川西亚高山2种典型针叶林土壤碳氮磷和碳稳定同位素分布特征
2021-07-02杨玉婷徐振锋
古 韬,王 华,刘 瑞,杨玉婷,谭 波,张 丽,徐振锋,李 晗
(四川农业大学生态林业研究所/长江上游林业生态工程四川省重点实验室/长江上游森林资源保育与生态安全国家林业和草原局重点实验室/高山森林生态定位研究站,成都 611130)
土壤碳(C)、氮(N)、磷(P)是陆地生态系统植被生长发育的重要元素,能够反映土壤碳蓄积动态及养分供应能力,是衡量森林生态系统物质循环和能量流动的重要指标之一[1-2]。土壤碳含量不仅能够影响土壤稳定性和持水性等结构性能,在植被养分供应、分析植被类型和土壤类型方面均具有重要地位[3]。氮是植物在生长过程中从土壤中吸收利用最多的养分元素,其各种转化过程是评价森林生态系统功能的重要指标[4],而土壤磷作为植物所需磷元素的唯一来源,不仅用于合成植物体结构成分,并且参与多种生化反应过程[5]。已有研究表明,不同森林群落的叶片、凋落物和土壤C、N、P含量差异显著,且受到纬度、海拔等因素的显著影响[6]。在土壤中存在的13C这一稳定形态的碳同位素[7],认识其变化特征在探究土壤碳来源和土壤有机质周转周期等方面具有重要的指示作用[8]。研究表明,土壤δ13C值与土壤有机碳含量随时间的变化动态存在对应关系,即以δ13C含量最高处为界限,时间越久,其上层土壤δ13C含量越高,下层土壤δ13C含量越低[9-10],且13C的分布特征能够用于探究从年到百年尺度的土壤碳循环过程[11-12]。因此,研究不同林型土壤不同层次碳氮磷含量及碳稳定同位素组成特征及其关联性有助于深入认识森林生态系统的碳和养分循环机制。
川西亚高山森林是我国第二大林区的重要组成部分,地处长江上游,在水土保持、水源涵养、气候调节和生物多样性保育等方面具有重要的地位[13-14]。目前,针对该区域已开展关于土壤酶活性特征、土壤微生物群落结构和林地土壤质量演变等研究[15-18],但该区域典型针叶林生态系统土壤不同层次碳氮磷含量的分布仍不清晰,尤其对碳稳定同位素组成特征鲜有报道。因此,本研究选取川西亚高山2种典型针叶林岷江冷杉(Abiesfaxoniana)林和粗枝云杉(Picea asperata)林为研究对象,探究不同土壤层次(新鲜凋落物层、半分解层、腐殖质层,土壤 0~5、5~10、10~15和15~20 cm层)碳、氮、磷含量及碳稳定同位素组成,分析不同林型和土壤层次对碳氮磷及碳稳定同位素组成分布特征的影响,揭示不同森林类型土壤碳氮磷与碳稳定同位素组成的关系,以期为深入认识川西亚高山针叶林生态系统的碳和养分循环过程提供科学数据。
1 材料与方法
1.1 研究区域概况
研究区域位于四川省阿坝理县毕棚沟高山森林生态系统定位研究站(102°53′~102°57′E,31°14′~31°19′N),地处青藏高原东缘与四川盆地的过渡地带,海拔2 458~4 619 m。区域内年平均气温2~4℃,最高温度23.7℃,最低温度-18.1℃,年降雨量约为850 mm。森林植被随海拔由低到高依次为针阔混交林、高寒针叶林、高山灌丛和高山草甸,乔木树种主要由岷江冷杉、粗枝云杉、方枝柏(Sabinasaltuaria)和四川红杉(Larixmastersian)、红桦(Betulaalbosinensi)等组成;灌木主要由红刺悬钩子(Rubusaurantiacus)、红脉忍冬(Lonicranervosa)、疏花槭(Acer laxiflorum)、紫花卫茅(Euonymusporphyreus)等组成;主要草本由柳叶菜(Epilobiumlaetum)、卵叶葱(Alliumovalifolium)、蟹甲草(Cacaliaroborowskii)和禾本科部分属种等组成[19]。该地区成土母质主要为灰岩、页岩、板岩等残积风化物,土壤为暗棕壤,土壤有机层厚度约12.5 cm,pH值约5.6。
1.2 样地设置和样品采集
于2018年9月在研究区域选择海拔3 000 m处的亚高山2种典型针叶林岷江冷杉林(192 a)和粗枝云杉林(30 a)为研究对象,胸径、树高和郁闭度分别为39.8 cm/11.4 cm、20 m/8 m、0.7/0.8。在每种林型内分别设置3个坡度、坡向、郁闭度相似的20 m×20 m样地,每个样地中选择5个1 m×1 m样方,挖取土壤剖面按层次采样。分为土壤有机层(新鲜凋落物层、半分解层和腐殖质层)和矿质层(0~5、5~10、10~15和15~20 cm)共7个层次,装入封口袋中,带回实验室,风干后储存备用。
1.3 室内分析
将风干土壤样品过2 mm筛,凋落物样品采用球磨机粉碎后过60目筛。全碳含量采用重铬酸钾法测定[20],全氮含量采用半微量凯氏定氮法测定[21],全磷含量采用钼锑钪比色法测定[22]。土壤和凋落物样品过200目筛后,加入过量1 mol/L HCl,利用振荡机振荡,使土壤与HCl充分接触,于室温下处理24 h,以去除碳酸盐组分,用蒸馏水反复洗涤至中性,过滤后于65℃烘干至恒重,采用TOC-同位素质谱仪(CCIA-38,ABB Inc.)测定稳定同位素 δ13C 值。
1.4 数据处理和统计分析
采用独立样本T检验分析不同林型同一土层之间土壤碳、氮、磷和碳稳定同位素的显著性差异。利用单因素方差分析(one-way ANOVA)和最小显著差异法(LSD)检验同一林型不同土层之间土壤碳、氮、磷和碳稳定同位素的显著性差异。运用Pearson相关性系数和线性回归分析检验土壤碳、氮、磷和碳稳定同位素值之间的相关性关系。采用双因素方差分析(two-way ANOVA)检验不同林型和土层及其交互作用对各土层碳、氮、磷以及碳稳定同位素的影响。以上分析在 Excel 2013、SPSS 27.0(IBM Corp.)和 Prism 8.0(Graphpad Software Inc.)中运行,显著性水平设定为=0.05。
2 结果与分析
2.1 不同林型土壤剖面各层次碳、氮、磷含量分布特征
由图1可知,研究区岷江冷杉林和粗枝云杉林土壤有机质层碳含量变化范围分别为94.5~449.8 g/kg和 171.5~414.3 g/kg,矿质土壤层为 12.0~147.9 g/kg和24.2~138.4 g/kg。研究区岷江冷杉林和粗枝云杉林土壤有机质层碳含量变化范围分别为94.5~449.8 g/kg和 171.5~414.3 g/kg,矿质土壤层为 12.0~147.9 g/kg和24.2~138.4 g/kg。岷江冷杉林和粗枝云杉林土壤磷含量在新鲜凋落物层、半分解层和腐殖质层的范围分别为 0.24~0.33、0.26~0.34、0.43~0.76;0.24~0.34、0.25~0.37 和 0.40~0.71 g/kg,在矿质层的范围分别为0.38~0.60 g/kg和 0.40~0.55 g/kg。双因素方差分析结果表明(表1),林型对于土壤碳含量具有显著影响(P<0.05),土层以及林型与土层的交互作用对土壤碳含量的影响极显著(P<0.01)。土壤氮含量受到林型和土层的显著影响(P<0.01),但林型与土层的交互作用对氮含量无显著影响。在同一林型不同土层中,岷江冷杉林和粗枝云杉林的土壤氮含量在有机层显著高于矿质层。土壤磷浓度在林型、土层及两者交互作用这3种因素中,仅在土层中具有极显著差异。岷江冷杉林和粗枝云杉林在腐殖质层土壤磷含量显著高于其余土层(P<0.05),新鲜凋落物层和半分解层差异不显著,但均显著低于其余土层(P<0.05)。在各层土壤之间,两种林型差异不明显,仅有5~10 cm矿质层土壤磷含量表现为岷江冷杉林显著高于粗枝云杉林(P<0.05)。
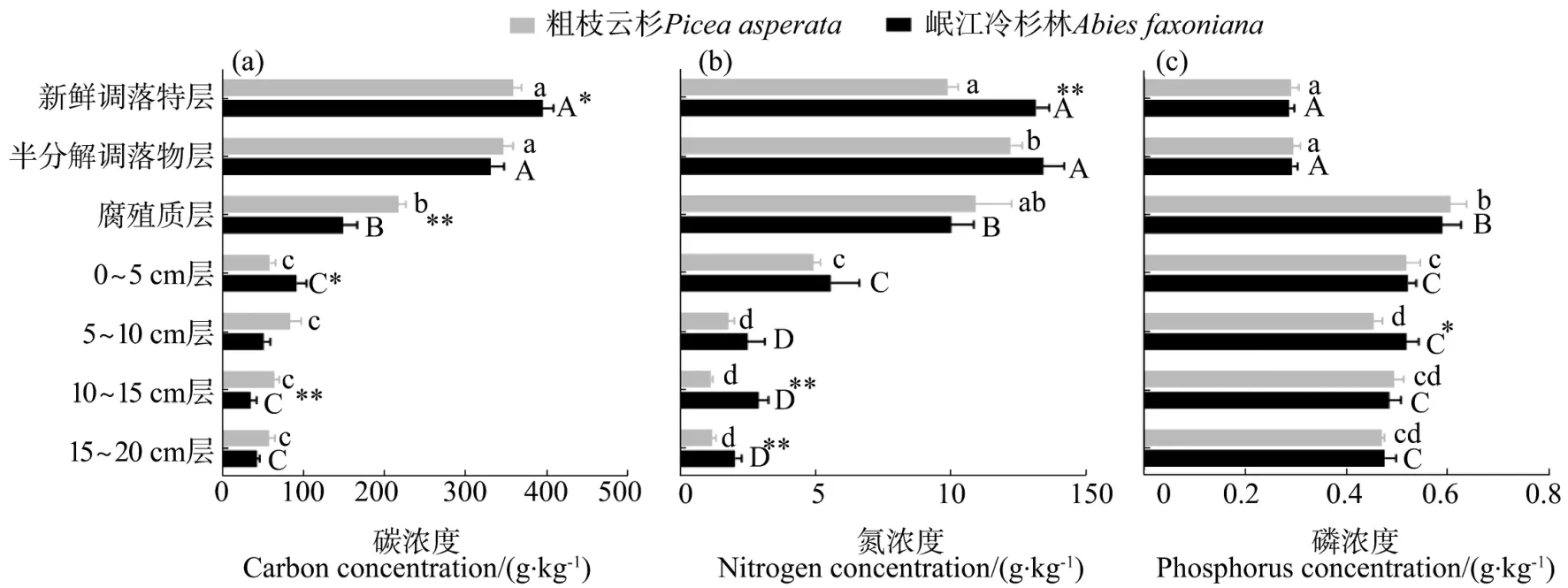
图1 岷江冷杉林和粗枝云杉林土壤剖面各土层碳、氮、磷浓度分布特征Figure 1 Concentrations of carbon,nitrogen and phosphorus in different soil layers of Abies faxoniana and Picea asperata forests
2.2 不同林型土壤剖面各层次碳稳定同位素分布特征
土壤有机碳δ13C值与林型、土层、林型与土层的交互作用3种因素都存在极显著差异(P<0.01)(表1)。岷江冷杉林和粗枝云杉林土壤δ13C值在有机层的含量低于矿质层(图2),在有机层δ13C值表现为腐殖质层<新鲜凋落物层<半分解层。岷江冷杉林的腐殖质层δ13C值与半分解层具有显著差异(P<0.05),但均与新鲜凋落物层的差异不显著;而粗枝云杉林腐殖质层中的δ13C值与新鲜凋落物层、半分解层都具有显著差异(P<0.05)。在矿质层中,岷江冷杉林δ13C值维持在相似水平,粗枝云杉林δ13C在矿质层含量呈现较小波动。在不同林型同一土层中,岷江冷杉林δ13C值均低于粗枝云杉林,且在腐殖质层中差异显著(P<0.05),在 0~5、10~15 和 15~20 cm矿质土壤层中差异极显著(P<0.01)。相关性分析和线性回归分析结果表明有机碳δ13C值与土壤碳和氮含量呈显著负相关,但与磷含量并不具有相关性(P>0.05)(图 3,表 2)。

表1 岷江冷杉林和粗枝云杉林土壤剖面碳、氮、磷浓度和碳稳定同位素(δ13C)的双因素方差分析Table 1 Two way-ANOVA on the effects of forest type and soil layer on C,N,P concentration and δ13C value in the soil profile of the Abies faxoniana and Picea asperata forests

图2 岷江冷杉林和粗枝云杉林土壤剖面各土层碳稳定同位素(δ13C)分布特征Figure 2 δ13C value in the soil profile of the Abies faxoniana and Picea asperata forests

图3 岷江冷杉林和粗枝云杉林土壤剖面碳稳定同位素(δ13C)与碳、氮浓度的线性回归分析Figure 3 Linear regression of δ13C and C,N concentration in the soil profile of the Abies faxoniana and Picea asperata forests

表2 岷江冷杉林和粗枝云杉林土壤剖面碳、氮、磷和碳稳定同位素(δ13C)的Pearson相关性分析Table 2 Pearson correlation coefficient among carbon,nitrogen,phosphorus concentration and δ13C value in the soil profile of the Abies faxoniana and Picea asperata forests
3 讨论
3.1 不同林型对土壤剖面碳氮磷含量和碳稳定同位素特征的影响
土壤有机碳稳定同位素组成δ13C和C/N比值都可用于研究土壤有机质的不同来源[23]。由于土壤有机质长期存留过程中都携带有特定的同位素信号,其周转速率可以决定δ13C值的变化,因此理论上土壤稳定同位素的组成特征会随着不同森林类型的地下物质循环过程的不断发生而产生差异[24]。本研究中,相较于粗枝云杉林,岷江冷杉林各层土壤δ13C值更趋向于正值,且随土壤深度增加这种差异更为显著,说明土壤有机质在岷江冷杉林中分解得更为彻底,且岷江冷杉林的土壤条件相对于粗枝云杉林更加稳定。另外,两种林型土壤δ13C与刁浩宇等在长白山针对阔叶红松林对该元素的研究结果相似[25],证明不同林型的土壤碳稳定同位素组成存在明显差异。
在相同气候区域的不同森林类型中,地上植被吸收养分元素的差异以及凋落物质量和数量以及分解速率的差异,都将影响土壤碳和氮、磷养分以及δ13C在土壤中的分布特征[26]。本研究结果表明,岷江冷杉林与粗枝云杉林相比较,其土壤碳含量更低、土壤氮含量更高,与谌贤等探究该两种林型的研究结果一致[27]。邓仁菊等与贺若阳等的研究结果显示岷江冷杉林土壤碳、氮含量均高于粗枝云杉林[26,28]。本研究中两种林型土壤碳含量分布的差异可能源于粗枝云杉林为人工林,在造林过程中所受到的人为因素干扰更剧烈有关。土壤磷含量在两种林型中无显著差异,这可能与磷的主要来源为母质层的风化,而两种林型母质层差距较小,因此土壤磷含量受林型影响较小。本研究土壤碳氮磷的含量分布也与一些研究结果有差异[26-28]。有研究表明,季节变化会造成土壤碳、氮、磷含量的差异,因此不同研究中含量差异可能源于季节的影响[29]。
3.2 不同土壤层次对土壤剖面碳氮磷含量和碳稳定同位素特征的影响
已有研究表明土壤碳含量随土壤深度的增加而降低[28-29],本研究中土壤碳含量所表现出的垂直分布情况与之相一致。其主要原因是土壤碳的主要来源之一是凋落物的分解,因此表层土壤相较于矿质层土壤,其碳含量会相对较高[30]。土壤δ13C在研究生态系统碳循环过程中具有重要地位,其含量可为研究土壤周转周期提供重要数据基础。已有研究表明土壤δ13C通常随着土壤深度的增加而增加,且不同林型的有机层中,新鲜凋落物层与半分解层、腐殖质层的含量大小并不能肯定[25]。本研究结果显示,岷江冷杉林和粗枝云杉林的土壤δ13C在有机层中均为先增后减,但是在矿质层中,岷江冷杉林土壤δ13C便随着土壤深度的增加而减少,这说明在岷江冷杉林中土壤δ13C快循环组分损失程度是随着土壤深度增加而加快的,土壤δ13C慢循环组分也经历着在不断分解的过程[25];粗枝云杉林则没有明显规律,这可能是因为粗枝云杉林为人工林,在栽培过程中土层自然状况受到了强烈的人为干扰所导致。
土壤氮作为植物从土壤中吸收量最大的矿质元素,在森林生态系统中主要来源于枯枝落叶、生物固氮等,其含量影响着氮元素形态的转化,进而影响森林生态系统的氮素循环过程[4,31-32]。在本研究中,土壤氮含量与土壤碳含量变化趋势相同,与碳含量具有极显著正相关关系。这与土壤碳和土壤氮均主要来源于凋落物的分解有关,且本研究区域温度较低,凋落物分解速度相对较慢,因此碳和氮在表层土壤的积累量较高。土壤磷在不同土壤层次的含量变化趋势表现较为复杂。有研究结果表明土地利用方式会影响土壤磷的含量,一为土壤磷含量随着土壤深度的加深而减少,二为在有机层中土壤磷含量同样随着土壤深度的增加而增加,但在矿质层保持基本一致[33];另有研究结果显示土壤磷含量在不同土壤深度中差异不大,但仍有土壤深度越深,土壤磷含量越低的趋势。此外,邓仁菊等在对川西亚高山森林的研究中测得:土壤磷在从半分解层到腐殖质层再到母质层过程中,其含量为先增加再降低[28]。本研究中土壤磷含量基本表现为腐殖质层高于其余层次,矿质层之间相差不大且略低于腐殖质层,新鲜凋落物层和半分解层远低于腐殖质层和矿质层,整体趋势与邓仁菊等的研究结果相似[28],并且土壤磷在矿质土壤层的分布趋势与吕金林等的研究结果一致[34]。磷在土壤垂直剖面上的变化趋势与碳、氮不同,其主要原因是元素的来源不同,磷受土壤母质风化的影响较大[35],本研究区域与邓仁菊等研究区域同处川西亚高山森林,因此其含量相似度更高[28]。
4 结论
研究表明,不同森林类型和土壤层次显著影响了土壤碳氮含量和碳稳定同位素组成的分布特征,但磷含量仅在不同土层的分布中有显著差异。总体而言,岷江冷杉林各土层碳含量低于粗枝云杉林,而氮含量相反;土壤碳、氮含量在岷江冷杉林和粗枝云杉林中均为随着土层深度的增加而减少;磷含量在腐殖质层和矿质层显著高于新鲜凋落物层和半分解层;土壤有机层δ13C值高于矿质层,且岷江冷杉林土壤δ13C总体低于粗枝云杉林。在两种林型中,土壤碳含量与氮含量呈极显著正相关,与磷含量和碳稳定同位素组成均呈显著负相关,而土壤磷含量与土碳稳定同位素组成无相关关系。研究结果可为深入了解川西亚高山针叶林生态系统的物质循环过程提供一定的科学依据。
