红蓝光交替照射下生菜能量利用与光合性能分析
2021-06-29陈晓丽杨其长王利春李友丽郭文忠
陈晓丽 杨其长 王利春 李友丽 郭文忠
(1.北京农业智能装备技术研究中心, 北京 100097; 2.中国农业科学院都市农业研究所, 成都 610213)
0 引言
光是植物生长发育的能量来源及代谢调节信号,通过光质、光强以及光周期三方面作用于植物[1-3]。其中,红光和蓝光光质因其对应于植物光合色素最大吸收波段而成为近年来的研究重点[4-8]。研究表明,红蓝光同时照射植物的效果往往优于红光或蓝光单色光照射效果[3,9-11]。然而,一些研究认为,红光和蓝光在植物某些生理活动中的作用可能相反、也可能一致[12-14],植物对红光和蓝光的需求机制以及红、蓝光的作用途径之间的关系尚不明确。随着LED及光配方系统调控硬件的深度开发,光配方不再局限于对光质、光强度以及光期的调节,而是可以拓展到供光模式的调节,如高频率的交替供光、间歇供光等,这些为深入研究红、蓝光作用途径之间的关系提供了硬件保障。
在总光量一致的基础上,渐变式光强照射下甜土豆的干质量是连续光照射下的1.1倍,渐变式供光模式下光源的能量利用率得到了提高[15]。研究显示,与恒定的连续光照射相比,强、弱交替供光提高了辣椒叶黄酮醇的积累,同时刺激了花青素和类胡萝卜素的合成,而且这种交替照射似乎也有利于降低单一蓝光对植物的负面作用[16]。有研究指出,在日累积光积分相等的前提下,与红蓝同时供光相比,红、蓝光交替供光显著促进了生菜植株的生长[17]。还有研究表明,在相同能耗基础上,与红蓝光同时供光相比,以8 h和1 h为间隔进行红光和蓝光交替照射的处理,其生菜的生物量有所提高[18]。以上研究表明,与常规连续供光模式相比,非连续供给的红蓝光能够通过优化产量或某些指标而达到提升光源电能利用率的目的。除了常见的光因素(光质、光强、光周期)之外,供光模式(渐变供光、交替供光、间歇供光)也对植物生长和生理过程产生较大的影响,并且有提高人工光源电能利用率的可能,因此,综合考虑植物产出效益和植物工厂系统电能利用效率,研究相同能耗条件下交替供光策略的潜在优势具有重要的实际应用价值。
本文设置不同交替间隔时长的红蓝交替光处理,以红蓝光同时供光为对照,探究不同交替模式对生菜能量利用率及叶片光合特性、荧光特性的影响,以期揭示红、蓝光在作用于植物生长发育过程中的相互关系,为植物工厂高能量利用率的供光方式提供理论依据。
1 材料与方法
试验在北京农业智能装备技术研究中心的全人工光型植物工厂(长×宽×高:5 m×4 m×2.8 m)中进行,试验材料为奶油生菜(LactucasativaL.)。将生菜种子播种至海绵块中育苗,14 d后定植到不同光环境的水培种植箱中,种植密度为25株/m2。植物工厂内昼/夜温度设置为24℃/20℃,空气相对湿度65%,CO2摩尔比700 μmol/mol,霍格兰营养液[19]pH值、EC分别保持在6.5和1.45 mS/cm左右,营养液7 d更换一次。从播种日起第54天(即定植后40 d)进行收获并测定收获指标。
试验共8个处理,试验光源采用北京农业智能装备技术研究中心研制的LED植物光配方调控系统,光源垂直悬挂于植物顶部。该系统可以设置不同光质、光量配比以及供光模式和照射频率。定植当天即开始不同的光处理照射。光处理如表1所示,共设5个红蓝交替光处理、1个红蓝光同时供光处理以及纯红、纯蓝光处理。纯红光与纯蓝光的处理中,红光和蓝光光强度均为200 μmol/(m2·s);其他处理中红、蓝光光强度分别设定为180、20 μmol/(m2·s),即整个生育期内红光与蓝光的光量子数比为9。

表1 红蓝交替光试验处理
关于红蓝光交替处理模式的设置和命名方法见表1。在每天16 h的光周期里,红、蓝光每5 min切换1次,交替频率为96次/d,处理记作R/B(5 m),同理红、蓝光交替间隔时间为10、15、30、60 min分别记作R/B(10 m)、R/B(15 m)、R/B(30 m)和R/B(60 m),16 h光期里对应的红蓝光交替频率则分别为48、32、16、8次。纯红光和纯蓝光处理分别记作R、B,红蓝光同时供光的处理为对照,记为RB。关于处理间的光量子数和耗电量见表2。本试验中红蓝光同时供光的处理RB每天的光期为8 h,这是为了该处理作为对照与其他交替红蓝光处理具有相等的总光量和总耗电量,以进行后期各种指标的对比。
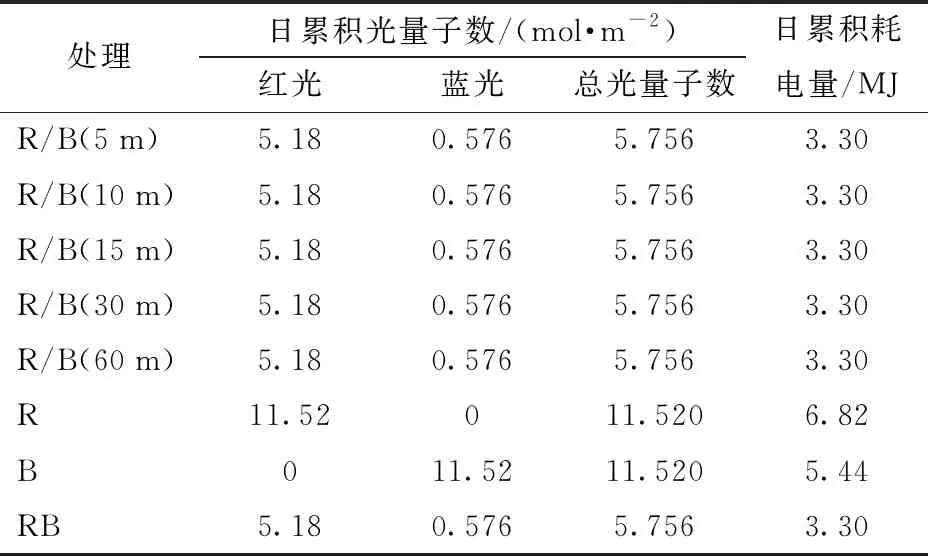
表2 不同处理下的日累积光量子数及耗电量
2 测定与分析
2.1 光合色素测定
从播种日起第54天取样测定光合色素,每处理随机选取6株生菜植株作为待测样品。叶绿素和类胡萝卜素含量测定采用分光光度法[20];数据处理采用 Microsoft Excel 2013,显著性差异分析采用SAS统计分析软件。
2.2 光合测定
采用便携式光合仪(CIRAS-3型,PPSYSTEMS,美国),从播种日起第54天从各处理随机选取生菜植株,统一选定第3片完全展开叶供光合测定。
2.3 荧光测定
从播种日起第54天从各处理随机选取生菜植株,统一选定第3片完全展开叶供荧光测定。采用连续激发式荧光仪(Handy-PEA型,Hansatech, 英国) 测量快速叶绿素荧光诱导动力学曲线 (O-J-I-P荧光诱导曲线)。叶片先暗适应20 min,然后暴露在饱和脉冲光 (3 000 μmol/(m2·s))下1~2 s,分析获得的O-J-I-P荧光诱导曲线,读取并计算PSⅡ反应中心光化学效率及能流分配参数等[21-23],参数分别为:Fo(20 μs时荧光, O相)、Fk(300 μs时荧光, K相)、Fj(2 ms时荧光, J相)、Fm(最大荧光, P相)、RC(反应中心)、ABS(天线色素吸收的光能)、Vj(J点的相对可变荧光)、Mo(O-J-I-P荧光诱导曲线的初始斜率)、CS(单位面积)。相关计算公式为:
暗适应下的PSⅡ最大光化学效率
φ=(Fm-Fo)/Fm
(1)
Ψo=1-Vj
(2)
单位面积吸收(ABS/CS)、捕获(TRo/CS)、用于电子传递(ETo/CS)和热耗散(DIo/CS)的光能
ABS/CS≈Fo
(3)
TRo/CS=φ(ABS/CS)
(4)
ETo/CS=Ψo(TRo/CS)
(5)
DIo/CS=ABS/CS-TRo/CS
(6)
单位面积有活性的反应中心的密度
RC/CS=φ(Vj/Mo)(ABS/CS)
(7)
以吸收光能为基础的性能指数
PⅠabs=(RC/ABS)[φ/(1-φ)][Ψo/(1-Ψo)]
(8)
2.4 能量利用率计算
电能利用率(EUE)的计算公式为
(9)
式中EUEi——电能利用率
DWi、DWi-1——第i、i-1次取样时,生菜植株地上部分平均干质量,g/株(本试验将定植时幼苗干质量忽略不计,即DW0≈0)
Wche——每克干质量对应的化学能,取2×104J/g[24]
S——栽培面积,m2
Di——第i次取样时的栽培密度,株/m2
P——光源的实时工作功率,W
t——第i次和第i-1次取样之间的时间间隔,s
光能利用率(LUE)的计算公式为
(10)
式中LUEi——光能利用率
Wr——单位面积植株冠层接受到的光合有效辐射能,W/m2
单位面积里生产单位质量的生菜所需要的光量子数p(μmol/g)计算公式为
(11)
式中PPFD——光强度,μmol/(m2·s)
T——整个生育期对应光质的光照射时间,s
Wj——整个生育期生菜地上部分的平均干质量,g/株
D——栽培密度,株/m2
单位面积里生产单位质量的生菜所消耗的电量K(J/g)计算公式为
(12)
3 结果与分析
3.1 交替红、蓝光照射下生菜的LUE和EUE
由图1(图中不同小写字母表示处理间在P<0.05水平差异显著,短线表示标准误差,下同)可知,交替红蓝光照射模式对生菜光能利用率LUE有一定的影响,较红蓝光同时供光的对照RB而言,所有交替处理均显著提高了生菜LUE,提高幅度为34.3%~78.6%。所有处理中,R/B(30 m)下的生菜LUE最高,达到5.84%,R/B(15 m)次之,与最大值无显著性差异;其他处理下LUE均显著小于最大值,其中,纯蓝光处理下LUE为2.26%,显著低于其他任意处理。
由图2可知,较对照RB而言,所有交替处理的生菜EUE均显著提高,提高幅度为34.6%~79.4%。所有处理中,R/B(30 m)下的生菜EUE最高,达到1.92%,R/B(15 m)和R/B(60 m)次之,与最大值无显著性差异。纯蓝光下LUE显著低于其他处理,但EUE却不是最低的,这可能是因为蓝光波长短、光子能量大,造成LUE计算时分母的值最大。
由表3可知,本试验中,每平方米种植面积里,生产单位干、鲜质量生菜所需光量子数最少,分别为1.82、0.08 mol/g,生产单位干、鲜质量生菜所需的耗电量最少,分别为1.04、0.05 MJ/g,且p和K的最小值均出现在R/B(30 m)处理下,最大值是最小值的1.6~2.1倍。与红蓝光同时供光的处理相比,所有交替光处理下生菜地上部鲜质量均有所提高,提高幅度为18.6%~53.6%。
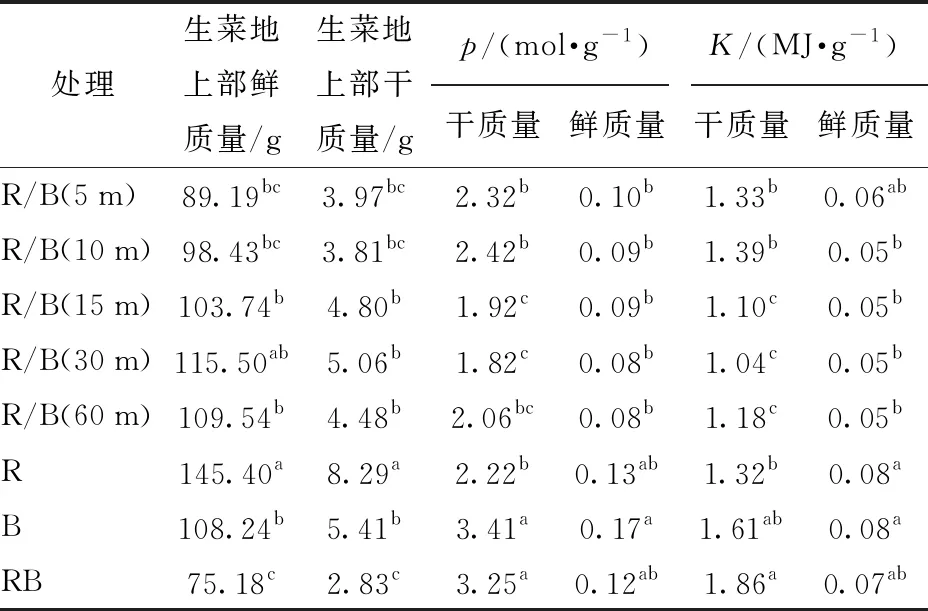
表3 生产单位质量生菜所消耗的光量子数及电量
3.2 交替红、蓝光照射下生菜光合色素含量
图3显示了不同交替红、蓝光照射下,生菜叶片叶绿素a、b以及类胡萝卜素的含量。与红蓝光同时供光的处理RB相比,交替光处理均显著提高了叶绿素和类胡萝卜素的含量;交替光的5个处理之间,红、蓝光交替间隔从5 min到60 min的变化过程中,生菜叶绿素和类胡萝卜素含量均呈现先降低后升高的趋势,R/B(5 m)处理下叶绿素和类胡萝卜素含量均最高,而最低值均出现在R/B(30 m)处理下;在所有处理之间对比可见,纯蓝光下叶片叶绿素a、叶绿素b最高,但R/B(5 m)处理下叶绿素a、叶绿素b含量均与最大值之间无显著性差异,同时,R/B(5 m)处理下类胡萝卜素含量显著高于其他任意处理,这表明,高频次的红蓝光交替可能刺激了类胡萝卜素在叶片中的合成和积累。
3.3 交替红、蓝光照射下生菜叶片的光合及荧光特性
3.3.1不同处理下生菜叶片的光合特性
由表4可见,叶片净光合速率在R/B(30 m)下表现为最大,其次为R/B(60 m)和B处理,三者之间叶片净光合速率无显著性差异;从5 min到30 min的红蓝光交替处理里,随交替间隔的延长,叶片净光合速率有逐渐提高的趋势。纯蓝光下叶片蒸腾速率、胞间CO2浓度以及气孔导度均最大,其中,叶片蒸腾速率显著高于其他任意处理;纯红光下胞间CO2浓度、气孔导度均最低,其中胞间CO2浓度显著低于其他任意处理;所有交替光处理下胞间CO2浓度无显著性差异。所有处理中,叶片光合作用水分利用效率在R/B(30 m)处理下最高,在B处理下最低,最高和最低值与其他处理相比均达到显著水平。总体来看,较红蓝同时供光的对照RB而言,R/B(30 m)处理下生菜叶片净光合速率和水分利用效率均显著提高。
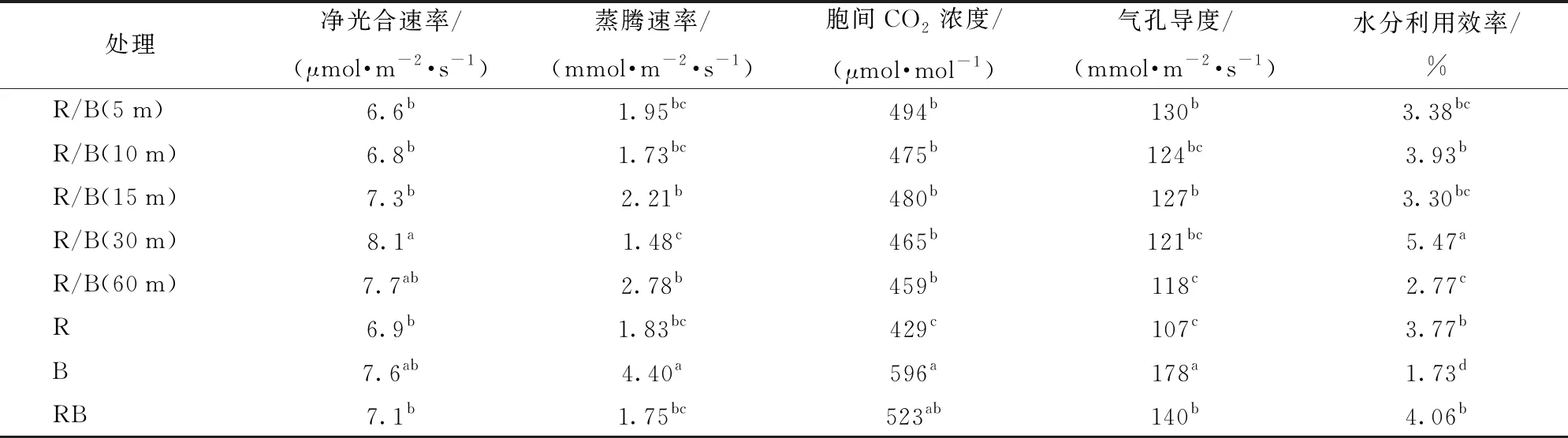
表4 不同交替光处理下生菜叶片光合参数
3.3.2不同处理下生菜叶片的荧光特性
植物体内叶绿素荧光的变化一定程度上反映了环境因子对植物的影响,通过对不同环境条件下快速叶绿素荧光诱导动力学曲线的分析,可以深入探究环境差异对以PSⅡ为主的光合机构的影响以及光合机构对环境的适应机制。
φ反映了暗适应后的最大光化学效率,是能够表现植物生长胁迫和光抑制的一个指标。由表5可见,所有处理下生菜叶片φ均大于0.8,说明不同处理下的植株均未受到生长胁迫。这表明,交替光环境对于生菜植株来说并没有构成光环境胁迫。Vj反映了照光2 ms时PSⅡ有活性的反应中心的关闭程度,Ψo则反映了照光3 ms时PSⅡ有活性的反应中心的开放程度。结果表明,R/B(30 m)和B处理下PSⅡ有活性的反应中心的开放程度最高,R/B(10 m)和R/B(15 m)次之。RC/CSo表示单位面积上的反应中心的数量,结果表明,R/B(30 m)和B处理下RC/CSo最高,二者无显著差异,但显著高于其他处理。PⅠabs是以吸收光能为基础的光合性能指数,能够反映植物光合机构的状态,由于PⅠabs对某些胁迫的敏感度高于φ,因此PⅠabs被认为是能够更好地反映环境对光合机构产生影响的综合指标。结果表明,PⅠabs在纯蓝光处理下最大,显著高于其他处理,而在纯红光处理下最小,显著低于其他处理。这说明,蓝光有利于提高光合机构性能参数; 交替光处理中,15、30、60 min处理下的叶片PⅠabs显著大于5、10 min的处理,这可能说明交替频率太高不利于光合机构的有效运转。总体来看,较红蓝同时供光的对照RB而言,R/B(30 m)处理下生菜叶片Ψo、RC/CSo、PⅠabs均显著提高。
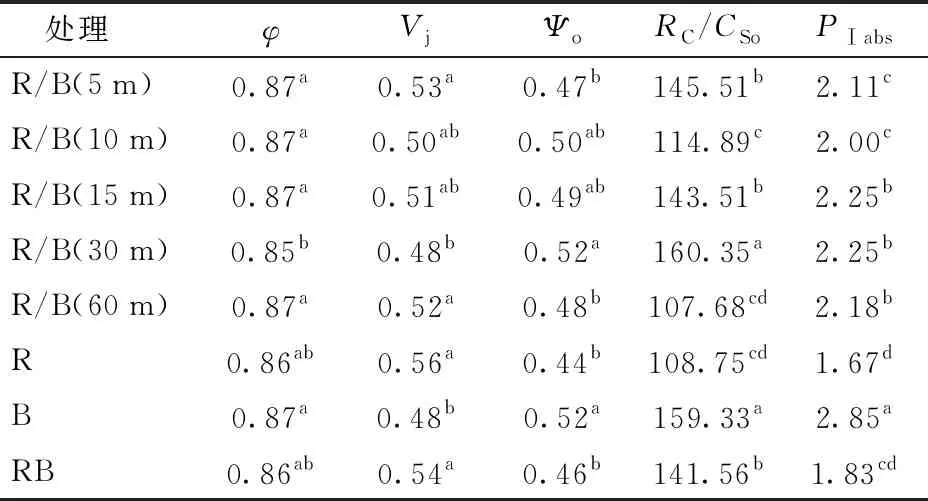
表5 不同交替光处理下生菜叶绿素荧光动力学参数
为了更确切地反映不同光处理下生菜植株光合器官对光能的吸收、耗散、传递以及转化等情况,本研究测定计算了光合机构的比活性,即单位受光面积的各种量子效率,包括单位面积吸收(ABS/CS)、捕获(TRo/CS)、用于电子传递(ETo/CS)和热耗散(DIo/CS)的光能量子效率等。结果表明(表6),R/B(30 m)处理下ABS/CS、TRo/CS、ETo/CS、DIo/CS均最高,尤其是最终用于电子传递的量子效率ETo/CS显著高于其他处理。这可能表明,30 min间隔的交替红光照射有效地提高了整个PSⅡ光合机构的比活性。对比ABS/CS、TRo/CS、DIo/CS与ETo/CS可以发现,大部分处理下,光能的吸收、捕获、传递以及热耗散基本呈现一致趋势,也就是说吸收光能多的处理,一般而言捕获到的光能、用于电子传递的以及热耗散的光能均较高。但是,纯红光处理下,生菜叶片最终用于电子传递的光能量子效率ETo/CS明显低于吸收、捕获和热耗散的相对水平,相反,纯蓝光下ETo/CS则有升高趋势,这表明,与红光相比,蓝光光量子的能量更有利于进入实质的光合电子传递链中。
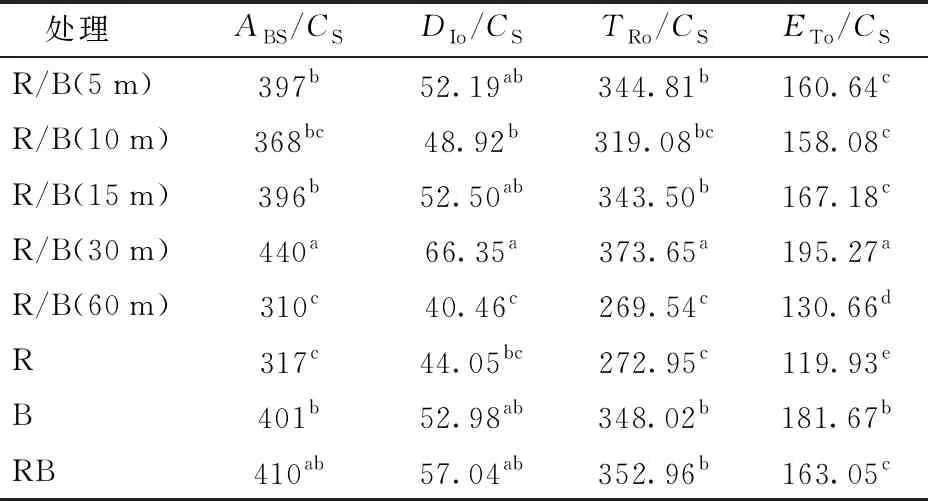
表6 交替红蓝光对叶片PSⅡ反应中心能流分配的影响
4 讨论
与红蓝光同时照射的处理RB相比,所有交替光处理下的生菜地上部鲜质量、LUE、EUE均有所提高,提高幅度与交替间隔时间有关,30 min交替间隔的处理下3个指标均最大,这表明除了光质外,还可以通过交替照射来调节叶片内同化物质的积累。SHIMOKAWA等[17]的研究中,不同R/B交替间隔的生菜鲜质量从大到小顺序为12 h/12 h、3 h/3 h、24 h/24 h、6 h/6 h、1 h/1 h、48 h/48 h。因此,可以推测,交替R/B可能影响生菜的生长,并且结果随交替间隔(即交替频率)不同而异。本试验中,在纯蓝光下生菜叶片净光合速率与最大值无显著差异,且PⅠabs表现为所有处理间最大值,这说明蓝光量子的能量更有利于进入实质的光合电子传递链中,蓝光有利于提高光合机构的性能参数和单个叶片的光合能力,然而蓝光下生菜LUE和EUE却最低或与最低值无显著性差异,这可能是因为蓝光下叶片数少,总叶面积及整体光合能力受到限制。此外,R/B(5 m)处理下生菜叶片类胡萝卜素含量显著高于其他任意处理,高频次的红蓝光交替可能刺激了类胡萝卜素在叶片中的的合成和积累,有利于对光合器官的保护。
在叶绿素荧光分析结果中可知,所有交替光处理下生菜叶片φ均大于0.8,这说明交替变化的光环境对于生菜植株来说并没有构成光环境胁迫。与红蓝光同时供光的对照RB相比,R/B(30 m)处理下生菜叶片净光合速率、水分利用效率、Ψo、RC/CSo、PⅠabs、ABS/CS、TRo/CS、ETo/CS均得到显著提高,该处理下PSⅡ光合机构的比活性整体最高,这可能是该处理下生菜鲜质量、LUE、EUE较大的原因之一。与红蓝光同时供光的对照RB相比,R/B(30 m)处理下生菜叶片净光合速率、水分利用效率、Ψo、RC/CSo、PⅠabs、ABS/CS、TRo/CS、ETo/CS均得到显著提高,该处理下PSⅡ光合机构的比活性整体最高。交替光照射的处理之间,随交替间隔的延长,叶片净光合速率有逐渐提高的趋势。
红光和蓝光通过光受体影响植物的光反应,目前发现的红光受体有5种光敏色素(phyA~phyE),蓝光受体包括3种隐花色素(cry1、cry2、cry3)和2种向光素(phot1、phot2)[25-27]。光受体之间的关系可能是协同的或者拮抗的,这与光环境及其特定的植物生理活动有关,也就是说,R和B的信号转导途径在某些情况下是独立的,但在其他情况下是相互作用的,可能存在光受体信号转导途径的串扰。文献[17]认为,R和B光受体的激活途径在交替模式和同时照射模式之间可能不同,因此,如果R和B之间的光受体响应存在一些冲突,则交替照射可以准确地解决冲突,只要不同的光以适当的间隔(例如,从红光切换到蓝光)照射可能使得单色R或B可以充分发挥其功能而没有负面效应,这可能是交替照射模式下产生栽培益处的一种可能的解释。
5 结论
(1)与红蓝光同时供光的处理RB相比,所有交替光处理下的生菜地上部鲜质量、LUE、EUE均有所提高,其中R/B(30 m)处理下鲜质量、LUE、EUE均最高,分别为115.50g、5.84%、1.92%;纯红光下,生菜地上部生物量最大,但LUE和EUE均显著小于R/B(30 m)处理;纯蓝光下,生菜地上部生物量在处理间居中,但LUE和EUE在处理间最低或与最低值无显著性差异。
(2)所有交替光处理下生菜叶片φ均大于0.8,说明交替变化的光环境对生菜植株并没有构成光环境胁迫。与红蓝光同时供光的对照RB相比,R/B(30 m)处理下生菜叶片净光合速率、水分利用效率、Ψo、RC/CSo、PⅠabs、ABS/CS、TRo/CS、ETo/CS均得到显著提高,该处理下PSⅡ光合机构的比活性整体最高。交替光照射的处理之间,随着交替时间间隔的延长,叶片净光合速率有逐渐提高的趋势。
