湖北省随州地区稻瘟病菌群体结构分析及生态模式菌群的建立
2021-06-29吴尧韩玉揭春玉王林吴双清
吴尧 韩玉 揭春玉 王林 吴双清


摘要 :采用人工接種方法,用7个中国鉴别品种和95个自选资源材料对2014年至2015年间采自湖北省随州地区的76个稻瘟病菌单孢菌株进行致病型鉴定。分别利用传统生理小种种群划分方法和近邻传播聚类法(affinity propagation clustering,AP聚类法)进行菌群结构分析,比较两种方法的差异。结果表明,传统生理小种种群划分方法将76个菌株分为6个菌群,14个中国小种, ZB菌群为优势种群,占比65.79%,ZB15为优势生理小种,占比达到60.53%;近邻传播聚类法将76个菌株分为14个菌群,Cluster 2、Cluster 6、Cluster 7、Cluster 8为随州地区优势菌群,占比分别为18.42%、19.74%、22.37%、17.11%,Cluster 6、Cluster 7和Cluster 8具有相对较高的致病力;两种方法划分结果虽不完全一一对应,但划分趋势相同,近邻传播聚类法在稻瘟病菌种群结构研究中具有一定应用前景;根据试验结果筛选出1个代表随州地区稻瘟病总致病谱的菌群,菌群由12个菌株组成,将其命名为“随州生态模式菌群”。
关键词 :湖北; 随州; 稻瘟病菌; 菌群结构; 生态模式菌株; 近邻传播聚类
中图分类号:
S 435.111.41
文献标识码: A
DOI: 10.16688/j.zwbh.2020122
Population structure analysis of rice blast fungus (Magnaporthe oryzae) and
the establishment of ecological type flora in Suizhou area of Hubei province
WU Yao, HAN Yu, JIE Chunyu, WANG Lin, WU Shuangqing*
(Enshi Tujia and Miao Autonomous Prefecture Academy of Agricultural Sciences, Hubei Province, Enshi 445000, China)
Abstract
Seven Chinese cultivars and 95 self-selected varieties were used to artificially inoculate and identify 76 isolates of Magnaporthe oryzae collected from Suizhou area of Hubei province from 2014 to 2015. The population structure was analyzed by using traditional race population division method and affinity propagation clustering. The differences between the two methods were compared. The results showed that 76 isolates were divided into six groups and 14 Chinese races by the traditional race population division method. ZB group was the dominant population, accounting for 65.79%, and ZB15 was the dominant race, accounting for 60.53%. 76 isolates were divided into 14 clusters by affinity propagation clustering, and Cluster 2, Cluster 6, Cluster 7 and Cluster 8 were the dominant groups in Suizhou, accounting for 18.42%, 19.74%, 22.37%, and 17.11%, respectively. Cluster 6, Cluster 7 and Cluster 8 have relatively high pathogenicity. Although the division results of the two methods are not completely one-to-one, but the division trend is the same. The affinity propagation clustering has certain application prospects in studying the population structure of M.oryzae. A group of isolates, representing the total pathogenic spectrum of M.oryzae in Suizhou area, was selected, consisting of 12 isolates, which was named “Suizhou ecological type flora”.
Key words
Hubei; Suizhou; Magnaporthe oryzae; population structure; ecological type isolate; affinity propagation clustering
稻瘟病是影响水稻生产的主要病害之一,水稻整个生育期均可感染稻瘟病。稻瘟病大暴发在我国各水稻种植区时有发生。2014年和2015年湖北省中稻稻瘟病大发生,除鄂西南山区稻瘟病常发区外,随州、孝感、荆州、荆门等地区稻瘟病也严重发生,其中随州损失最大,造成400 hm2水稻基本绝收[12]。
到目前为止,抗病品种的选育与利用依然是防治稻瘟病最为环保且经济的手段。但在生产中发现,原本抗性表现较好的水稻品种在连续种植多年后,抗性逐渐丧失,其最主要的原因是稻瘟病菌种群结构发生了变化。因此,弄清楚当地稻瘟病菌种群结构,对指导抗病育种和建立稻瘟病持久抗性具有非常重要的意义。
基于“基因对基因”假说理论[3],稻瘟病暴发流行可解释为稻瘟病菌的无毒基因与水稻品种抗病基因间的直接或间接相互作用。因此,利用合适的鉴别品种对稻区菌群进行分析是了解当地稻瘟病菌种群结构及种群多样性的有效手段。为此,国内外学者先后设置了多套鉴别品种。1960年美国学者Latterell用‘辛尼斯等10个品种建立了美国稻瘟病菌生理小种鉴别品种[4],1965年将品种增加至12个[5];1961年日本建立了第一套日本鉴别品种,1976年建立了由9个粳稻品种组成的第二套鉴别品种[67];1961年我国台湾用‘昆山五香粳等16个品种对不同来源菌株进行了鉴定[8];1962年韩国学者用‘辛尼斯等10个鉴别品种对来自不同区域的菌株进行了鉴定[9];1963年菲律宾通过对11个美国鉴别品种、12个日本鉴别品种、16个我国台湾鉴别品种以及东南亚和南亚等地的品种进行鉴别能力筛选,筛选出了由‘克拉罗西等12个品种组成的菲律宾鉴别品种[10];1963年-1965年由日、美学者协作筛选出了‘拉米纳德Str.3等8个品种组成的国际鉴别品种[11];1976年我国建立了全国稻瘟病生理小种联合试验组,利用全国21个省、市、区的1 739个单孢分离物对212个水稻品种进行筛选测定,从中选出‘特特勃‘珍龙13‘四丰43‘东农363‘关东51‘合江18和‘丽江新团黑谷7个品种作为中国鉴别品种[12];20世纪80年代中期,国际水稻研究所育成4个近等基因系[13];Ling等以普感品种‘丽江新团黑谷为轮回亲本,选育出6个单基因近等基因系[14]; 国际水稻所与日本学者合作,分别于2000年将24个稻瘟病抗性基因导入到‘丽江新团黑谷背景中育成31个单基因系,于2010年将11个抗性基因通过连续回交导入‘丽江新团黑谷中育成了20个近等基因系,于2011年将14个抗性基因通過连续回交导入到‘CO39中育成了27个CO39-ML[1517];2015年印度学者以印度香米‘Pusa Basmati 1为背景通过回交导入7个抗性基因,育成了36个近等基因系,包含14个单基因系,16个双重基因堆叠系(two-gene pyramids),6个三重基因堆叠系(three-gene pyramids)[18]。
为理清随州地区稻瘟病菌种群结构,2018年至2019年间笔者使用95个自选资源材料和7个中国鉴别品种对湖北随州稻瘟病大发生期间采集的菌株进行致病型鉴定。利用传统生理小种种群划分方法,并探索性地使用近邻传播聚类法对菌群结构进行分析。本文是对随州地区稻瘟病菌种群结构调查的首次报道,试验结果对随州地区以及江汉平原抗稻瘟病育种和水稻品种布局具有重要参考意义。
1 材料与方法
1.1 参试菌株
参试稻瘟病菌单孢菌株76个(表1),从随州市何店镇谌家岭村、何店镇王店村、厉山镇、万店镇夹子沟村、新街镇刘家岗村等5处地点采集的43份标样中分离得到,标样采集时间为2014年至2015年,单孢分离时间为2018年。
1.2 鉴别材料
鉴别材料共102个,包括7个中国鉴别品种和95个自选资源材料。7个中国鉴别品种分别为:‘特特勃‘珍龙13‘四丰43‘东农363‘关东51‘合江18和‘丽江新团黑谷。95个自选资源材料为恩施自治州农业科学院植保土肥所经30余年收集改良筛选出的稻瘟病监测材料。
1.3 育苗与接种
试验采用苗期孢子悬浮液人工喷雾接种,接种、隔离、诱导发病均在恩施自治州农业科学院作物病害鉴定大棚内进行,具体操作如下:
在1.5 m×1.5 m×0.25 m的育秧池内育苗,池内填充肥沃菜园土,灌水整平。每个育秧池内条播3行‘丽江新团黑谷分厢,同时作为诱发品种和指示品种。厢内每个鉴别材料播种1行,每行5粒种子,株距2.5 cm,行距5 cm。秧苗生长期间酌情增施氮肥,保持叶片嫩绿。秧龄达到三叶一心至四叶期时择阴雨天傍晚进行接种。
菌株诱导产孢[19],用0.02% 吐温-20溶液洗脱孢子,孢子悬浮液浓度为100倍显微镜视野下10个以上。接种至调查结束,整个时间段均用自制白色无纺布罩将育秧池罩住,保持各育秧池之间严格隔离。用自制文丘里喷头连接空气压缩机喷雾,一个菌株接种一个育秧池,每个育秧池内接种300 mL菌液,接种完毕后遮光处理48 h,后期适时喷雾降温保湿,维持大棚温湿度相对恒定。
1.4 调查与分级
接种10 d后开始调查,分级标准参考《水稻品种试验稻瘟病抗性鉴定与评价技术规程(NY/T 2646-2014)》[20],发病等级由轻到重分为0~9共10个等级。调查2次,间隔7 d。最终统计数据取两次调查值的最高值。0级至3级定义为抗病,记为“R”,4级至9级定义为感病,记为“S”。
1.5 种菌结构及多样性分析
根据鉴别品种的反应类型,“S”记为1,“R”记为0;中国小种的划分方法参照全国稻瘟病菌生理小种联合试验组的标准进行[12]。
近邻传播聚类算法是将全部样本看作网络的节点,然后通过网络中各条边的消息传递计算出各样本的聚类中心。聚类过程中,共有两种消息在各节点间传递,分别是吸引度(responsibility)和归属度(availability)。近邻传播聚类算法通过迭代过程不断更新每一个点的吸引度和归属度值,直到产生m个高质量的exemplar(类似于质心),同时将其余的数据点分配到相应的聚类中[21]。统计分析及作图软件使用R-3.5.1。
1.6 生态模式菌群的构建
稻瘟病生态模式菌群是指在特定生态稻区内通过稻瘟病单孢致病型监测及其相关数据分析,选择代表该生态稻区稻瘟病菌种群综合致病力的不同致病型菌株组成的群体。
筛选原则是:首先找出致病谱最广的菌株A,观察菌株A不能侵染的品种,将其作为监测品种,从剩余菌株中找出能侵染这些监测品种个数最多的菌株B,若有多个菌株侵染个数相同,则比较它们的总致病率,总致病率最高的菌株入选;然后再把菌株A和B都不能侵染的品种作为监测品种,从剩余菌株中找出能侵染这些监测品种个数最多的菌株C,依次类推,直到组合菌株不能侵染的监测品种也不能被剩余的所有菌株侵染为止。
2 结果与分析
2.1 中国小种划分
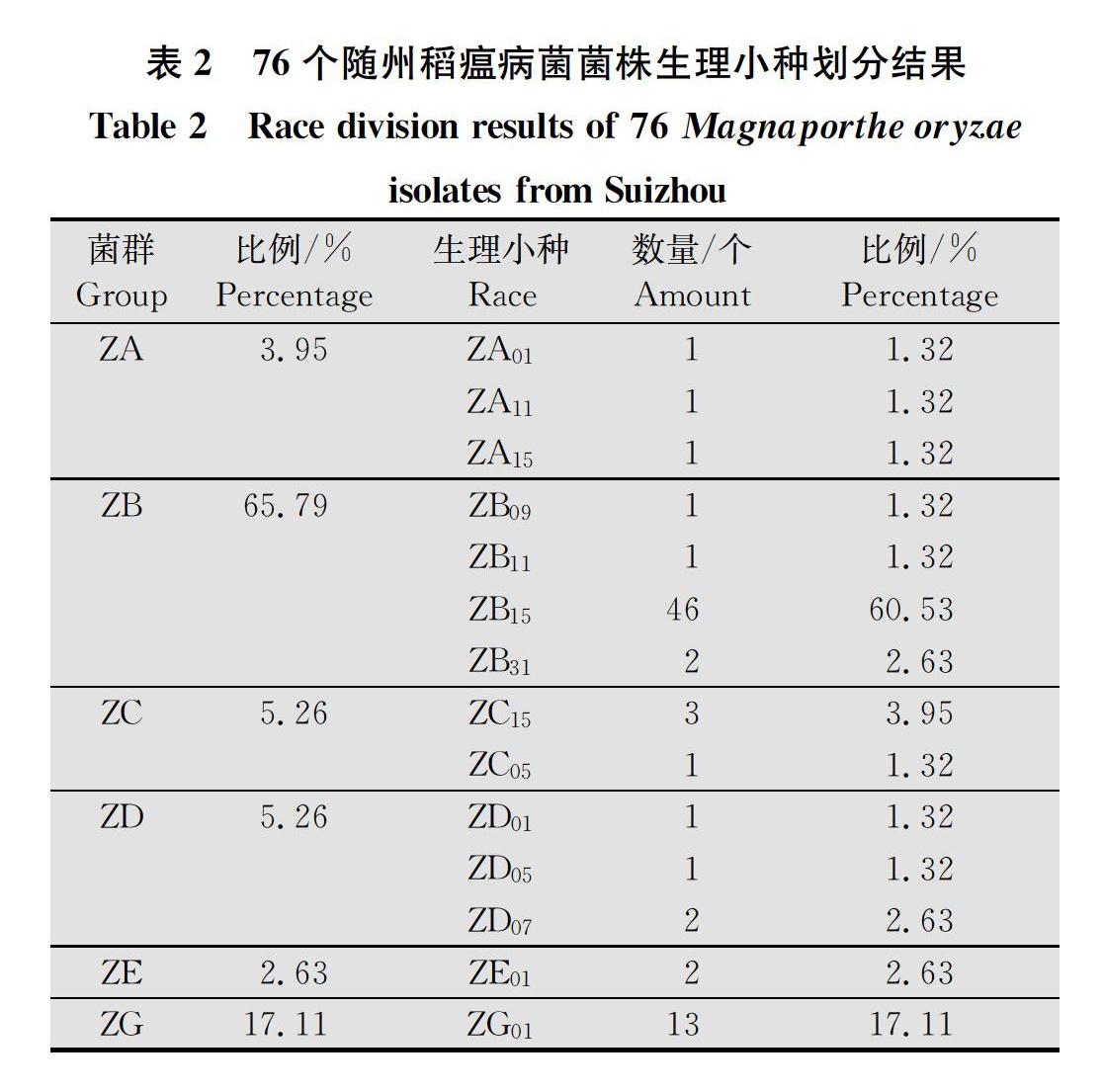
76个单孢菌株在7个中国鉴别品种上进行鉴定,按鉴别品种发病表现可将菌株分为6个菌群,14个中国小种(表2)。ZA群出现频率为3.95%,包括ZA01、ZA11、ZA15等3个小种,出现频率均为1.32%;ZB群出现频率为65.79%,包括ZB09、ZB11、ZB15、ZB31等4个小种,出现频率分别为1.32%、1.32%、6053%、2.63%;ZC群出现频率为5.26%,包括ZC05和ZC15两个小种,出现频率分别为1.32%和3.95%;ZD群出现频率为5.26%,有ZD01、ZD05、ZD07等3个小种,出现频率分别为1.32%、1.32%、2.63%;ZE群出现频率为2.63%,有ZE011个小种;ZG群出现频率为17.11%,有ZG011个小种。
其中ZB菌群为优势种群,占比65.79%;其次为ZG菌群,占比17.11%;出现频率较低的是ZA、ZC、ZD、ZE菌群,合占17.11%;ZF种群未出现;ZB15生理小种占绝对优势,所占比例达到60.53%,ZG01生理小种其次,所占比例为17.11%。
2.2 近邻传播聚类法划分菌群
76个菌株在95个自选资源材料上的反应类型经过近邻传播聚类法分析,结果表明,最佳聚类组数为14个,每个聚类组均给出一个具有代表性样例的菌株。Cluster 1、Cluster 3、Cluster 4、Cluster 9、Cluster 10、Cluster 11、Cluster 12、Cluster 13、Cluster 14等9个聚类组均只含有單个菌株,所占比例均为1.32%,致病率分别为1.05%、13.68%、15.79%、35.79%、35.79%、35.79%、38.95%、38.95%、40.00%。Cluster 2、Cluster 5、Cluster 6、Cluster 7、Cluster 8为多菌株聚类组,所含菌株数量分别为14、8、15、17、13个,所占比例分别为18.42%、10.53%、19.74%、22.37%、17.11%,致病率范围分别为7.37%~12.63%、14.74%~22.11%、15.79%~25.26%、23.16%~32.63%、30.53%~35.79%(表3)。从图1中可以看出,EBSN205-2、EBSN175-2、EBSW157-2、EBSN195-4、EBSN181-1、EBSN170-2、EBSL154-2、EBSN205-4、EBSN184-1与其他菌株的交叉点颜色均较深,差异较大。在聚类树上这9个菌株处在相似水平最低的位置,而它们之中又以EBSN205-2具有最低的相似水平。这9个菌株各自单独形成聚类组,分别对应Cluster 12、Cluster 14、Cluster 13、Cluster 11、Cluster 1、Cluster 9、 Cluster 10、 Cluster 3、Cluster 4。其他多菌株聚类组内菌株之间矩阵交叉点颜色较浅,相似度较高。此外,可以看出Cluster 6、Cluster 7、Cluster 8虽然被分为3个聚类组,但它们处在同一分支下,菌株之间矩阵交叉区域颜色较浅,相似度较高。Cluster 2和Cluster 5虽然也处于同一分支,但该分支在聚类树上处于相似水平较低的位置,聚类组间菌株矩阵交叉点颜色也相应较深,相似度较低。
由此可见,Cluster 2、 Cluster 6、 Cluster 7、 Cluster 8在随州地区为优势菌群,Cluster 6、Cluster 7和Cluster 8具有较高致病力,且相似水平较高。EBSN175-2为最强致病力菌株,所在的Cluster 14菌株数量少,暂未形成规模暴发。
2.3 两种菌群划分方法比较
76个菌株在7个中国鉴别品种上被分为6个菌群,而在95个自选常规水稻品种上被分为14个菌群。虽然两种方法划分的菌群没有出现严格的一一对应的关系,但通过观察可以发现,两种方法的划分趋势是一致的,如ZG01对应Cluster 2,ZB15主要对应Cluster 6、Cluster 7和Cluster 8,仅个别聚类组与生理小种划分对应关系不明显,如Cluster 5聚类组中的菌株无规律地分散于ZB、ZC、ZD、ZE、ZG等多个生理小种菌群中(表4)。结果表明,利用95个自选资源材料结合近邻传播聚类法对稻瘟病菌株进行聚类分析划分菌群较传统生理小种划分具有更高的分辨率,且划分趋势相同,用于致病型分析是可行的。
2.4随州生态模式菌群
分析76个随州稻瘟病菌单孢分离物在102个鉴别材料上的反应,最终从中筛选出了1个“随州生态模式菌群”,该模式菌群致病谱与76个随州稻瘟病菌单孢分离物的总致病谱相同,总致病率均为76.47%。模式菌群包含有12个菌株,致病率从15.69%至44.12%,分属于ZA、ZB、ZC、ZE等4个生理小种群或Cluster 3、4、5、6、7、8、9、11、12、13、14等11个聚类群(表5)。
3 结论与讨论
水稻品種的抗感表现是由稻瘟病菌的致病力、环境适应性、寄主的抗病力及敏感生育期等多种遗传特性以及特定的环境因素共同决定的,因此利用稻瘟病菌和水稻品种之间的相互关系来分析稻瘟病菌致病型和评价抗病品种抗病力是符合实际生产规律的。
本研究利用7个中国鉴别品种和95个自选资源材料,对2014年至2015年采自随州的76个稻瘟病菌单孢菌株进行了致病型分析。76个稻瘟病菌菌株被分为6个菌群,14个中国小种,ZB为优势种群,ZB15和ZG01为优势生理小种,且出现频率远高于其他小种。20世纪80年代末和90年代初有报道称,湖北省生理小种优势种群主要是ZB和ZG[2224],此后较长时间研究报道湖北省生理小种优势种群转变为ZA[2528],但2009年后相关报道认为,ZB出现频率增加,又逐渐转变为次优势种群和优势种群[19, 2930]。本研究也进一步指向了这一变化趋势,由此推断近年来随州地区乃至江汉平原稻瘟病大暴发的现象极有可能是由于南方籼型菌群ZB大量增殖所造成。
本研究探索性地在植物病原致病型分类领域使用了近邻传播聚类分析法,76个稻瘟病菌被自动划分为14个菌群。近邻传播聚类分析法和传统生理小种种群划分法得到的划分趋势相对一致,且近邻传播聚类分析法具有相对较高的分辨率。传统生理小种种群划分方法的鉴别品种固定为7个,根据相应划分方法,不同来源菌株最多可划分为ZA至ZH共8个菌群,鉴别能力有一定局限性。相较通常使用的非加权组平均法(UPGMA),近邻传播聚类分析法无需通过指定特定相似水平来确定聚类数,对使用者的经验要求较低。由此可见利用近邻传播聚类分析法结合自选资源材料分析稻区稻瘟病菌种群是可行的。但值得注意的是本研究使用的自选资源材料所含抗性基因不明确,因此暂无法弄清楚湖北随州稻瘟病菌无毒基因型分布结构,今后有必要通过分子生物学手段探明这95个自选资源材料遗传背景,进一步删减补充优化,以期建立一套适合湖北及周边省市的稻瘟病鉴别材料。
通过分析76个随州稻瘟病单孢菌株在102个鉴别材料上的反应,本研究筛选出了1个由12个稻瘟病菌株组成的“随州生态模式菌群”。该模式菌群致病率与76个参试菌株总致病率完全一致,在一定程度上能够代表随州地区的稻瘟病菌致病力。另外值得注意的是,该菌群致病率仅为76.47%,部分抗性较强的鉴别品种未发现感病,因此推测可能试验所采用的菌株样本在时空上对调查区域覆盖相对较窄,群体较小。在今后的试验中,我们将扩大调查群体,在更大区域内持续动态监测随州地区稻瘟病菌种群结构,对这一观点进行验证,对该菌群加以更新完善,提高其总致病率。今后若将这套菌株运用于检测随州及江汉平原水稻主栽品种抗稻瘟病能力,将对该区域水稻品种布局和抗病品种选育具有极大的参考价值。
参考文献
[1] 刘芹, 张求东, 陈雁, 等.湖北省2014年中稻稻瘟病局部流行的原因分析[J].湖北植保, 2015(2): 4345.
[2] 刘芹, 袁浩, 杨俊杰, 等.湖北省2015年水稻稻瘟病发生特点及原因分析[J].湖北植保, 2017(5): 4647.
[3] FLOR H H. Current status of the gene-for-gene concept [J]. Annual Review of Phytopathology, 1971, 9: 275296.
[4] LATTERELL F M, TULLIS E C, COLLIER J W. Physiologic races of Piricularia oryzae Cav. [J]. Plant Disease Reporter, 1960, 44: 676683.
[5] LATTERELL F M, MARCHETTI M A, GROVE B R. Co-ordination of effort to establish an international system for race identification in Pyricularia oryzae. The rice blast disease [M]. Baltimore Maryland. US: Johns Hopkins Press, 1965: 257274.
[6] 山崎义人, 高坂淖尔. 稻瘟病与抗病育种[M]. 凌忠专, 孙昌其, 译. 北京: 农业出版社, 1990: 353.
[7] YAMADA M, KIYOSAWA S, YAMAGUCHI T, et al. Proposal of a new method for differentiating races of Pyricularia oryzae Cavara in Japan [J]. Annals of the Phytopathological Society of Japan, 1976, 42: 216219.
[8] WOO S S. Some experimental studies on the inheritance of resistance and susceptibility to rice leaf blast disease, Piricularia oryzae Cav.[J]. Botanical Bulletin Academia Sinica, 1965, 6: 208217.
[9] LEE S C, MATSUMOTO S. Studies on the physiologic races of rice blast fungus in Korea during the period of 19621963 [J]. Annals of the Phytopathological Society of Japan, 1966, 32: 4045.
[10]BANDONG J M, OU S H. The physiologic races of Piricularia oryzae Cav. in the Philippines [J]. Philippine Agriculture, 1966, 49: 655667.
[11]ATKINS J G, ROBERT A L, ADAIR C R, et al. An international set of rice varieties for differentiating races of Piricularia oryzae [J]. Phytopathology, 1967, 57: 297301.
[12]全国稻瘟病生理小种联合试验组.我国稻瘟病菌生理小种研究[J].植物病理学报, 1980(2): 7182.
[13]MACKILL D J, BONMAN J M. Inheritance of blast resistance in near-isogenic lines of rice [J]. Phytopathology, 1992, 82: 746749.
[14]LING Zhongzhuan, MEW T V, WANG Jiulin, et al. Development of near-isogenic lines as international differentials of the blast pathogen [J]. International Rice Research Newsletter, 1995, 20(1): 1314.
[15]HIROSHI T, MARY J T Y, EBRON L A, et al. Development of monogenic lines of rice for blast resistance [J]. Breeding Science, 2000, 50: 229234.
[16]TELEBANCO-YANORIA M J, KOIDE Y, FUKUTA Y, et al. Development of near-isogenic lines of Japonica-type rice variety Lijiangxintuanheigu as differentials for blast resistance [J]. Breeding Science, 2010, 60: 629638.
[17]TELEBANCO-YANORIA M J, KOIDE Y, FUKUTA Y, et al. A set of near-isogenic lines of Indica-type rice variety CO39 as differential varieties for blast resistance [J]. Molecular Breeding, 2011, 27: 357373.
[18]KHANNA A, SHARMA V, ELLUR R K, et al. Development and evaluation of near-isogenic lines for major blast resistance gene(s) in Basmati rice [J]. Theoretical and Applied Genetics, 2015, 128: 12431259.
[19]吴尧, 吴双清, 王林, 等. 湖北稻瘟病病菌种群鉴定与分析[J].湖北农业科学, 2014, 53(21): 51595162.
[20]农业部种子管理局.水稻品种试验稻瘟病抗性鉴定与评价技术规程: NY/T 26462014 [S].北京:中国农业出版社, 2014.
[21]FREY B J, DUECK D. Clustering by passing messages between data points [J]. Science, 2007, 315(5814): 972976.
[22]艾仁孝, 颜学明. 恩施地区稻瘟病菌生理小种研究[J].湖北农业科学, 1981(12): 2025.
[23]侯明生, 杨绳桃, 王清锋, 等. 湖北稻瘟病菌生理小種种群及其分布的研究[J].湖北农业科学, 1988(5): 1720.
[24]陈永坚, 赵永静, 肖炎农. 湖北省稻瘟病菌生理小种鉴定[J].湖北农业科学, 1992(6): 2224.
[25]杨小林, 喻大昭, 陈其志, 等. 湖北省稻瘟病生理小种的组成与分布[J].植物病理学报, 2005(S1): 204.
[26]杨小林, 陈其志, 张舒, 等. 湖北省稻瘟病菌生理小种的组成与分布[J].华中农业大学学报, 2006(2): 132133.
[27]沈彬. 湖北稻瘟病菌生理小种和水稻抗瘟性的鉴定及生化机制研究[D]. 武汉:华中农业大学, 2007.
[28]杨小林, 陈其志, 张舒, 等. 稻瘟病菌在中国鉴别品种及CO39近等基因系上的致病型分析[J].湖北农业科学, 2009, 48(12): 30123014.
[29]王永崇. 湖北省稻瘟病菌群体遗传多样性及水稻品种(系)抗瘟性研究[D]. 武汉:华中农业大学, 2009.
[30]杨小林, 张舒, 吕亮, 等. 基于中国鉴别品种及单基因品系的湖北省稻瘟病菌的致病型分布[J].湖北农业科学, 2010, 49(11): 27792781.
(责任编辑:田 喆)
