地涌金莲MlCYP734A6基因的克隆与表达分析
2021-07-11万友名刘雄芳张秀姣曹毓蓉李正红
安 静,万友名,马 宏,刘雄芳,张秀姣,曹毓蓉,李正红
(中国林业科学研究院资源昆虫研究所,云南 昆明 650224)
油菜素甾醇(BRs)是一类在植物生长发育过程中发挥重要调节作用的甾醇类化合物的总称,作为植物的第六大内源激素,BRs广泛存在于低等和高等植物中,尤其是被子植物中,而且存在于植物的各个组织器官中[1-2]。目前,从自然界分离鉴定出的BRs已经超过了70种,其中,被认为分布最广泛且活性最强的是油菜素内酯(BL),通常被人们用来指代BRs[3-4]。BRs在植物整个生长发育和环境适应过程中发挥着重要的作用,尤其调控着许多重要的农艺性状,如植物结构、开花时间、种子产量和抗逆性等[5-8]。微剂量的BRs就可以显著促进植物生长,但过量的BRs则抑制植物的生长发育,因此,维持和调节内源BRs水平对植物的最佳生长发育状态至关重要[9]。植物的生长发育主要受环境因素和各种植物激素的调控,已有多篇文献证明油菜素内酯与不同的植物激素如生长素、细胞分裂素、脱落酸、乙烯、赤霉素、茉莉酸、多胺、水杨酸等相互作用,引发植物代谢及其生长发育的各种反应[10]。对BRs的研究近10多年一直呈逐年上升的态势[11]。
由于BRs不能进行组织和器官间的长距离运输,所以BRs的生物合成和分解代谢是维持BRs在植物体内稳态含量的两个关键拮抗过程[12-13]。目前,几乎完整的BRs生物合成及代谢途径已经被解析出来[14-16]。BRs代谢通路上最关键的失活基因CYP734As已经在多种植物中被挖掘及研究[17-22]。CYP734As基因属于细胞色素P450酶家族,在BRs失活过程中发挥着关键作用,可以催化BL和栗甾酮(CS)的C26位置上发生羟基化,使BRs失活,从而下调植物体内活性BRs水平[8,23]。BAS1/CYP734A1是第一个被研究报道出来对BRs失活有明确羟基化作用的基因,在拟南芥(Arabidopsis thaliana(L.)Heynh.)中过表达AtCYP734A1可导致植物体内BRs含量下降,植株出现极度矮化、叶片紧缩的表型[17]。而后,该基因的同源基因在其他植物中被陆续研究报道,番茄(Solanum lycopersicumMill)中 的CYP734A7、棉 花(Gossypium hirsutumL.)中的PAG1、胡萝卜(Daucus carotaL.)中的DcBAS1以及水稻(Oryza sativaL.)中的CYP734As,都被研究证实了具有使BRs失活的功能[18-20,22]。内源活性BRs具有反馈机制,当含量多时会下调合成基因和上调代谢基因,CYP734As的上调表达通常是由于BRs的反馈调控引起的[24]。CYP734As基因还受到其他影响因子的正负调控,有报道称,BAS1基因的表达受到转录因子BZR1和生长素响应因子ARF7的反向调控,二者都与BAS1启动子中的相同motifs结合,ARF7可抑制BAS1的表达或促进BRs生物合成基因DWF4的表达,来增加内源性BRs含量[25]。
地涌金莲(Musella lasiocarpa(Franchet) C. Y.Wu ex H. W. Li.)是中国金沙江河谷流域的特有珍稀单种属植物,属于芭蕉科(Musoideae)地涌金莲属(Musella),具有优美奇特的莲座状花序及端正笔挺的株形,文化及观赏价值极高。本研究组率先找到了几处地涌金莲野生种群,从中发现了株高、苞片颜色、吸芽数量等性状的不同变异类型,并注册登记了5个地涌金莲新品种[26-27]。笔者从前期高杆和矮杆2个无性系的转录组数据库中发现,与BRs代谢相关的CYP734A6基因的表达水平在矮杆品系中显著高于高杆品系,由此推测,地涌金莲植株高矮性状可能与CYP734A6基因的差异表达导致BRs含量不同相关。为进一步探究地涌金莲中CYP734A6基因的相关信息及表达规律,本研究克隆得到了MlCYP734A6基因的cDNA序列全长,并对其编码蛋白的理化性质、结构特征和系统进化进行了分析,同时应用qRT-PCR分析了MlCYP734A6基因在不同地涌金莲类型及不同组织部位中的表达模式,并与油菜素内酯含量做了相关性分析,以期为探究MlCYP734A6基因在地涌金莲生长发育过程中的调控作用提供分子依据,对今后采用分子手段培育不同株形性状新品种也具参考价值。
1 材料与方法
1.1 材料
植物材料种植于中国林业科学研究院资源昆虫研究所位于云南省禄丰县的滇中高原试验站。以地涌金莲矮杆类型(RD05)和高杆类型(YN01)为试验材料,二者均为来自于单株的组培苗无性系。2019年5月,从定植3 a的2个无性系中各随机选择30株调查平均株高,测量数据(RD05: 21.50 ±4.31 cm, YN01: 54.74 ± 6.06 cm)显示二者存在极显著差异(p< 0.01);各随机选择3株长势健壮的初花期植株,分别取每株的苞片、花序轴(假茎中间的茎秆)、叶、吸芽芽点及根尖5个部位为样品,每个部位至少3 g,采集的样品立即放入液氮中,随后保存于-80°C冰箱中备用。
1.2 方法
1.2.1 总RNA提取与cDNA第一链合成 采用CTAB法[28]提取地涌金莲样品总RNA;采用1.0%琼脂凝胶电泳和NanoDrop2000微量分光光度计检测RNA质量;按照Fermentas公司的反转录试剂盒VRevertAid First Strand cDNA Synthesis Kit说明书中的步骤合成cDNA第一链。
1.2.2MlCYP734A6基因的克隆 根据本研究组前期地涌金莲转录组信息(材料来源与本研究相同),在油菜素内酯代谢通路上筛选到1个注释为CYP734A6的显著差异基因片段,采用Primer Premier 5.0软件设计2条特异性扩增引物QA029F2和QA029R1(表1)。以RD05的5个部位材料等量混合提取RNA后反转录合成的cDNA为模板进行PCR扩增,PCR扩增体系为:2 × PCR Buffer for KOD FX Neo 25.0 μL,dNTP Mix (10 mmol·L-1) 1.0 μL,KOD FX Neo (1 U·μL-1)1.0 μL,cDNA第一链5.0 μL,ddH2O 15.0 μL,正反引物各1.5 μL。PCR扩增程序为:98℃预变性5 min;98℃变性10 s,55℃退火30 s,72℃延伸2.5 min,35个循环;72℃延伸10 min。

表1 引物序列Table 1 Primer sequences used in this study
PCR扩增产物经1.0%琼脂凝胶电泳检测合格,回收目的条带,使用TaKaRa公司的T4连接酶试剂盒进行连接后,4℃过夜,采用感受态细胞进行转化,获得阳性克隆后交由上海生工生物工程有限公司进行测序。
1.2.3MlCYP734A6基因的生物信息学分析 应用NCBI网站中的ORF finder和CDD功能查找克隆获得的全长序列的开放阅读框和保守结构域;应用Expasy网站(https://www.expasy.org/)中的ProtParam和Protscale在线软件分析蛋白的理化性质和亲疏水性;应用DTU.dk网站(https://services.healthtech.dtu.dk/)中的SignaIP5.0、NetPhos3.1和TMHMM2.0在线软件预测蛋白的信号肽、磷酸化位点和跨膜结构;使用BUSCA(http://busca.biocomp.unibo.it/)和SOPMA(https://npsa-prabi.ibcp.fr/NPSA/npsa_sopma.html)在线软件预测蛋白的亚细胞定位和蛋白的二级结构模型;应用DNAMAN8.0软件进行氨基酸序列比对;应用MEGA6.0软件的邻接法(Neighbor-joining)进行系统进化分析,以上在线分析软件的设置系数皆为系统默认值。
1.2.4MlCYP734A6基因的实时荧光定量分析 根据地涌金莲MlCYP734A6基因全长序列设计出1对荧光定量特异性引物AJ17F和AJ17R,扩增产物长度为146 bp,以地涌金莲TUBA为内参基因,设计内参引物TUBAF和TUBAR,扩增产物长度为190 bp(表1)。分别以地涌金莲RD05和YN01的苞片、花序轴、叶、吸芽芽点及根尖部位的cDNA为模板,对MlCYP734A6进行定量表达分析。仪器为BIO-RAD CFX ConnectTM荧光定量PCR检测系统,采用Power SYBR®Green PCR Master Mix (Applied Biosystems®Cat: 4 367 659)试剂盒,反应体系为20 μL,反应程序为:95.0℃预变性3 min;95.0℃ 10 s,55.0℃ 20 s,72.0℃ 20 s,75.0℃ 5 s,40个循环。每个样品设置3次重复,采用2-ΔΔCT方法进行数据分析,利用SPSS软件中的ANONA模块进行Duncan多重比较法进行数据的显著性差异分析。
1.2.5 油菜素内酯的提取与检测 取地涌金莲RD05和YN01各3株的苞片、花序轴、叶、吸芽芽点及根尖5个部位为油菜素内酯检测材料,每个部位至少3 g,交由南京卡文思检测技术有限公司采用高效液相色谱-质谱联用方法(HPLC-MS/MS)进行BL含量测定,每个样品重复3次。检测浓度带入公式:BL含量(ng·g-1)= 检测浓度(ng·mL-1)× 体积系数(mL)/质量系数(g),式中体积系数为0.20 mL,质量系数为样品实际称取质量。应用SPSS软件中的双变量相关性计算方法,计算皮尔逊相关系数r值和显著性P值,对MlCYP734A6及油菜素内酯含量进行相关性分析。
2 结果与分析
2.1 地涌金莲MlCYP734A6基因的克隆及生物信息学分析
以地涌金莲RD05的cDNA为模版,应用特异性引物对目的基因进行PCR扩增,电泳结果显示:在位于1 000~2 000 bp间有1条与预期大小相符的条带,经回收、测序后,显示该片段实际大小为1 639 bp(图1)。结合3′和5′ RACE技术,拼接出地涌金莲CYP734A6的基因序列全长,命名为MlCYP734A6(GenBank登录号MW013148)。该基因全长为1 936 bp,其中,5′非编码区长度为131 bp,3′非编码区长度为221 bp,包括长度为118 bp的polyA尾巴(图2)。

图1 MlCYP734A6的PCR扩增产物Fig.1 PCR amplification of MlCYP734A6

图2 地涌金莲MlCYP734A6基因的核苷酸序列及编码的氨基酸序列Fig.2 cDNA sequence and amino acid sequence coded of MlCYP734A6
MlCYP734A6包含1个长度为1 584 bp的完整开放阅读框,编码527个氨基酸,存在1个细胞色素P450蛋白家族结构域。TMHMM在线预测该蛋白在7~29氨基酸残基位置存有1个螺旋跨膜区域;SignalP5.0在线预测无信号肽位点,属于非分泌性蛋白质;BUSCA在线预测蛋白亚细胞定位于细胞质膜;蛋白二级结构预测结果显示,该蛋白包含51.04%的α-螺旋、12.14%的延长链、5.31%的β-折叠和31.50%的无规则卷曲(图3)。理化性质分析结果推测出,MlCYP734A6蛋白的分子式为C2728H4287N751O737S20,相对分子量为60 038.84 Da,等电点为9.44,酸性氨基酸残基总数(Asp+Glu)为54,碱性氨基酸残基总数(Arg+Lys)为67,稳定性系数为44.32,属于不稳定蛋白,脂肪系数为94.00;Protscale在线分析结果显示,总平均亲水性(GRAVY)为-0.111,推测MlCYP734A6为亲水性蛋白(图4A);磷酸化位点预测结果显示,磷酸化位点有40处,其中,丝氨酸19处、苏氨酸17处、络氨酸4处(图4B)。

图3 MlCYP734A6蛋白的二级结构预测Fig.3 Secondary structure prediction of MlCYP734A6

图4 MlCYP734A6的亲/疏水性分析和磷酸化位点分析Fig.4 Hydrophilicity/hydrophobicity analysis and phosphorylation sites analysis of MlCYP734A6
2.2 氨基酸序列比对及系统进化分析
将地涌金莲MlCYP734A6编码的氨基酸序列与其他植物进行比对,发现与多种植物的相似性达到75%以上,其中,与小果野蕉亚种(Musa acuminatasubsp.malaccensis)的相似性最高,达到94.69%,其次是凤梨(Ananas comosus(L.) Merr.)和油棕(Elaeis guineensisJacq.),分别为73.66%和73.33%。应用DNAMAN8.0软件将MlCYP734A6氨基酸序列与相似度比较高的几种植物进行比对分析,显示这几种植物均存在1个结构相似的细胞色素P450蛋白家族结构域(图5)。

图5 地涌金莲与其他植物的CYP734A6氨基酸序列比对Fig.5 The alignment of amino acid sequences of CYP734A6 in M. lasiocarpa and other plants
为了分析CYP734A6在不同植物间的亲缘关系,应用MEGA6.0软件,采用Neighbor-joining方法将地涌金莲MlCYP734A6与其他14种植物的同源蛋白进行系统进化分析,聚类结果显示,地涌金莲MlCYP734A6与小果野蕉亚种和油棕的同源蛋白遗传关系最近,而与黍(Panicum halliVasey.)、谷子(Setaria italica(L.) Beauv.)等单子叶植物遗传关系较远(图6)。
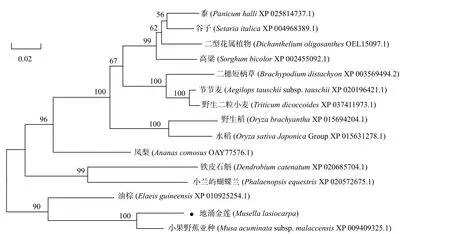
图6 地涌金莲MlCYP734A6与其同源序列的系统进化分析Fig.6 Phylogenetic analysis of MlCYP734A6 of M. lasiocarpa and itshomologous
2.3 MlCYP734A6基因的表达模式分析
运用实时荧光定量PCR检测MlCYP734A6基因在不同地涌金莲类型和不同组织部位中的相对表达量,结果(图7)表明:MlCYP734A6基因在2种类型地涌金莲的所有组织中均有表达且相对表达量差异显著(p< 0.05),矮杆类型地涌金莲RD05的花序轴中MlCYP734A6的相对表达量最高,为15.722,其次是根尖、苞片和吸芽芽点,最低的部位是叶片,仅为0.907;高杆类型YN01中,根尖中的相对表达量最高,为2.589,其次是苞片、花序轴、芽点,最低的部位也是叶片,仅为0.444。2种类型地涌金莲同一组织部位的比较分析结果显示,RD05在各个组织部位的相对表达量皆高于YN01,RD05的花序轴、根尖、苞片、吸芽芽点和叶片的相对表达量分别是YN01的14.13、5.08、4.47、1.28、2.04倍。

图7 MlCYP734A6在不同类型地涌金莲各组织中的相对表达量Fig.7 Relative expression of MlCYP734A6in different tissuesof M. lasiocarpa
2.4 油菜素内酯含量测定及与MlCYP734A6基因表达量水平的相关性分析
为了分析MlCYP734A6表达水平与油菜素内酯含量的相关性及地涌金莲株高与油菜素内酯的关系,笔者采用HPLC-MS/MS分别测定了2个类型地涌金莲5个组织部位的BL含量,检测结果(图8)显示:地涌金莲的5个组织部位皆能检测出BL,但不同组织中BL含量差异显著。在YN01中,叶片中的BL含量最高,为0.032 ng·g-1,其次是根尖、芽点、苞片和花序轴,分别是0.014、0.009、0.006、0.003 ng·g-1;在RD05中,苞片中的BL含量最高,为0.067 ng·g-1,其次是花序轴、芽点、叶片和根尖,分别是0.056、0.035、0.008、0.007 ng·g-1。结合MlCYP734A6的表达模式分析,YN01中的MlCYP734A6基因在叶中的表达水平最低,但BL含量却最高,而MlCYP734A6在花序轴的表达量较高,但BL含量却最低;在RD05中,花序轴中MlCYP734A6表达水平最高,但BL含量也较高,叶片中MlCYP734A6表达水平最低,BL含量也非常低。通过SPSS软件对不同组织中MlCYP734A6基因表达水平和BL的相关性进行分析,计算结果(YN01:r值0.568,p值0.318;RD05:r值-0.212,p值0.732)显示,二者之间不存在显著相关性。
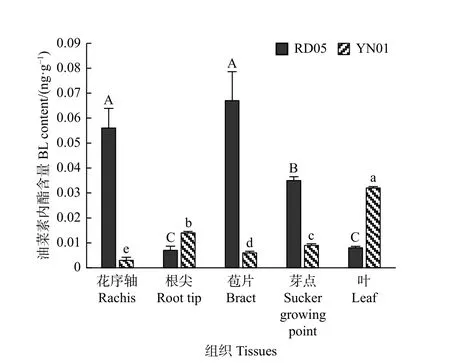
图8 不同类型地涌金莲不同组织中的BL含量Fig.8 The content of BL in different tissues of M. lasiocarpa
3 讨论
本研究成功地从地涌金莲中克隆到1个油菜素内酯代谢通路上的失活基因MlCYP734A6,通过多序列比对及系统进化分析,结果显示该基因编码的MlCYP734A6蛋白与同为芭蕉科的小果野蕉亚种的同源蛋白具有94.69%的高相似性,并且与其他植物的CYP734As进行比对,显示都具有细胞色素P450蛋白家族结构域,说明CYP734As在生物演变过程中是高度保守的,即MlCYP734A6可能具有与其他CYP734As类似的使BRs羟基化的功能。
对地涌金莲MlCYP734A6基因在不同株高类型地涌金莲中的表达模式分析发现,MlCYP734A6在地涌金莲不同组织部位的表达水平差异显著,2种类型地涌金莲分别在花序轴和根尖中的表达水平最高,这一结果与马铃薯StCYP734A1和水稻CYP734A6的表达模式类似,StCYP734A1在马铃薯的根中的表达水平最高,其次是茎,而水稻CYP734A6也是在茎节中的表达水平最高[21,29]。MlCYP734A6在地涌金莲中的这种表达模式与棉花中的PAG1基因的表达模式相反,在地涌金莲中,MlCYP734A6在花序轴和根尖中的表达水平最高,而在叶中的表达水平最低,而在棉花中,PAG1在根部和茎部的表达水平最低,在叶片中的表达水平最高[18]。由此可见,CYP734As基因在不同植物中的表达模式不尽相同,可能是由于BRs在不同植物的不同组织器官中行使着不同的功能,所以导致调控BRs含量的代谢基因CYP734As的表达模式差异很大。
分析HPLC-MS/MS检测结果可知,BL在地涌金莲各个组织器官中均可检测到,且含量差异显著,但与MlCYP734A6表达水平无显著相关性。在YN01中,叶中的MlCYP734A6表达水平最低,而BL含量却最高,花序轴中MlCYP734A6的表达水平较高,而BL含量却最低,说明MlCYP734A6可能在YN01的花序轴的BRs代谢通路上起到重要的代谢调控作用,主效下调着BRs含量的积累;而在RD05中,苞片中的BL含量最高,而MlCYP734A6的表达水平有也较高,是YN01的14.13倍,推测苞片中BRs合成基因的表达水平可能比较高,与代谢基因MlCYP734A6共同调控着BRs含量水平,而且MlCYP734A6在RD05中的高表达水平可能也受到了高含量BRs的反馈上调调控[24]。
高杆类型YN01的平均株高是矮杆类型RD05的2.57倍,结合地涌金莲转录组数据,前期分析筛选出BRs生物合成代谢途径中有3个显著差异Unigene,分别注释为合成通路中的CYP90A1/CPD、CYP90B1/DWF4和代谢通路中的CYP734A6,而信号转导通路中没有显著差异基因,这3个Unigene均是在RD05中显著上调,其中,CYP734A6基因的表达量最高,显著差异性最大,结合RD05的矮杆表型,故初期推测RD05中的BL含量应较低,但HPLC-MS/MS检测结果与前期预测不符,检测结果显示,RD05花序轴中的BL含量是YN01的18.67倍。推测可能的原因是,虽然RD05中的代谢基因CYP734A6显著上调,可使BRs含量下降,但合成通路中2个合成基因的显著上调发挥更大的调控作用,导致合成量高于代谢量,在花序轴中积累了过量的BRs。由于BRs具有双向调控的功能,缺乏或过量都会抑制植株生长,但目前对于过量BRs抑制植物生长的分子机理还尚未解析[29],所以推测地涌金莲RD05的矮化表型,可能是由于过量的内源BRs所致。植物的整个生长发育过程中,激素间的协同互作发挥着重要作用[30-31]。当BRs含量较高时,可抑制赤霉素的合成,从而抑制植物生长[29]。ABA作为植物生长抑制激素,虽然BRs在某些植物生理反应的调控中与ABA存在拮抗关系,但当BRs含量升高时,可上调若干个ABA响应基因和ABA生物合成基因[32]。故不排除其他激素对地涌金莲株高上的影响。所以接下来可以对不同类型地涌金莲在不同生长阶段进行全激素含量检测分析,并结合转录组数据进行激素和调控基因间的交互分析,为后续利用分子生物学技术改良地涌金莲株形提供参考数据。
4 结论
本研究成功克隆了地涌金莲MlCYP734A6基因,并分析了其编码蛋白的序列结构及蛋白特性,明确了MlCYP734A6在地涌金莲中的表达模式,分析了油菜素内酯与株高的关系。本研究结果为后续应用转基因或RNAi技术等调控MlCYP734A6的表达来平衡地涌金莲中油菜素内酯含量,进而为地涌金莲的生长发育水平奠定了分子基础,也为研究CYP734As在其他植物中的调控作用提供理论数据。
