滇楸优良无性系对水淹胁迫的生理响应及综合评价
2021-07-06杜克兵赵天宇黄国伟马林江张新叶
杜克兵,彭 婵,赵天宇,黄国伟,马林江,张新叶
(1.华中农业大学 a.园艺林学学院;b.湖北省林业信息技术研究中心,湖北 武汉 430070;2.湖北省林业科学研究院,湖北 武汉 430075;3.襄阳市林业科学研究所,湖北 襄阳 441052)
滇楸Catalpa fargesiiBur.f.duclouxii为紫葳科Bignonicaceae 梓树属的落叶乔木,是集用材、观赏和防风固沙等多种功能的优良树种,主要分布在我国云南、贵州、四川、湖南、湖北、广东等省[1]。因木材材质优良,滇楸被广泛应用于家具、建筑、工艺等行业[2]。湖北省江汉平原于2008年开始引种种植滇楸人工林,多年栽培试验表明,它们在江汉平原生长良好,可作为新型工业用材树种,目前已经选育出一批优良无性系[3-4]。同时,笔者也观察到滇楸对土壤水分过多十分敏感,地下水位较高以及季节性积水的地区植株长势弱,生长量小,甚至成片死亡[5]。研究滇楸在淹水逆境下的生理响应机制,并综合评价其抗涝能力具有重要的意义。滇楸和楸树为我国特有的乡土树种,抗逆性研究起步较晚,现在国内在楸树的抗涝性研究上取得了一些进展,但缺乏全面和系统性,而滇楸抗涝性则鲜有报道。本研究选取了湖北省林业科学研究院选育的4 个滇楸优良无性系和1 个楸树(C.bungei)优良无性系(作为滇楸抗涝性的对照)为材料,开展人工淹水胁迫试验,研究不同无性系在淹水胁迫下的生理响应,并采用16 项不同指标对其抗涝性进行了综合评价,以期为长江中下游季节性淹水地区滇楸优良抗涝无性系的选择、推广与应用提供理论依据。
1 材料与方法
1.1 试验材料
以湖北省林业科学研究院林业与园林研究所选育的优良无性系滇楸64#(简称DQ64)、滇楸67#(简称DQ67)、滇楸72#(简称DQ72)、滇楸78#(简称DQ78)和楸树2#(简称QS2)为试验材料,共5 个无性系。
1.2 植株培养
3月中旬,以5 个无性系(DQ64、DQ67、DQ72、DQ78、QS2)高3~4 cm 的生根试管苗为试材,移栽于装有营养土(山东商道生物科技股份有限公司,中国)的棕色塑料钵中(18 cm×14 cm×16 cm),并覆盖透明塑料杯保湿,每钵移栽1 株,钵底有12 个直径约1.2 cm 的排水孔。15 d 后移除覆盖的塑料杯自然生长。所有材料均于温室内进行移栽和培养。
1.3 淹水处理
5 个无性系的平均高度为40~50 cm 时,筛选生长状况良好且长势一致的单株开始淹水试验。全部植株随机分配到对照处理和淹水处理中:1)对照处理(CK),正常水分管理,土壤含水量为田间最大持水量的70%~75%;2)淹水处理,植株置于水箱中淹水,保持水面高出土壤表面10 cm。各个无性系每个处理的植株随机分为2 组:第一组用于表型、生长量、生理生态等对植株无损伤的指标的观测,植株采用完全随机区组排列,5 个区组,3 株/小区;第二组用于叶片与细根的采集,进行相关生理指标的测定,植株完全随机排列,9 株/无性系。淹水处理共计14 d。
1.4 测定方法
1.4.1 形态观察
试验期间,每天观察5 个无性系各个处理第一组植株的形态变化(包括叶片的颜色和数量、皮孔的膨大、不定根的产生等)以及死亡情况。
1.4.2 苗高与地径
试验的第0 天和第14 天,分别测定5 个无性系各个处理第一组植株的苗高和地径,计算试验期间苗木的生长量。
1.4.3 叶片气体交换、叶绿素荧光与叶绿素含量(SPAD 值)
试验的第0、7 和14 天,分别测定5 个无性系第一组植株的叶片气体交换参数、叶绿素荧光与叶绿素含量,5 株/处理。测定于9:00—11:00 晴朗无云天气进行,选取顶端以下第6 片完全展开的健康叶片测量。叶片气体交换参数采用Li-6400光合测定系统(Li-cor Inc.,Lincoln,NE,USA)测定,光照强度1 500 μmol·m−2·s−1,气体流速500 umol·s−1。测定内容包括净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、胞间CO2浓度/环境CO2浓度(Ci/Ca)等,潜在水分利用效率RWUEi=Pn/Gs[6]。采用Li-6400 荧光测定系统(Li-COR Inc.,Lincoln,NE,USA)测定叶绿素荧光,自然条件下叶片先暗适应20 min,然后测定其初始荧光(Fo)、可变荧光(Fv)和最大荧光(Fm)等参数。叶绿素含量采用SPAD-502plus 型便携式叶绿素测定仪(Konica Minolta Inc,Japan)测定SPAD 值。
1.4.4 生理指标:
试验的第14 天,分别采集5 个无性系第二组植株的叶片与细根,用于相对电导率(RMP)、丙二醛(MDA)、脯氨酸(Pro)和根系活力的测定。叶片采集植株顶端以下完全展开的第6~7 片叶,去除主脉;细根以蒸馏水快速洗净,滤纸吸干表面水分。全部样品用液氮速冻后,于-80℃冰箱中保存备用。RMP、MDA 含量、Pro 含量、根系活力的测定,参照王学奎等[7]的方法进行:采用3173 便携式电导率仪(Jenco Instruments,Inc,USA)测定叶片RMP;采用TBA 法(硫代巴比妥酸)测定叶片MDA 含量;采用酸性茚三酮比色法测定叶片与细根Pro 含量;采用TTC 法(2,3,5-三苯基氯化四氮唑)测定根系活力。
1.5 数据分析
图表中的显示数据均为平均值±标准误(Means±SE),其中叶片气体交换参数(Pn、Gs、Tr、RWUEi)、叶绿素荧光(Fv、Fv/Fo、Fv/Fm)与叶绿素含量(SPAD 值)的对照值均为其第0、7 和14 天对照处理的平均值。采用SAS 8.1统计软件进行方差分析(ANOVA)和多重比较(Duncan’s),方差分析前所有百分数均进行反正弦转换。淹水胁迫对植株造成的影响采用淹水变化率评价,淹水变化率=[(对照处理-淹水处理)/对照处理]×100%。
采用隶属函数值法对5 个无性系的抗涝性进行评价。分别计算每个无性系的Pn、Tr、Gs、Ci/Ca、RWUEi、Fv、Fv/Fo、Fv/Fm、SPAD、MDA、Pro、RMP、根系活力等指标在淹水第7 天和14 天的抗涝系数(=1-淹水变化率)的隶属函数值。如果分析指标与抗涝性呈正相关,则其计算公式为:

如果分析指标与抗涝性呈负相关,则计算公式为:
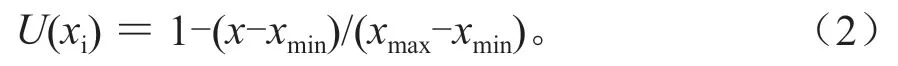
式(1)~(2)中:U(xi)是i品种某个指标的隶属函数值;x代表i无性系某个指标的均值;xmax是所有材料中某个指标的最大值;xmin是所有材料中某个指标的最小值。将每个无性系各指标的隶属函数值累加起来,求其平均值,均值越大则表明其抗涝性越强[8]。
2 结果与分析
2.1 植株形态
试验期间,5 个无性系的所有对照植株均未出现落叶和死亡现象,正常生长。至试验结束(第14 天)成活率均为100%。淹水处理下,5 个无性系植株的形态均发生了明显变化,包括叶片生长减缓、黄化和萎蔫,皮孔膨大等。5 个无性系均于淹水3~4 d 后开始出现茎基部皮孔膨大的现象,但试验期间均未出现不定根。DQ78 与QS2 于第4天开始出现叶片黄化现象,DQ64 和DQ67 的叶片黄化现象出现在第5 天,DQ72 出现在第6 天。叶片萎蔫出现的时间除DQ64 发生在第10 天之外,其余无性系均出现在第9 天。至试验结束时,5 个无性系的淹水植株均出现了不同程度的萎蔫现象,从外部形态看,差异并不显著。
2.2 生长量
淹水胁迫对5 个无性系苗高的生长都产生了显著抑制作用(P<0.05,图1),但仅对DQ78和QS2 地径的生长产生了显著抑制,其余3 个无性系的地径生长虽有降低,但与对照差异不显著(P>0.05)。至试验结束时,DQ64、DQ67、DQ72、DQ78 和QS2 苗高的淹水变化率分别为74.67%、77.28%、60.89%、78.74% 和60.91%,地径的淹水变化率分别为16.53%、21.18%、17.71%、24.73%和31.83%。可见,淹水处理对DQ72 苗高与地径生长的影响相对较小,对DQ67和DQ78 苗高与地径生长的影响相对较大。
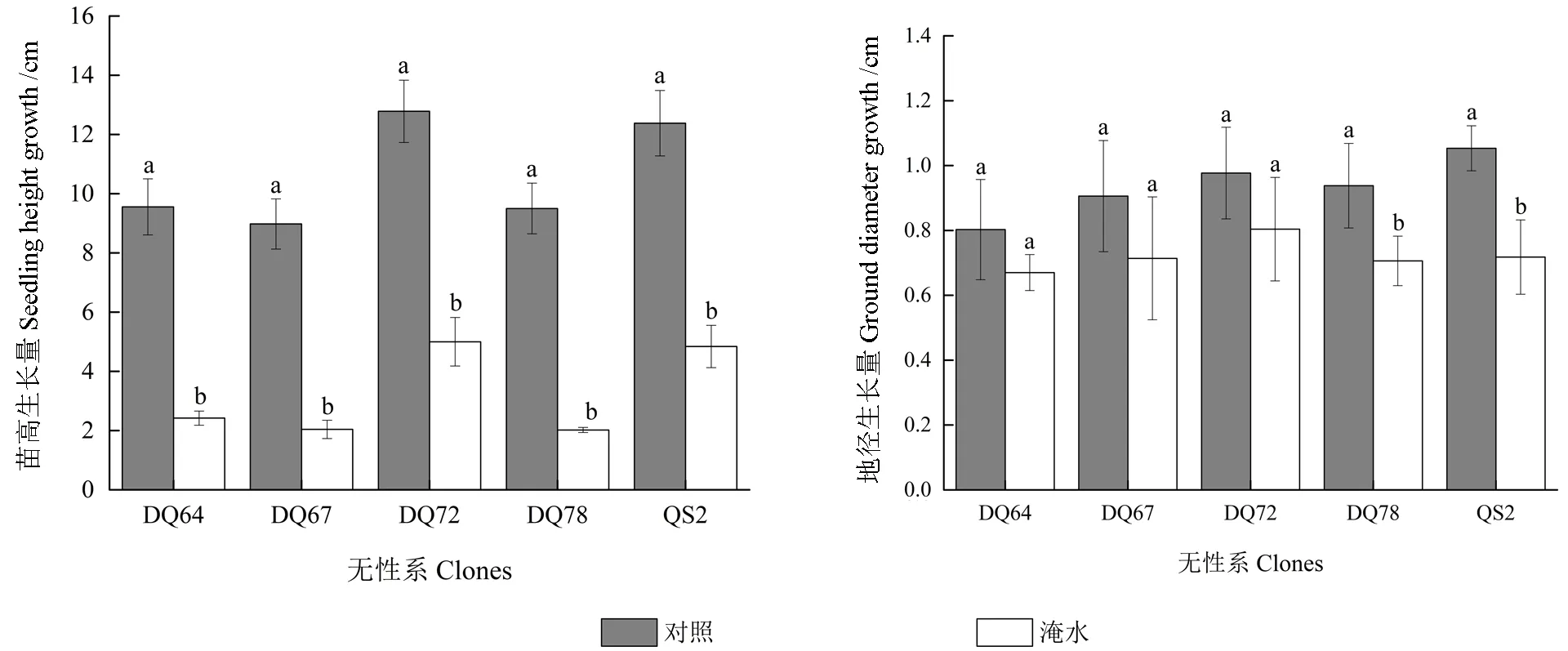
图1 淹水胁迫下5 个无性系苗高与地径生长量Fig.1 Height and root-collar diameter growth of the five clones under partial submergence treatment
2.3 叶片气体交换参数与叶绿素含量
淹水胁迫显著降低了5 个无性系叶片的Pn、Gs、Tr、RWUEi和叶绿素含量(SPAD 值),显著提高 了Ci/Ca(P<0.05,图2)。Pn、Gs和Tr在淹水7 d 时已经显著降低,14 d 时持续下降,但下降幅度较淹水7 d 时小。除DQ72 之外,多数无性系的RWUEi在淹水7 d 时下降幅度较小,甚至略有上升;淹水14 d 时,除DQ72 之外,其余4 个无性系的RWUEi均大幅降低。淹水7 d时,Ci/Ca的变化不显著,14 d 时大幅升高。SPAD 值自淹水处理开始持续下降,于第14 天时达到最小值,5 个无性系的变化趋势一致。淹水7 d 和14 d 时,5 个无性系的Pn、Gs、Tr、RWUEi和SPAD值淹水变化率的平均值见表1。5 个无性系各个指标的淹水变化率具有一定差异,其中DQ72 的SPAD 值变化较小,而Pn、Gs、Tr和RWUEi值的变化较大;DQ78 的SPAD 值变化较大,而Pn、Gs、Tr和RWUEi值的变化相对较小。

图2 淹水胁迫下5 个无性系的叶片气体交换参数与叶绿素含量Fig.2 Gas exchange parameters and chlorophyll contents in leaves of the five clones under partial submergence treatment
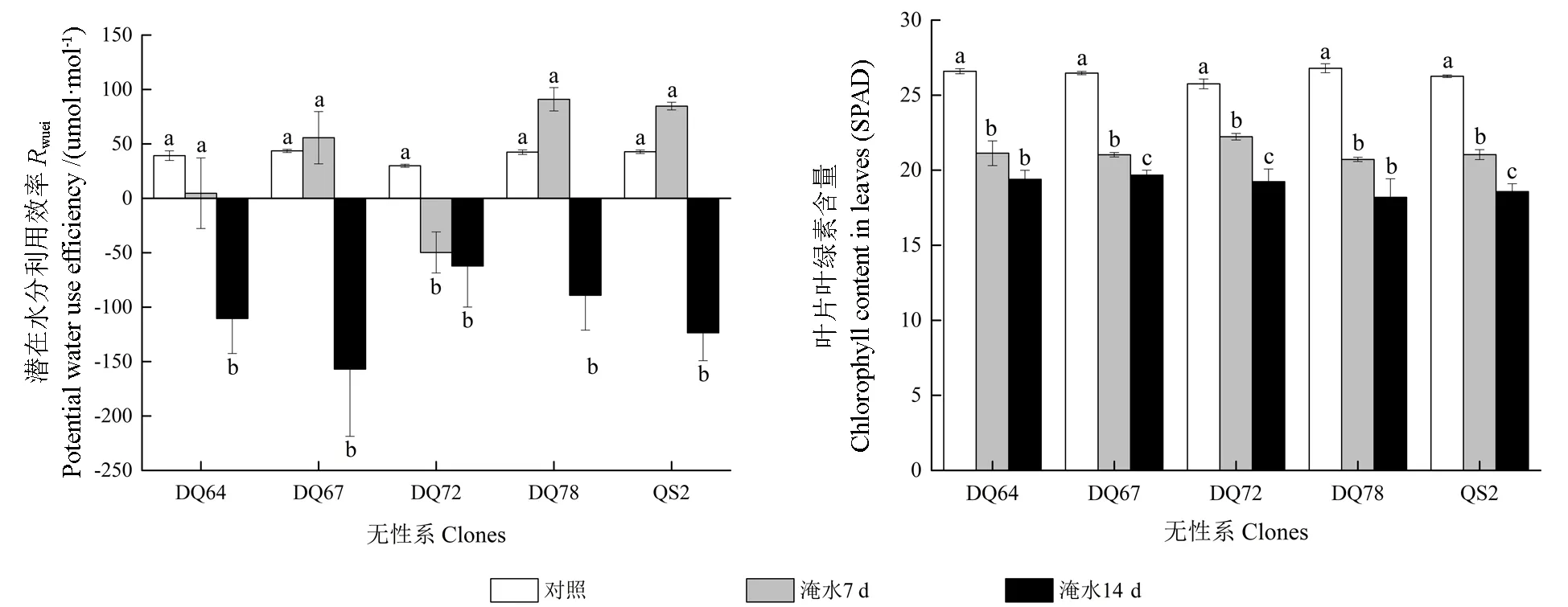
续图2Continuation of Fig.2
2.4 叶片叶绿素荧光
淹水胁迫显著降低了5 个无性系叶片的叶绿素荧光参数Fv、Fv/Fm和Fv/Fo(P<0.05,图3)。其中,Fv反映了光系统Ⅱ(PS Ⅱ)光化学活性的大小;Fv/Fm(PS Ⅱ最大光化学量子产量)反映了PS Ⅱ利用光能的能力和光抑制程度;Fv/Fo则反映了PS Ⅱ的潜在活性[9-12]。淹水7 d 时,除DQ72的Fv/Fo之外,5 个无性系的Fv、Fv/Fm和Fv/Fo均变化不显著,至14 d 时出现显著降低。淹水7 d和14 d 时,5 个无性系的Fv、Fv/Fm和Fv/Fo淹水变化率的平均值见表1。5 个无性系各个指标的淹水变化率具有一定差异,其中DQ72 的变化相对较大,DQ78 的变化相对较小。

图3 淹水胁迫下5 个无性系的叶片叶绿素荧光参数Fig.3 Chlorophyll fluorescence parameters in leaves of the five clones under partial submergence treatment
2.5 叶片相对电导率与MDA 含量
淹水胁迫显著提高了5 个无性系的叶片相对电导率(RMP)与MDA 含量(P<0.05,图4)。随着淹水时间的延长,RMP 值与MDA 含量在5个无性系中均持续增加,且对照、淹水7 d 和淹水14 d 的3 个时间点之间均存在显著差异。淹水7 d 时,DQ64、DQ67、DQ72、DQ78 和QS2的RMP 淹水变化率分别为-75.63%、-53.94%、-89.21%、-73.77%和-94.98%;淹水14 d 时,分别为-142.68%、-114.68%、-175.49%、-137.68%和-177.24%。对 于MDA 含量,DQ64、DQ67、DQ72、DQ78 和QS2 在淹水7 d 时的淹水变化率分别为-53.79%、-34.49%、-31.93%、-54.80% 和-49.82%;在淹水14 d 时的淹水变化率分别为-87.19%、-70.03%、-78.48%、-91.10% 和-103.47%。可见,淹水胁迫对DQ67 的RMP 的影响最小,对QS2 的影响最大。
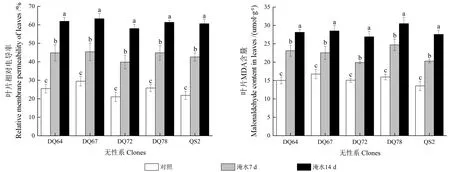
图4 淹水胁迫下5 个无性系的叶片相对电导率与MDA 含量Fig.4 Relative membrane permeability and malonaldehyde contents in leaves of the five clones under partial submergence treatment
2.6 叶片与细根的脯氨酸含量
淹水胁迫显著提高了5 个无性系的叶片与细根中的脯氨酸含量(P<0.05,图5)。细根中脯氨酸含量的变化趋势在5 个无性系中基本相同,随着淹水时间的延长逐渐升高。叶片中脯氨酸含量的变化趋势在不同无性系中略有不同。除DQ72 叶片中的脯氨酸含量随着淹水时间的延长逐渐升高之外,其余4 个无性系的叶片中脯氨酸的含量均呈现先升高后降低的趋势。淹水7 d时,DQ64、DQ67、DQ72、DQ78 和QS2 的 叶片中脯氨酸含量的淹水变化率分别为-88.89%、-69.45%、-46.40%、-115.43% 和-76.12%;淹水14 d时,分别为-25.08%、-31.18%、-76.95%、-42.34%和-28.29%。在细根中,DQ64、DQ67、DQ72、DQ78 和QS2 在淹水7 d 时脯氨酸含量的淹水变化率分别为-219.93%、-150.85%、-106.70%、-78.00%和-133.48%;淹水14 d 时,分别为-322.46%、-292.35%、-195.59%、-79.39%和-198.36%。可见,淹水胁迫下DQ78 的叶片中积累了较多的脯氨酸,DQ64 的细根中积累了较多的脯氨酸。
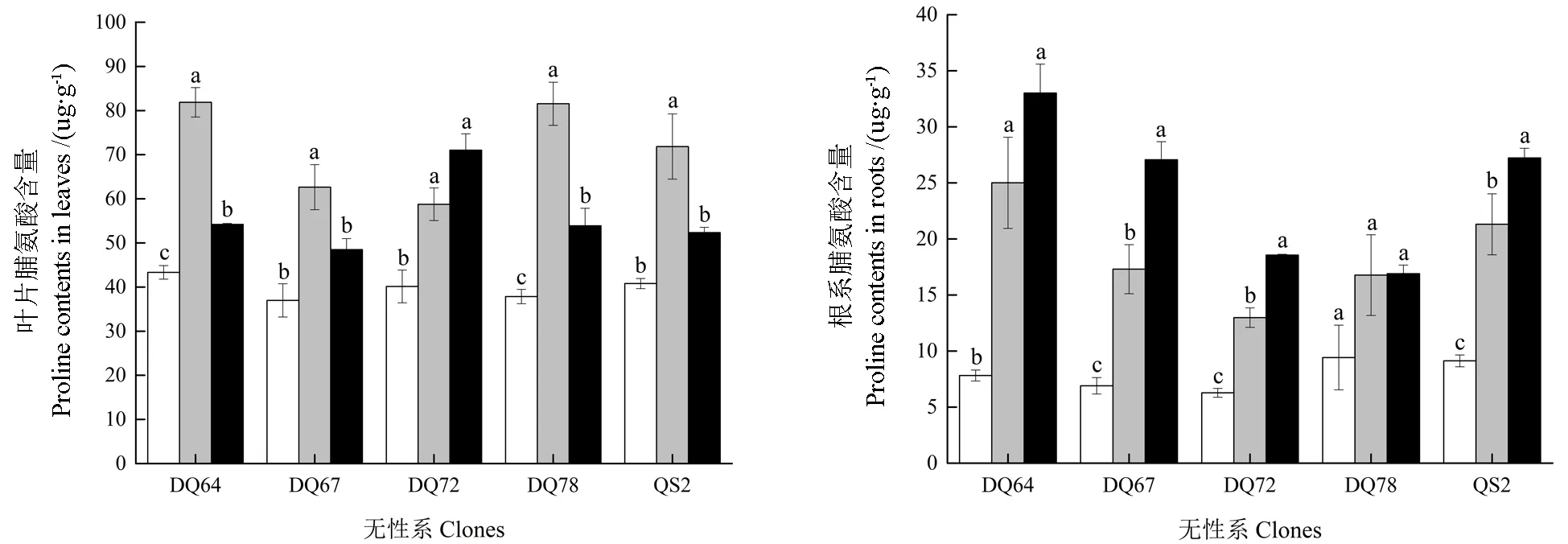
图5 淹水胁迫下5 个无性系的叶片与细根中的脯氨酸含量Fig.5 Proline contents in leaves of the five clones under partial submergence treatment
2.7 根系活力
淹水胁迫显著降低了5 个无性系的根系活力(P<0.05,图6)。随着淹水时间的延长,5 个无性系的根系活力均持续降低,于14 d 时达到最小 值。DQ64、DQ67、DQ72、DQ78 和QS2 在 淹水7 d 时根系活力的淹水变化率分别为28.39%、27.70%、20.07%、27.61% 和41.58%。淹 水14 d时的淹水变化率分别为55.19%、50.33%、42.51%、52.14%和59.78%。可见,淹水胁迫对DQ72 根系活力的影响最小,对QS2 的影响最大。
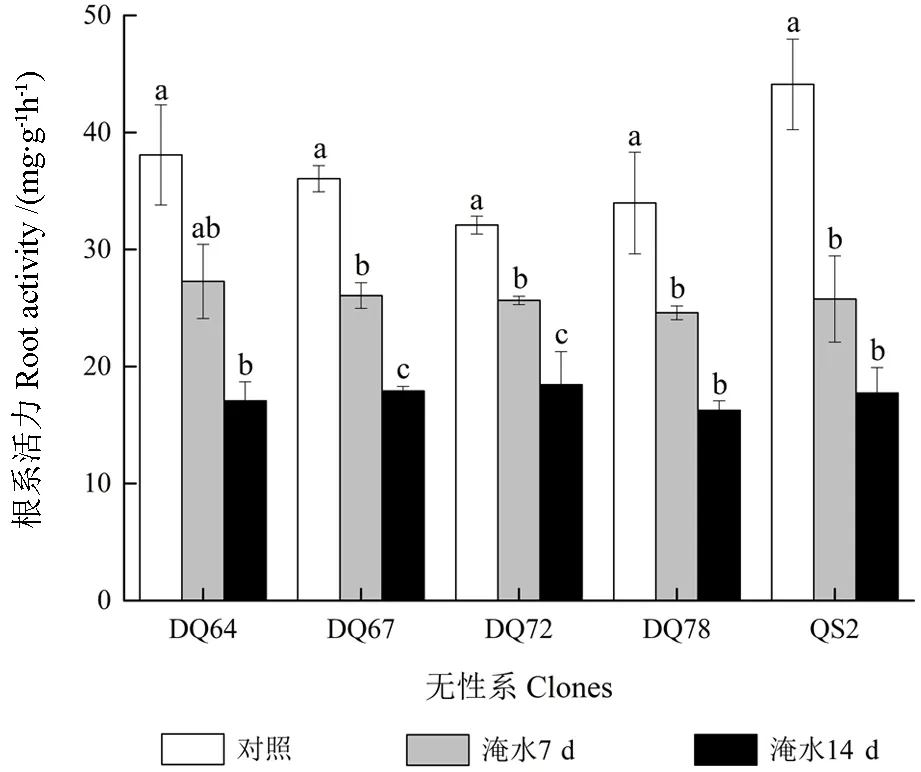
图6 淹水胁迫下5 个无性系的根系活力Fig.6 Root activity of the five clones under partial submergence treatment
2.8 隶属函数分析
淹水胁迫下5 个无性系抗涝指标的隶属函数值见表2。依据平均隶属函数值的大小,5 个无性系抗涝性的强弱排序为:DQ67>DQ78>DQ72>DQ64>QS2。滇楸4 个无性系的抗涝性差异不明显,但均强于QS2。

属隶值数均函Average membership value 0.476 7光荧The membership values of floodingresistance indices平荧始变可/初光Fv/Fo 0.236 4最学率PSⅡ化效光子大量Fv/Fm 0.290 4可荧变光Fv 0.418 5水效分率潜利RWUEi在用0.491 9胞浓CO2 浓/环CO2境度间度Ci/Ca 0.385 9†值数腾率Tr函蒸速0.633 7属隶的标孔度Gs气导指0.621 6涝抗合系率Pn光速性净0.310 4无5 个酸系氨下根脯Proline 1.000 0迫胁水值片酸2 淹数函叶氨Table 2Themembership values of floodingresistance indicesin thefiveclones underpartialsubmergencetreatment脯Proline 0.307 8属表隶标片指叶MDA 0.265 4涝抗力系Root活根metabolism 0.439 2电RMP率导0.511 9叶SPAD素绿0.486 4径地Ground diameter 1.000 0高苗Seedling height 0.227 9无Clones系性DQ64 0.530 1 0.500 0 0.500 0 0.500 0 0.386 4 0.345 4 0.514 5 0.583 9 0.325 2 0.694 7 0.225 8 0.944 1 0.596 1 1.000 0 0.587 3 0.696 1 0.081 8 DQ67 0.512 4 0.161 0 0.200 2 0.014 6 0.500 0 0.500 0 0.322 1 0.278 0 0.500 0 0.340 1 0.500 0 0.873 7 1.000 0 0.084 2 1.000 0 0.923 7 1.000 0 DQ72 0.527 5 0.712 7 0.719 6 0.856 6 0.993 5 0.792 1 0.500 0 0.500 0 0.930 2 0.000 0 0.666 4 0.185 0 0.545 6 0.574 4 0.000 0 0.463 7 0.000 0 DQ78 0.399 8 0.355 3 0.497 3 0.485 3 0.713 5 0.617 1 0.325 8 0.579 5 0.665 6 0.440 2 0.246 3 0.108 9 0.000 0 0.000 0 0.364 0 0.000 0 0.998 7 QS2。值均平的14d 时7 d 和水淹为均值数函属隶的标指余其,外径地与高苗除中† 表Exceptforseedling height andground diameter,subordinativefunction valueof otherindexesin thetablewere mean values of waterlogging for7days and14 days.
3 结论与讨论
3.1 结 论
淹水胁迫对5 个无性系的形态、苗高与地径生长、叶片的气体交换参数、叶绿素荧光、叶绿素含量、相对电导率、丙二醛含量、根系活力、叶片与细根的脯氨酸含量均产生了不同程度的影响,表明5 个无性系受到淹水胁迫的伤害有所不同。5 个无性系的抗涝性强弱排序为DQ67>DQ78>DQ72>DQ64>QS2。
3.2 讨 论
淹水胁迫对植物造成损伤的主要原因是土壤缺氧以及乙烯、CO2、锰和铁等元素的过量积累[13]。淹水胁迫会对植物的解剖结构、营养代谢、激素平衡、光合作用、呼吸作用等产生显著影响[14-15]。植物的抗涝性评价指标主要包括存活率、生长量、生物量、形态结构和生理代谢等[16-17]。
形态结构是反应植物受害程度最直接的指标,也是植物适应淹水胁迫最基础的要素。淹水胁迫下植物形态结构的变化主要包括叶片黄化、萎蔫与脱落、生长变缓、根系死亡、皮孔膨大、不定根诱导、根系通气组织形成等[18]。淹水胁迫下,皮孔膨大与不定根产生有助于植株从水体中获取O2、水分与营养元素,并释放C2H2、乙醇等有害物质,以减轻其毒害,适应长期的淹水胁迫[19]。本试验中,5 个无性系在淹水胁迫下均出现明显的叶片黄化(4~6 d)和萎蔫(9~10 d)等症状,有皮孔膨大的现象(3~4 d)但未形成不定根,难以适应长时间的淹水胁迫。5 个无性系之间虽有一定差异,但皮孔膨大、叶片黄化与萎蔫的时间等差异并不明显,仅相差1~2 d。淹水胁迫通过减缓叶片和根系生长、降低水分与矿质营养吸收、改变叶绿素含量和组分、破坏光合机构的结构与性能等,降低植物光合同化能力,进而抑制其生长和生物量积累,但高抗植物受影响的程度较小[13]。本试验中,淹水胁迫显著抑制了5 个无性系苗高(60.89%~78.74%)与地径(16.53%~31.83%)的生长,5个无性系之间受抑制的程度存在一定差异。
淹水胁迫会导致植物关闭气孔,降低光合作用,并提高水分利用效率,这有助于植物降低水分散失,维持水分平衡[20]。本试验中,5 个无性系的Pn、Gs和Tr均随着淹水时间的延长而逐渐降低,这与很多植物在淹水胁迫下的表现一致[11、21]。Gs的变化是导致Pn、Tr、Ci/Ca 和RWUEi等变化的直接原因[20,22]。Farquhar 等[23]认为,植物的光合抑制源自气孔限制和非气孔限制两种原因,若Pn、Gs和Ci/Ca 同时下降,则Pn降低主要受气孔限制;若Pn与Gs呈下降趋势,而Ci/Ca 却上升,则Pn降低主要受非气孔限制。本研究中,淹水7 d时DQ67、DQ78 和QS2 的Pn降低主要由气孔因素造成,因为气孔关闭和气孔导度下降,导致CO2供应不足,而DQ64 和DQ72 主要由非气孔因素造成;淹水14 d 时5 个无性系Pn的降低均主要由非气孔因素造成,因为叶肉细胞光合活性的受损致使同化力不足,这与杨树PopulusL.、巴西胡椒木Schinus terebinthifolius等的研究结果一致[9,22]。淹水7 d 时5 个无性系的Pn、Gs和Tr即大幅降低,淹水14 d 时RWUEi显著减小,说明滇楸与楸树对淹水胁迫均十分敏感,这与其叶片的表型变化一致。淹水变化率显示,5 个无性系叶片气体交换参数的变化幅度有所不同,表明其光合系统受损程度不同。
叶绿素荧光参数Fv、Fv/Fo和Fv/Fm是检测植物光化学反应状况的3 个重要参数,可准确揭示植物光系统对光能的吸收、传递、耗散和分配等情况[12,24]。已有报道表明,淹水胁迫下,这3 个参数均会不同程度降低[25]。本试验中,5 个无性系淹水植株的Fv、Fv/Fo和Fv/Fm均显著低于对照,表明淹水胁迫导致叶绿体光合机构受损,光合作用原初反应过程受抑制,光合CO2同化效率降低[12]。5 个无性系Fv、Fv/Fo和Fv/Fm在淹水7 d 时下降大多不显著,而在14 d 时大幅降低,说明淹水时间对其光合机构的影响很大,这可能与植株茎基部未能诱导形成不定根有关。淹水变化率显示,5 个无性系Fv、Fv/Fo和Fv/Fm的变化幅度有所不同,表明其光合系统的受损程度存在差异,这与其叶片气体交换参数的表现基本一致。
叶片MDA 含量与RMP 均为显示植物在逆境胁迫下受害程度的重要指标。MDA 是细胞膜膜脂过氧化作用的产物之一,淹水胁迫会导致膜脂过氧化反应加剧,MDA 含量升高,引起膜系统损伤,进而影响细胞膜透性,其含量能反映膜脂过氧化的程度,从而间接反映植物组织抗氧化的能力[26]。RMP 通过测定细胞内电解质外渗的程度,从而显示细胞膜结构的受损程度[27]。已有研究结果表明,淹水胁迫往往导致植物叶片MDA 含量与RMP 均升高[6,9]。本研究结果与已有报道一致。5 个无性系的叶片MDA 含量与RMP 均随着淹水时间的延长持续上升,但5 个无性系之间上升幅度略有不同,表明其受到淹水胁迫的伤害有差异。植物细胞中脯氨酸是一种重要的渗透调节物质。当植物遭受低温胁迫时,脯氨酸具有维持细胞渗透压,避免细胞过度失水的功能,还可以通过和蛋白质分子结合来稳定结构,使植物新陈代谢处于正常状态中,提高植物对逆境的适应性[28]。本试验中,5 个无性系淹水植株叶片与细根中的脯氨酸含量均显著高于对照,在叶片中呈现先升高后下降的趋势,这可能与淹水9~10 d 后叶片开始萎蔫有关。根系活力代表了植物根系吸收水分和矿质营养的能力,显示了植物在逆境胁迫下的活力与受害程度[29]。本研究中,5 个无性系的根系活力均随着淹水胁迫逐渐降低,与杨树等树种对淹水胁迫的响应一致[6]。淹水变化率显示,5 个无性系之间的变化幅度存在一定差异,表明其受淹水胁迫的程度有所不同。
采用16 个形态和生理生化指标对5 个无性系的隶属函数综合分析发现,4 个滇楸无性系的抗涝性存在一定差异,且均强于QS2,这与张烨然等[6]的研究结果相似。本研究结果可为了解滇楸与楸树的抗淹性提供参考,但由于本试验是在人工控制条件下进行的,且试验周期较短,还不足以完全展现5 个无性系在复杂多变的自然环境条件下对长期淹水胁迫的响应,所以各无性系的抗涝能力还有必要进一步进行田间试验加以检验。同时,滇楸和楸树对淹水胁迫敏感的原因尚不清楚,还需要进一步深入研究,以便为滇楸和楸树的抗涝性育种提供理论依据。
