of 2200–3500 m a.s.l.(Fu and Zheng 1978).Because it originates in high altitudes,it may show poor growth at low altitudes.P.schrenkiana(Altai Mountains of Tianshan),P.hirtella,P.spinulosa(Himalayas),andP.retroflexa(high mountainous areas of southwestern Sichuan Province) are found at altitudes of 2000–4000 m a.s.l.,which is higher than the study site (1160 m a.s.l.).This may explain the poor growth of the four species.In addition,the annual average sunshine of the test site was lower than that of the seed sources forP.schrenkiana,P.spinulosa,andP.meyeri,which may be another reason for the weak performance of these species.
Phylogenetic background effect on growth performance
Phylogenetic background is also a factor affecting growth performance (Givnish 1987).Piceaspecies distribution and population structure have changed significantly in response to the severe climatic oscillations associated with arctic glacial cycles during the late Tertiary and the Quaternary (Qian and Ricklefs 2000;Petit et al.2003;Hewitt 2004).With the advance and retreat of ice sheets,Asia may have been a refugia for the genusPicea(Wright 1955;Nienstaedt and Teich 1972),which is a hypothesis for the origin and evolution of North American spruces.P.abiesmay exhibit better growth in China due to its prehistoric origin in East Asia(Sullivan et al.2017).From there,it migrated to Europe which is assumed to have been a refuge during the last Ice Age (Siberia and the Urals).An ecogeographic analysis of four spruce species,P.glauca,P.engelmannii,P.sitchensis,andP.pungens,showed that their speciation was allopatric(Egorov and Afonin 2018).In addition,recent studies in molecular genetics have revealed thatP.sitchensisandP.pungensare sister species to the closely related P.engelmannii andP.glauca.There is a high probability that these four species shared a common ancestor in the first half of the Neogene (Lockwood et al.2013).
Picearetroflexa,P.crassifolia,P.wilsonii,P.purpurea,P.likiangensis,P.balfouriana,P.spinulosa,P.linzhiensis,andP.hirtellaare distributed in the Qinghai-Tibetan Plateau (QTP) and its adjacent regions.However,the growth of these species showed significant differences when grown at our research site.Phylogenetic,phylogeographic,and ecological studies indicate that plant diversification occurred on the QTP through multiple mechanisms,such as allopatric speciation via geographic isolation,climatic oscillations and divergences,pollinator-mediated isolation,diploid hybridization and introgression,and allopolyploidy (Petit et al.2003).Adaptive evolutionary radiation resulted inPiceadiversifying on the QTP,which resulted in marked morphology and adaptability differences.However,divergences may be relatively recent;thus,there has not been sufficient time to accumulate genetic variation (Wen et al.2014).P.schrenkiana,P.farreri,P.likiangensis,P.smithiana,andP.spinulosaare classified in the same cluster clade.It was suggested that these species may have been widely distributed in western China,but today’s limited distribution may be due to the rapid uplift of the QTP (Ran 2007;Li et al.2010).
Genetics and environment interactions on growth performance
Plant growth is not only influenced by the environment but is also by genetics and the interactions between genetics and the environment.The link between branching characteristics and wood quality has been recognized for several spruce species and has been shown to be associated with light availability (Sattler et al.2014).The number of first lateral branches,the length of the top whorl of branches,and branch angles ofP.abiesandP.pungenswere greater than for those of the other species.In addition,correlation analysis showed that the number of first lateral branches and the length of the top whorl of branches were positively correlated with height (Fig.4).A previous study showed that heights ofP.abiesclones from mother trees with branch angles of 90° to 110° were taller than clones from mother trees with 20° branch angles (Raiskila et al.2006).These results indicate thatPiceatrees with larger branch angles should grow taller.In this study,the branch angles of the two fastest growingPiceaspp.was > 60°.The lateral branch angle may be strongly tied to light capture,which has important implications for plant productivity and competition.In addition,an association between photosynthesis and branch angle has been established.The TILLER ANGLE CONTROL 1 (TCA 1) gene is a target of photosynthetic signals and is partially required for changes in lateral branch angles that are driven by photosynthesis (Waite and Dardick 2018).Species with slow growth rates likeP.hirtellaandP.schrenkinanahad values 30% −60% less than the average of the 17 species for the number of first lateral branches and the length of the top whorl of branches.
Relationships between growth traits and wood tracheid traits at the interspecific level
In this study,fast-growing species had larger tracheid lengths and smaller WT/R_D1 ratios.An increased tracheid length and decreased tracheid radial lumen diameter ratios are generally favorable for pulping,as they facilitate improved tear-tensile strength of paper.Consequently,the positive correlation of tracheid length with height indicates that early selection can be made at the interspecific level based on height.Like branch characteristics,wood properties are under strict genetic control (Zhang 1998).Height and branch length exhibited positive correlations with most tracheid traits.Of the 17 species,P.abieswas the tallest and had the largest radial lumen diameter ratios and TL/T_D2 values.Aside from genetic factors,P.abieshas also demonstrated that variations in maximum density were due to disparities in the cell wall thickness of the last formed cells,which differ depending on summer weather(Yasue et al.2000).A previous study in the central part of the Ceskomoravská Upland showed that the tree-ring width ofP.abiesis positively correlated with summer precipitation and negatively with summer temperatures (Rybnícek et al.2012).Higher temperatures in late winter and early spring increased ring width,whereas high summer temperatures generally had a negative impact on tree-ring width during the growing season (Koprowski 2013).In the present study,ring width displayed significantly positive correlations with latitude,but strongly negative correlations with longitude.Sudo (1955) also demonstrated a close relationship between wood anatomical characteristics and their native ranges.ThePiceaspecies were divided into three groups,A,B,and C,based on their wood anatomical traits (spiral thickenings in tracheid,spiral thickenings in ray tracheids,and crystals in ray parenchyma).The species in group A have limited natural ranges in Asia,(e.g.,P.maximowiczii),while those in group C are found in Asia,North America,and Europe(Sudo 1955).Hence,the differences in wood anatomical characteristics among thePiceaspecies in the same environment were related to innate characteristics,which are the result of long-term adaptation to their environment.
Conclusions
At our study site,the introducedPiceasuch asP.abiesandP.pungensdemonstrated better growth than the native species,and provides useful information for the utilization of germplasm resources from the genus.Moreover,the results offer critical guidance for forest managers with respect to decision-making about reforestation in this part of China.Possible reasons for superior growth of the five introduced species include more efficient photosynthesis,higher endogenous auxin levels,and greater water/nutrient use efficiency,all of which should be examined in future studies.Furthermore,long-term measurements in multi-site tests are needed to determine whether these improved growth factors will continue.
杂志排行

Journal of Forestry Research的其它文章
- Flexible transparent wood enabled by epoxy resin and ethylene glycol diglycidyl ether
- Diversity and surge in abundance of native parasitoid communities prior to the onset of Torymus sinensis on the Asian chestnut gall wasp (Dryocosmus kuriphilus) in Slovenia,Croatia and Hungary
- Ozone disrupts the communication between plants and insects in urban and suburban areas:an updated insight on plant volatiles
- Testing visible ozone injury within aLight Exposed Sampling Site as aproxy for ozone risk assessment for European forests
- Logging and topographic effects on tree community structure and habitat associations in a tropical upland evergreen forest,Ghana
- Spatial pattern dynamics among co-dominant populations in early secondary forests in Southwest China