芦可替尼和地西他滨协同抑制TET2敲降的HEL细胞生长作用
2021-07-09付莉霞王颖韶肖方楠宋濬哲白姣姣
付莉霞,王颖韶,肖方楠,宋濬哲,白姣姣,周 圆,白 洁
[1.天津医科大学第二医院,天津 300211;2.中国医学科院血液病医院(中国医学科学院血液学研究所),实验血液学国家重点实验室,国家血液系统疾病临床医学研究中心,天津 300020]
经典的费城染色体阴性骨髓增殖性肿瘤(myeloproliferative neoplasms,MPN)是一组造血干细胞恶性克隆性疾病,主要包括真性红细胞增多症(polycythemia vera,PV)、原发性血小板增多症(essential thrombocythemia,ET)和原发性骨髓纤维化(primary myelofibrosis,PMF)。MPN可进展为骨髓纤维化(myelofibrosis,MF),甚至转化为急性髓系白血病(acute myeloid leukemia,AML)[1]。2005年,多个课题组发现MPN的造血细胞中存在JAK2V617F突变,在其发病机制和靶向治疗研究中有重要意义[2-3]。该突变导致JAK-STAT级联信号通路过度激活,从而引起红细胞、血小板或粒细胞的过度增殖[4]。
TET2属于α-草酸戊二酸依赖酶的家族,催化5-甲基胞嘧啶(5-mc)转化为5-羟甲基-胞嘧啶(5-hmc),通过改变基因的甲基化状态并在表观遗传水平调控基因转录,该突变是MPN发展过程最常见的突变之一[5]。Moran-Crusio等[6-7]通过小鼠模型研究证实了JAK2V617F伴TET2缺失可以促进髓系细胞增生,从而加速MPN的进程。此外,Iurlo等[8-9]通过分析MPN病人的基因突变与其预后的相关性发现TET2突变的存在会增加PV患者向AML转化的风险。
进展为AML的MPN患者生存极差,且常规的化疗药物缓解率低,患者耐受性差。DNA甲基转移酶抑制剂地西他滨和阿扎胞苷是加速/急性期MPN(accelerated or blast phase disease MPN,MPN-AP/BP)的主要治疗药物。其中,地西他滨除诱导G2/M期的细胞阻滞外,还可通过抑制DNA甲基转移酶来影响DNA的转录来杀伤肿瘤细胞。研究报道,伴有TET2突变的MDS/AML去甲基化治疗有效率较高。但地西他滨对TET2和JAK2V617F共突变肿瘤细胞的有效性,及芦可替尼是否协同地西他滨杀伤TET2和JAK2V617F共突变肿瘤细胞尚未明确。本文在携带JAK2V617F的HEL细胞中建立了TET2敲降模型,对地西他滨和芦可替尼的敏感性及其协同作用进行评价,为TET2和JAK2V617F共突变的MPN患者提供临床治疗的理论依据。
1 材料与方法
1.1 药物、主要试剂及仪器地西他滨购自MCE(批号17685);芦可替尼为诺华公司惠赠;细胞培养基DMEM(批号8119392)、RPMI 1640(批号8120354)和胎牛血清(fetal bovine serum, FBS)(批号04-001-1)均购自BI;CCK-8试剂盒(批号04-001-1)购自日本Dojindo Laboratories; TRIzol(批号279511)购自Invitrogen; ImpromⅡ反转试剂盒(批号0000134688)购自Promega; Real-time DNA聚合酶mixture(批号)(FastStart Univeral SYBR Green Master)购自Roche(批号46660900);无内毒素质粒小提中量试剂盒购自天根(批号U8915);流式细胞仪Canto II购自美国Becton Dickinson公司;荧光倒置显微镜TE2000-S购自日本Nikon 司;高速离心机(型号Avavti J-26 XP)购自美国Beckman Coulter公司;ImageQuant LAS 4000发光成像仪购自GE。
1.2 细胞株及培养条件293T细胞株,HEL细胞株均购自ATCC细胞库,由中国医学科学院血液病医院(中国医学科学院血液学研究所)细胞库保管。293T细胞培养于DMEM+10%FBS培养基中,HEL细胞悬浮培养于RPMI 1640+10% FBS中,细胞于37 ℃、5% CO2培养箱中培养,每1~2 d离心换液,293T细胞在 70%-90%融合后经0.125%胰蛋白酶消化后传代。
1.3 慢病毒包装及病毒滴度测定TET2和PIGPZ shRNA购自Dharmacon(GE Healthcare Dharmaco,美国)。shRNA的靶向序列是TATGAGTCTCGAAC TCGCT,靶向人TET2基因编码区(Coding sequence,CDS)2383-2401位点,位于外显子3(NCBI序列号:NM_001127208.3)。包装载体psPAX2、pCMV-VSV-G由英国帝国理工学院Ernesto Yague博士惠赠。载体骨架结构为Fig 1A,通过已建立的方法进行病毒包装和滴度测定[10]。
1.4 慢病毒感染HEL细胞取对数生长的HEL细胞加入24孔细胞培养板中(1×105/孔)。根据MOI值5-10计算加入病毒体积(感染细胞数×被感染细胞MOI值/浓缩后慢病毒滴度),最后用含10%FBS的RPMI 1640培养液定容至500 μL/孔。1 800 r·min-1,33 ℃,离心90 min,6 h后换液。d 2用相同方法做二次感染。感染48 h后,流式细胞仪分选GFP+细胞用作后续实验。
1.5 RNA提取及Real-time PCR取对数生长期的EV HEL和TET2-KD HEL细胞2×106,TRIzol法提取RNA后进行反转录,然后进行Real-time PCR,引物设计参照文献[11],反应体系为(10 μL):SYBR Green Master 5 μL,上下游引物各0.5 μL,Template cDNA 0.5 μL,ddH2O(RNase-free)4 μL。反应条件为:95 ℃ 10 min 1个循环,95 ℃ 15 s和60 ℃ 60 s共40-45个循环。根据2-△△Ct法分析数据。
1.6 细胞增殖检测取对数生长期的EV HEL和TET2-KD HEL细胞,用含有10% FBS的RPMI 1640调整浓度为8×107·L-1,接种于96孔板中,每孔90 μL,EV-HEL和TET2-KD HEL细胞各3个复孔。培养后每孔加入CCK-8 10 μL,37 ℃孵育4 h,轻轻震荡混匀。酶标仪检测450 nm光密度(OD)值,检测d 0、1、2、3、4、5和6细胞增殖情况。
1.7 72 h药物半数致死剂量(IC50)测定采用CCK-8测定细胞对芦可替尼和地西他滨等化疗药物的敏感性。取对数生长期的EV-HEL和TET2-KD HEL细胞,用含有10%FBS的RPMI 1640调整浓度为8×107·L-1,接种于96孔板中,每孔90 μL,分组加药,每个浓度设3个平行孔,处理组加不同浓度的药物,阴性对照组加等体积的药物溶剂,培养72 h后每孔加入CCK-8 10 μL,37 ℃孵育4 h,轻轻震荡混匀。酶标仪检测450 nm光密度(OD)值。按以下公式计算药物对细胞的生长抑制率。抑制率=(对照组OD值-处理组OD值)/对照组OD值[12]。
1.8 药物协同指数测定参考单药时的IC50值,芦可替尼选择0.5、1和2 μmol·L-1。地西他滨选择3.2、16 和80 nmol·L-1,进行单药抑制作用测定和各个浓度两两混合的抑制作用测定。参照测定药物IC50时的细胞浓度和铺板方式进行试验,每种浓度组合3个平行孔,设计3×3矩阵。加药处理72 h后,通过CCK-8检测细胞活性。分别计算每个浓度组合下对细胞的抑制率,CalcuSyn软件计算各个浓度组合下的药物CI值。CI值<1.0代表两种药物在该浓度组合下具有协同效应,CI值>1.0代表两种药物在该浓度组合下具有拮抗效应[13]。
1.9 细胞系克隆形成(colony formation cells, CFC)实验取对数生长期的EV-HEL和TET2-KD HEL细胞,用含有10%FBS的1640调整浓度为8×107·L-1后接种于24孔板中,每孔1 mL,根据72 h的IC50值分别用2 μmol·L-1的芦可替尼和30 nmol·L-1地西他滨分别处理EV-HEL、TET2-KD HEL1#和TET2-KD HEL2#24 h后,再次计数并调整细胞浓度为2×107·L-1,分别取30 μL加入到1.2 mL甲基纤维素半固体培养基H4230,补充300 μL RPMI 1640培养基使各处理组体积相同,涡旋混匀,4 ℃冰箱静置10 min至气泡升至液面上部。用1 mL注射器(20 mL的枪尖)将混匀的细胞加入24孔板中,每孔0.4 mL(约200个细胞),每个处理组3个重复孔。于37 ℃,5% CO2,湿度>95%的培养箱中培养,并用无菌的PBS填充其余的孔以防半固体培养基干燥,14 d后计数不同的大小集落形成单位。
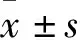
2 结果
2.1 建立TET2敲降的HEL细胞系HEL细胞是携带JAK2V617F人红白细胞白血病细胞系,常用作MPN研究。为了探究芦可替尼和地西他滨对JAK2V617F和TET2双突变细胞的影响,我们在体外利用shRNA在HEL细胞中敲降TET2(TET2-KD HEL)。首先,我们将TET2 shRNA和空载质粒进行了慢病毒包装,并分别感染HEL细胞。EV HEL作为对照组,TET2-KD HEL1#和TET2-KD HEL2#作为实验组。接着我们通过流式细胞术评价感染效率,EV-HEL、TET2-KD HEL1#和TET2-KD HEL2#感染效率分别为97.8%、88.4%和88%(Fig 1B)。因此我们进行了流式分选并在分选后再次通过流式细胞术进行阳性率评估,EV-HEL、TET2-KD HEL1#和TET2-KD HEL2#阳性率分别为99.6%、100%和99.9%(Fig 1C)。接下来,我们利用Real-time PCR对TET2敲降效率进行了评估,发现敲降组TET2的表达下降了35%-40%(Fig 1D)。提示成功建立EV HEL和TET2-KD HEL细胞系。
2.2TET2敲降后HEL细胞增殖能力增加有小鼠模型研究表明,在JAK2V617F和TET2敲降的双突变小鼠中髓系细胞细胞增殖能力增加[14]。为了研究TET2敲除对HEL细胞生物学功能的影响,我们通过CCK-8方法检测EV HEL和TET2-KD HEL细胞的增殖情况,通过检测d 0到d 6细胞的增殖情况,发现与EV HEL相比,TET2-KD HEL增殖能力明显增加(Fig 2,P<0.05)。
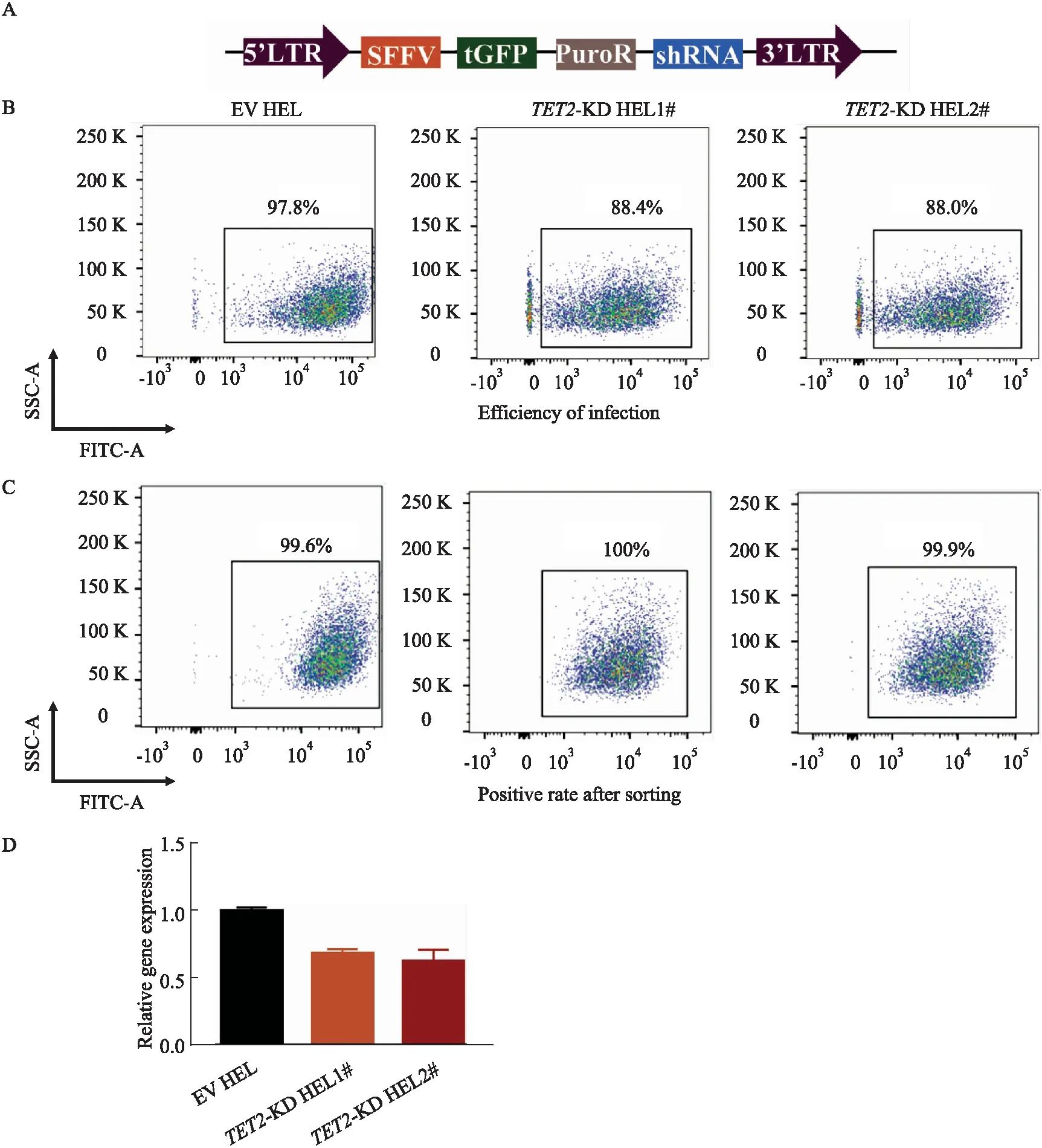
Fig 1 The positive rate and knockdown efficiency of HEL cells after lentivirus transfection

Fig 2 Proliferation of EV HEL, TET2-KD HEL1# and TET2-KD HEL2#
2.3TET2敲降后HEL细胞药物敏感性测定为了探索芦可替尼和地西他滨对TET2-KD HEL的抑制作用,我们通过药物处理72 h后的 IC50评价其对芦可替尼和地西他滨的敏感性。研究结果发现:芦可替尼在EV-HEL、TET2-KD HEL1#和TET2-KD HEL2#中IC50(72 h)差异无显著性,分别为(1.653±0.27)、(1.979±0.398)和(1.814±0.328 1)μmol·L-1(Fig 3A和3C,n=3)。但是,相较于EV-HEL而言,TET2-KD HEL1#和TET2-KD HEL2#对地西他滨的敏感性下降,其IC50(72 h)分别为(0.03±0.005)、(0.067±0.014)和(0.058±0.008)μmol·L-1,差异具有统计学意义(Fig 3B和3C,n=3,P<0.05)。接下来,我们分别用2 μmol·L-1芦可替尼和30 nmol·L-1地西他滨处理EV-HEL、TET2-KD HEL1#和TET2-KD HEL2#,并进行半固体克隆形成实验,结果显示TET2-KD HEL1#和TET2-KD HEL2#在芦可替尼和地西他滨单药处理后耐药克隆数均高于EV-HEL,且其中大克隆的数目明显增加(Fig 3D,n=3,P<0.05)。以上结果提示,虽然72 h药物敏感试验中TET2-KD HEL仍然对芦可替尼敏感,但是却对地西他滨存在耐药现象,且随着时间的增加,TET2-KD HEL对芦可替尼和地西他滨的药物敏感性均显著下降。
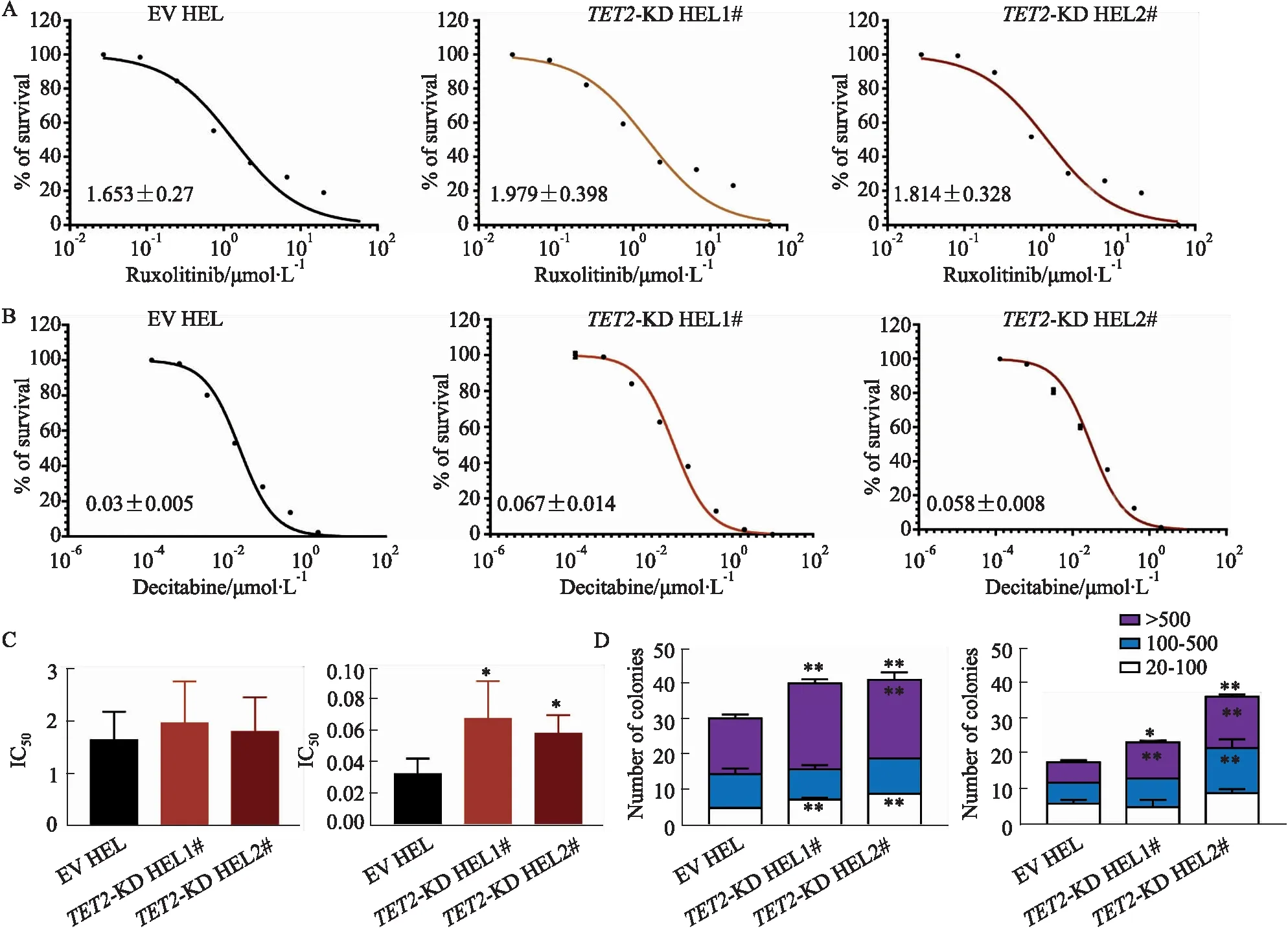
Fig 3 IC50 and drug-resistant colony numbers of ruxolitinib and dectabine on EV-HEL, TET2-KD HEL1# and TET2-KD HEL2# cells
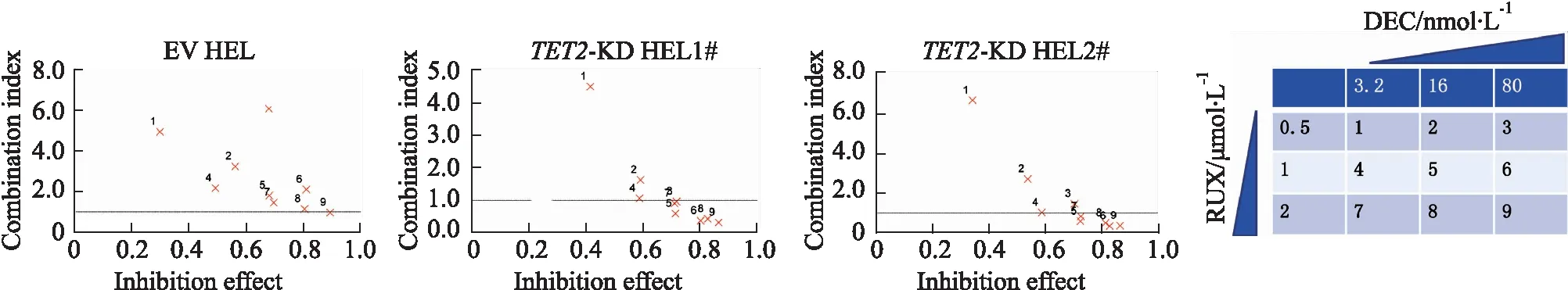
Fig 4 Combination index of ruxolitinib and decitabine
2.4 芦可替尼和地西他滨在TET2敲降HEL细胞中有协同抑制作用临床研究证实,在MPN-AP/BP患者联合芦可替尼和地西他滨,MPN-AP/BP耐受性较好[15]。考虑到TET2-KD HEL对地西他滨耐药,因此,我们在EV HEL和TET2-KD HEL中联合应用芦可替尼和地西他滨,观察其是否有协同作用。结果显示,与单独用药相比,这两种药物的组合对于EV-HEL细胞的生长无明显抑制作用,但在TET2-KD HEL1#和TET2-KD HEL2#细胞中,通过计算其抑制率并通过CalcuSyn软件分析其协同指数CI值,发现其在多个浓度组合下有明显协同效应,且芦可替尼1 μmol·L-1和地西他滨80 nmol·L-1联合用药对TET2-KD HEL1#和TET2-KD HEL2#抑制率分别可达到80%和82%。以上研究结果提示,对于TET2和JAK2V617F双突变的MPN患者而言,芦可替尼联合地西他滨可能是一种行之有效的用药方案。
3 讨论
JAK2V617F突变是MPN患者最常见的驱动型突变之一,该突变的发现在MPN患者机制研究和靶向治疗过程有里程碑意义,研究发现90%以上的PV患者有JAK2V617F突变,在ET和PMF患者中其突变频率也可达50%以上,该突变导致JAK激酶对细胞因子高度敏感或不依赖细胞因子仍能持续活化,从而激活下游的STAT3/5或PI3K/AKT等通路,促进造血细胞的过度增殖,并导致MPN的发生。研究表明,PV、ET和PMF10年内进展为AML的比率分别为2.3%-14.4%、0.7%-3%和10%-20%[16]。JAK1/2抑制剂芦可替尼可以明显改善MPN患者的体质性症状,缩小脾脏体积,并延缓脾大的发生,但芦可替尼单药治疗MPN-AP/BP患者还没有相关报导。
随着二代测序技术的发展,发现MPN患者中除了常见的JAK2V617F、CALR和MPL突变外,许多其它与表型和预后密切相关的基因突变也陆续被发现,其中包括TET2、ASXL1和DNMT3A等表观遗传学突变,TET2是其最常见的伴随突变之一。小鼠实验证实,TET2缺失的JAK2V617F突变小鼠表现为白细胞的增多、脾肿大、髓外造血及生存期缩短,且疾病进展迅速。而通过分析MPN病人的分子学突变与疾病发生和转归相关性,也发现TET2突变不仅参与MPN疾病的发生,还与疾病的进展及不良预后密切相关。因此,针对多种信号通路的联合用药可能会为改善MPN-AP/BP患者治疗现状、提高治疗效果提供可行的方案。
考虑到表观遗传学突变在MPN的发生与疾病进展中其重要作用,因此目前在MPN-AP/BP患者中使用芦可替尼联合表观遗传学相关的小分子药物也是临床研究的热点之一。DNA的去甲基化药物阿扎胞苷和地西他滨都可以调节DNA的去甲基化而影响其转录活性,但其对JAK2V617F和TET2双突变MPN患者具体作用尚未明确。
本研究通过建立TET2-KD HEL细胞系,发现相较于EV HEL,TET2-KD HEL细胞增殖能力增加,且TET2-KD HEL对地西他滨存在耐药现象。为了探索新的联合用药方案,我们将芦可替尼和地西他滨联合应用于EV HEL和TET2-KD HEL,发现该组合在TET2-KD HEL中有协同作用。有研究报道JAK2可以在酪氨酸1939和1946位点使TET2磷酸化,而磷酸化的TET2与类红细胞转录因子KLF1相互作用导致红系祖细胞的激活而促进MPN疾病的发生和进展。在MPN患者样本和小鼠模型中发现激活的JAK2V617F突变与TET活性和胞嘧啶羟甲基化的变化有关。这些表观遗传和功能变化也与一些致癌基因转录表达增加有关。且在MPN患者样本中发现JAK2V617F单突变的患者全基因组胞嘧啶甲基化显著降低,而在JAK2V617F和TET2并没发生该现象[17]。以上结果提示JAK2V617F和TET2双突变患者的甲基化异常状态可能是其对地西他滨耐药的主要原因。
总之,该研究证实TET2敲降后JAK2V617F阳性的HEL 细胞系对地西他滨耐药,芦可替尼和地西他滨在TET2-KD HEL有协同抑制作用。提示在TET2和JAK2V617F双突变MPN患者中联合应用芦可替尼和地西他滨可能是一种有效可行的治疗方案。
