氮素添加对荒漠化草原草本植物养分化学计量特征的影响
2021-06-28马英许志豪曾巧红孟建龙胡亚虎苏洁琼
马英,许志豪,曾巧红,孟建龙,胡亚虎*,苏洁琼
(1.兰州大学资源环境学院,甘肃省环境污染预警与控制重点实验室,甘肃 兰州730000;2.甘肃华鼎环保科技有限公司,甘肃 临夏731199;3.中国科学院西北生态环境资源研究院,沙坡头沙漠研究试验站,甘肃兰州730000)
生态化学计量特征可以反映植物对养分元素的吸收利用策略,并能反映植物的生长速率与养分吸收的内在关联[1-2]。近年来,随着全球气候的变化,加之人类的过度干扰,荒漠化草原不断退化,荒漠生态系统的稳定性及其流域生态系统服务功能逐渐降低[3-4]。草本植物是荒漠化草原生态系统的主要植被群落类型,在养分匮乏的条件下,其形成了自身独特的生理生态和养分生态化学计量特征,具有相对稳定的环境适应性[5-6],对草地的退化和恢复具有重要的作用[7]。生态化学计量特征通过影响植物生长、凋落物分解、微生物的相互作用与群落动力学以及养分的循环等来控制草地生态系统的过程[8-9],其与环境之间存在复杂的反馈机制,一旦它们之间的平衡被打破,就会引起生态环境的显著变化[10]。
氮(N)素是控制荒漠化草原草本植物生长的首要养分限制性因子[11]。研究表明,适量的N素添加可以提高荒漠生态系统的生产力,恢复退化的草地生态系统[11-12]。然而,长期或过量的N素添加引起的土壤N素积累可能会造成土壤—植被系统养分计量关系的失衡、生物多样性的降低以及生态系统服务功能的下降等问题[13]。尽管前人的研究已发现草本植物能够通过其养分化学计量调节作用适应变化的环境和维持生态系统的平衡[14],但养分动态及其相互作用的研究多是在非自然群落水平下取得的,且主要集中在土壤、植物地上部器官等方面[15]。因此,目前的研究仍不能全面反映草本植物在荒漠化草原自然植被群落水平下的养分动态响应和反馈机制。
物种属性在决定植物养分化学计量特征方面发挥着重要的作用,对预测未来养分富集情况下植物群落组成的改变具有重要的参考意义[16]。基于上述考虑,本研究以腾格里沙漠东南缘的典型荒漠化草原草本植物为研究对象,通过野外N素添加实验模拟全球性大气N沉降增加导致区域性土壤N素累积的情景,研究N素添加下荒漠化草原不同功能型草本植物根部和地上部碳(C)、N和磷(P)化学计量特征的变化。本研究旨在探明温带荒漠化草原自然植被群落水平下不同属性草本植物的生长机理及养分利用和C分配策略,进而为荒漠生态系统的恢复与管理提供一定的理论和实践依据。
1 材料与方法
1.1 研究区概况
研究区位于宁夏中卫市的沙坡头地区,该地区位于腾格里沙漠东南缘,常年降水量稀少,属于典型的荒漠化草原地带。研究区海拔1250 m,年均气温10.4°C,年均降水量186 mm,年均风速为2.8 m·s-1,每年大于5 m·s-1的风沙日在200 d左右。该地区地下水埋深超过60 m,不能被草本植物有效利用。研究区不同功能型草本植物的优势植物一年生草本为茵陈蒿(Artemisia capillaris)、多年生禾本科为无芒隐子草(Cleistogenes songorica)、多年生非禾本科为狗娃花(Aster hispidus)。
1.2 样地设置
采用随机区组设计,共设定了8个区组,每个区组包含4个2 m×2 m的样方,每个样方之间设有2 m的缓冲带。N素的添加设计了4个水平,分别为:0、3.5、7和14 g N·m-2·yr-1,分别对应不施肥的对照(N0)、低肥(N3.5)、中肥(N7)和高肥(N14)处理,每个区组的4个样方随机接受任一N素添加处理。2018年5和7月,以硝酸铵水溶液的形式将N素(每次投加1/2)均匀喷洒到不同的样方中,其中N0处理喷洒等量的水分。
1.3 样品采集与测定
2018年9 月,每个N素处理随机选择4个区组采集土壤和植物样品。土壤的采集是在每个区组中随机选取3个采样点,用直径3.5 cm的土钻取土壤样品,过2 mm筛后一部分于4°C保存用于植物有效态N素(硝态和铵态N)的测定,一部分风干后用于土壤总N的测定。植物样品按照功能型分类采集,每种功能型的植物均分为根部和地上部,然后装于信封中于75°C烘至恒重后粉碎用于植物全C、N和P的测定。土壤总N采用浓H2SO4+K2SO4+CuSO4的方法消煮,土壤硝态和铵态N采用2 mol·L-1的氯化钾溶液浸提,消煮液和浸提液均采用流动注射分析仪(FIAstar5000 Analyzer,FOSS,丹麦)测定N素的含量[17-18]。植物全N和P的前处理和测定方法同土壤全N的方法,植物全C的测定采用烘箱加热法[19]。
1.4 数据分析
所有数据基于样品干重,采用平均值±标准差(n=4)的形式表示。采用Microsoft Excel和SPSS 22.0软件进行数据的整理与统计分析,采用Origin 8.0软件作图。不同N素处理下土壤全N、硝态和铵态N以及单一植物体内全C、N和P的变化采用一元定量资料的单因素方差分析,不同植物间根部和地上部全C、N和P的变化采用一元定量资料的多因素方差分析。多重比较采用Duncan新复极差法,显著性水平设为P<0.05。
2 结果与分析
2.1 土壤氮素含量随氮肥处理水平的变化
由图1所示,相较于对照处理,土壤中的全N含量在所有施肥处理下均显著高于对照,且其随着土壤中N素添加水平的升高而显著增加(P<0.05)。对植物有效态的N素而言,N素的添加没有显著提高土壤中硝态N的含量,反而是在低(N3.5)、高(N14)N肥处理下显著降低了其含量(P<0.05)。土壤铵态N含量在N素添加水平较高的中(N7)、高N肥处理下显著高于对照和低N处理(P<0.05)。
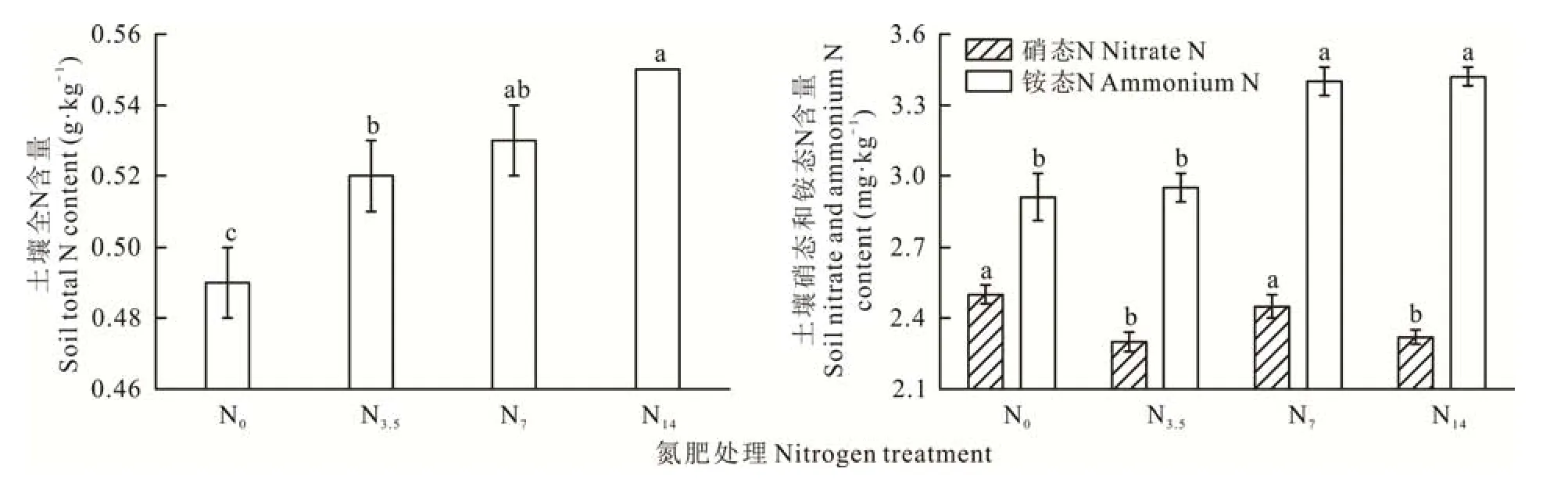
图1 不同N素添加处理下土壤全N和有效态N素含量的变化Fig.1 Changes of soil total N and available N content under different N addition treatments
2.2 植物器官中碳、氮和磷含量随氮肥处理水平的变化
从表1中可以看出,N肥处理、植物类型、植物器官以及它们之间的交互作用对植物体内C、N和P含量的影响均是显著的(P<0.05)。植物对N的吸收和分配如图2所示,N素添加后均会显著提高3种不同属性植物根部和地上部的N含量(P<0.05)。在所有处理下,3种植物地上部的N含量均高于其根部。在低、中和高N肥处理间,茵陈蒿和无芒隐子草根部和地上部的N含量均随着施肥水平的升高而显著增加,但狗娃花仅其地上部N含量的变化是随施肥水平的增加而显著增加的,其根部N含量的变化在3个施肥处理间差异并不显著(P>0.05)。
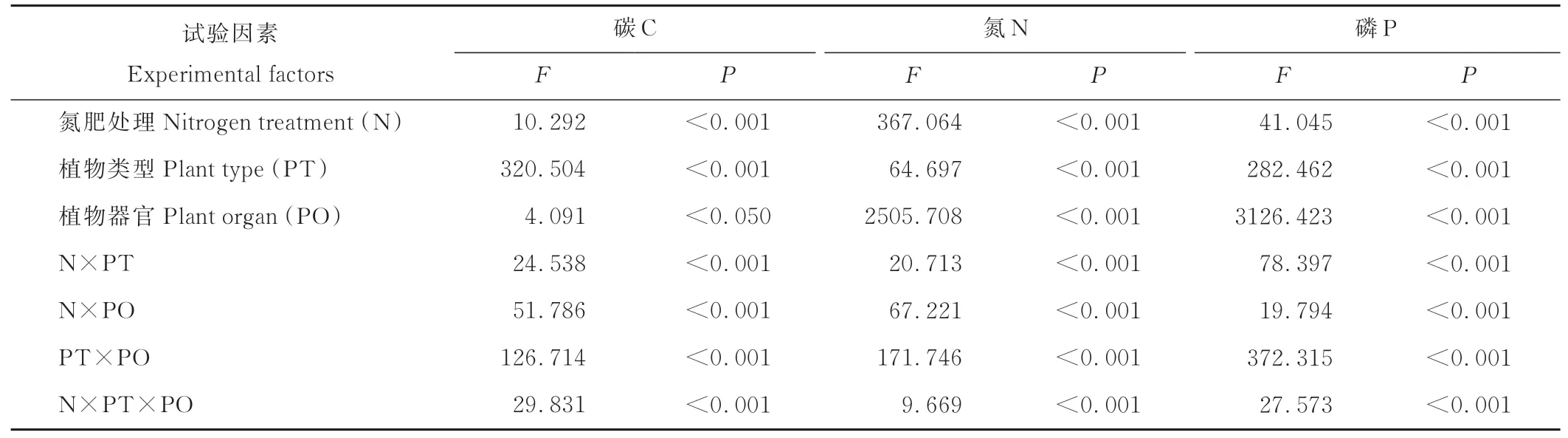
表1 植物C、N、P含量的多因素方差分析Table 1 Results of the univariate multi-factor ANOVA on the content of C,N,and P in the plant
外源添加N素后,3种植物对P的吸收和分配机制截然不同(图2)。具体来看,N素添加后茵陈蒿根部的P含量与对照相比是显著降低的,其地上部P含量仅在低肥处理下是显著高于对照的(P<0.05)。无芒隐子草体内P含量的变化仅在低、高肥处理下与对照相比是显著的(P<0.05),其中根部P含量在低、高肥处理下均是显著增加的,地上部P含量则在低肥处理下显著降低而高肥处理下显著增加。不同于茵陈蒿和无芒隐子草,狗娃花根部和地上部的P含量在所有N素添加处理下均是显著高于对照的(P<0.05),但其根部P含量在所有N素添加处理间却随着施肥水平的升高而显著降低(P<0.05)。
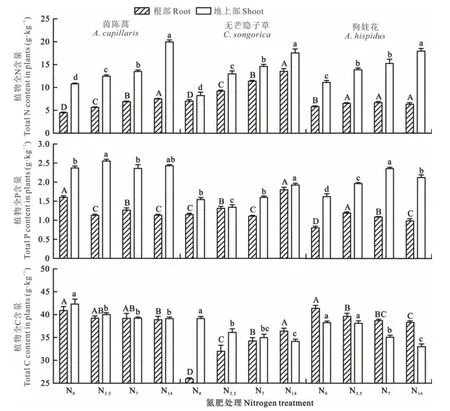
图2 不同N素添加处理下植物根部和地上部C、N、P含量的变化Fig.2 Changes of C,N and P contents in roots and shoots of plants under different N addition treatments
3种植物体内的C含量及其分配格局对外源添加N素的响应亦截然不同(图2)。对茵陈蒿而言,其地上部的C含量在N肥处理下均显著低于对照(P<0.05),但不同N肥处理间的差异并不显著(P>0.05);其根部的C含量仅在高肥处理下显著低于对照(P<0.05)。与对照相比,无芒隐子草根部和地上部C含量的变化呈现出截然相反的趋势,其根部的C含量在施N处理下显著高于对照且随着施肥水平的升高而显著增加(P<0.05),但其地上部的C含量在N素添加处理下却显著低于对照且随着施肥水平的增加而逐渐降低。狗娃花体内的C含量无论是根部还是地上部在施N处理下均是显著低于对照的,且均随着施肥水平的增加而显著降低(P<0.05)。
2.3 不同功能型植物碳、氮、磷化学计量比随氮肥处理水平的变化
3种植物的C/N无论是根部还是地上部在N素添加处理下均是显著低于对照且随着N素添加水平的升高而逐渐降低(P<0.05),其中茵陈蒿和狗娃花相较于无芒隐子草其根部的C/N在同一施肥处理下要远高于其地上部(图3)。对C/P而言,3种植物器官间的差异在不同施肥处理下的表现与C/N相似。随着施肥水平的升高,无芒隐子草和狗娃花地上部C/P的变化是逐渐降低的(P<0.05),但茵陈蒿地上部C/P的变化则不显著(P>0.05)。相反,施肥会显著提高茵陈蒿根部的C/P,低、中肥也会显著提高无芒隐子草根部的C/P,狗娃花根部的C/P也是随着施肥水平的增加而显著增加的,尽管其值在所有施N处理下是低于对照的(P<0.05)。对3种植物的N/P而言,无论是根部还是地上部,其值均小于12。整体上来看,施肥可以显著提高3种植物的N/P,但其随施肥水平升高而逐渐增加的趋势在茵陈蒿的根部和地上部表现的更加突出(P<0.05)。对狗娃花而言,仅高肥才能显著提高其地上部的N/P,同时施肥会抑制其根部N/P的增加,但这种效应随着施肥水平的增加会逐渐得到缓解(P<0.05)。
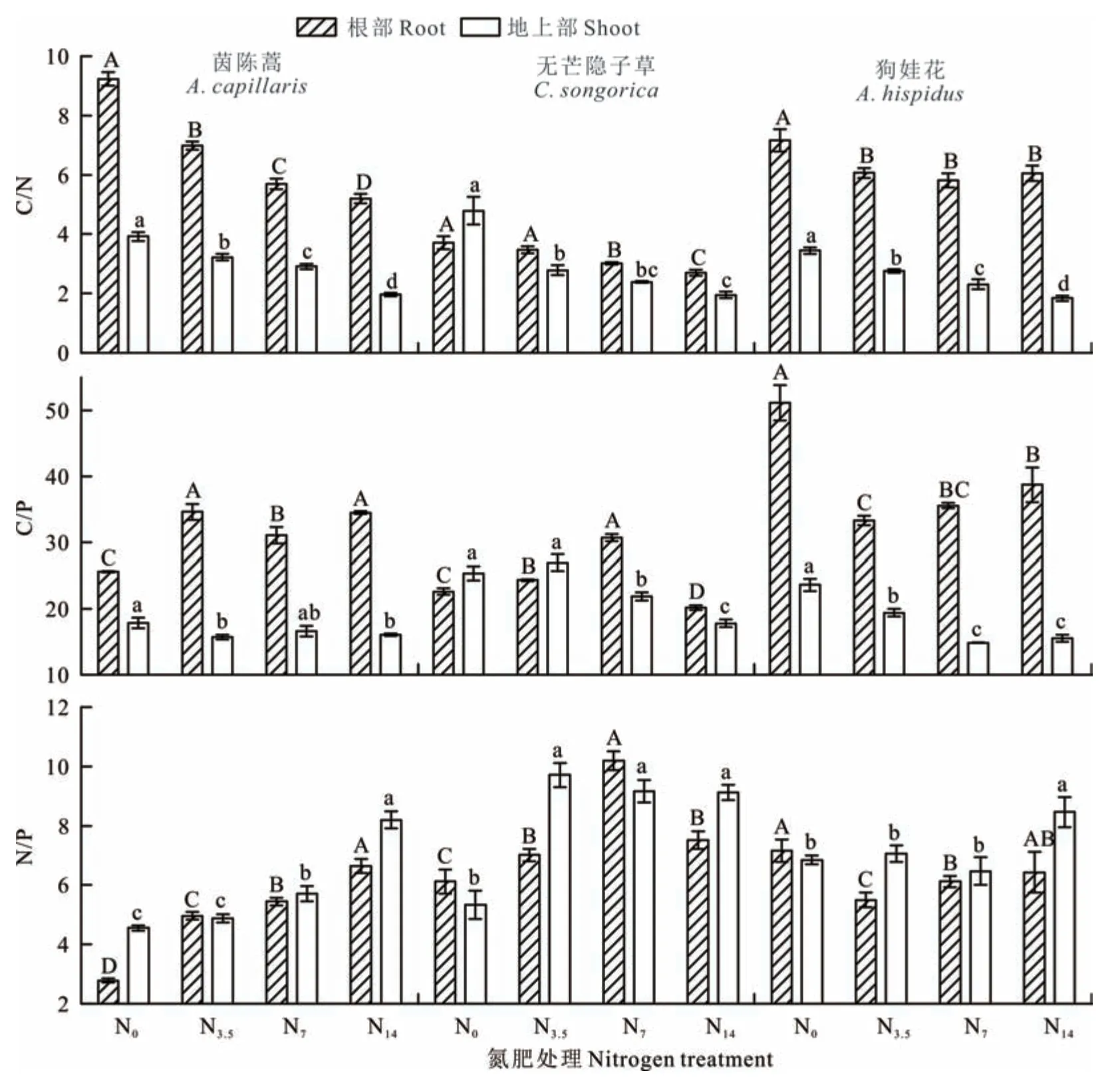
图3 不同N素添加处理下植物体内C、N、P生态化学计量比的变化Fig.3 Changes of ecological stoichiometry of C,N,and P in plant roots and shoots under different N addition treatments
3 讨论
3.1 氮素添加对土壤氮素水平与植物养分吸收及碳分配的影响
荒漠化草原在添加N素后土壤中总N和有效态N的含量均会显著增加[20]。然而,本研究却发现添加N素导致土壤中总N含量的增加并不会带来表层土壤中硝态N含量的显著增加(图1),其原因可能与研究区较低的植被覆盖条件下砂质土壤有利于硝态N的淋失有很大的关系[21]。相反,土壤中铵态N的含量却随施肥水平的升高而逐渐增加(图1),这意味着长期外源输入N素后,土壤中铵态N的积累可能会导致植物根际微环境的变化,如土壤的酸化、P有效性的增加以及C激发效应的减弱等[20,22-23]。
N肥处理、植物类型、植物器官以及它们的交互作用对植物体内C、N、P的含量均产生了显著影响(表1),这说明研究区荒漠草本植物对土壤N素水平变化的响应存在明显的植物特异性。通常,植物地上部的新陈代谢要比根部旺盛,因而地上部的养分需求要比根部高[24]。具体来看,外源添加N素后3种植物根部和地上部对N的吸收均会显著增加,且所有处理下地上部的N含量始终高于根部,但植物对N素吸收随施肥水平升高而显著增加的趋势在多年生非禾本科植物狗娃花的根部并未出现(图2)。这说明,荒漠草本植物的功能型不同,其对养分的利用策略存在明显差异[16,25]。
荒漠化草原土壤呈弱碱性,P容易与钙(Ca)结合形成Ca-P沉淀,这大大降低了P的有效性[26]。本研究中外源N肥的添加导致土壤中铵态N含量增加,这会导致植物根际土壤的酸化[20,27],进而诱发Ca-P结合态P的溶解释放,从而提高土壤中P的有效性并促进植物对P的吸收[28]。从3种植物对P的吸收来看,本研究中外源N素的添加对P吸收的促进作用集中体现在多年生草本植物上,其中多年生非禾本科植物狗娃花根部和地上部的P含量在所有施N处理下均显著高于对照处理,多年生禾本科植物无芒隐子草对P的吸收在高肥处理下是显著增加的(图2)。对一年生植物茵陈蒿来说,添加N素反而显著抑制了其根部对P的吸收,同时添加N对地上部P吸收的促进作用则十分有限(图2)。根据来自昆仑山荒漠草原和黄土高原的野外大田施肥试验表明,外源N素的添加对草本植物P素的吸收会产生中性或者抑制的效应[25,29]。很明显,在P有效性长期受限的荒漠化草原,土壤N有效性的增加及其对P有效性的提升会协同促进养分利用策略以保守为主的多年生草本植物的生长,尤其是非禾本科的多年生草本植物[23,30]。
植物体内的C含量是表征植物固C能力的重要指标,其反映了植物适应外界环境变化和维持体内C平衡的能力[31]。郭美玲等[32]在高寒草甸的研究发现,N肥处理会增加草本植物叶片的C含量。高宗宝等[16]在呼伦贝尔草甸草原的研究显示,N肥处理对4种优势草本植物的全C含量并无显著影响。本研究中,3种植物体内(除无芒隐子草的根部)的C含量在添加N后均降低,且N素添加水平越高降幅越明显,同时多年生植物的下降趋势比一年生植物更明显(图2)。这说明,在养分受限的荒漠化草原,外源N素的输入会削弱荒漠草本植物的固C能力。相较而言,一年生植物能够快速有效地吸收外源输入的养分完成其生活史,因而其显示出了较高的C同化能力[33]。需要特别指出的是,多年生禾本科植物无芒隐子草根部的C含量随着施肥水平的升高而显著增加,且在高肥处理下其值甚至超过了地上部(图2)。无芒隐子草在土壤中N有效性持续增加的情形下不断加大其根部C的分配,其与N素添加下植物通常会增加根部C的分配情形相同[34],这说明拥有庞大发达的根系是荒漠化草原多年生禾本科植物维持其生活史的重要机制,同时加强根部的C分配也是其调控外界环境变化,保障其正常生长的根本原因[35-36]。
3.2 植物碳、氮、磷化学计量比的变化及其指示意义
植物组织中C、N、P的化学计量比与植物生长密切相关,其对阐明元素生物地球化学与生态过程对全球变化的响应至关重要[14,37]。根据在欧洲基于40个独立人工外源施加N、P肥的试验,当N/P<14时,N为限制性因子;N/P>16时,P为限制性因子;而14<N/P<16时,N、P则同为限制性因子[38]。在本研究中,3种荒漠草本植物的N/P在所有施肥处理下均小于14(图3),说明N素的确是荒漠化草原草本植物生长的首要养分限制性因子[11]。通常,随着外源N素的添加,植物组织的C/N会降低,但N/P会升高[37]。在本研究中3种植物的C/N随着外源N素添加水平的升高而降低,N/P则相应地在增加,但一年生植物茵陈蒿的C/N和N/P变化趋势较多年生植物则更加明显(图3)。土壤中N有效性提升情形下植物体较高的C/N意味着其对N素的吸收利用快速而高效[39-40]。因而,荒漠化草原外源N素添加后其对草本植物尤其是一年生植物的生长有显著的促进作用。
外源N素的添加尽管缓解了荒漠化草原N的限制并在一定程度上提升了土壤中P的有效性,但却打破了植物的养分需求平衡,土壤中N/P化学计量比的增加必然迫使P的限制性作用进一步凸显[41-42],其反而会抵消N有效性增加对植物生长的促进作用[43]。相较于多年生草本植物,一年生草本植物茵陈蒿的生长需要更高P的投入(图3),外源N素添加后其根部和地上部的N/P均随着施肥水平的升高而显著增加,同时其根部C/P在施肥处理下显著高于对照,而其地上部C/P在施肥处理下随施肥水平的升高而降低的趋势却并不显著(图3)。这些变化说明外源N素输入解除荒漠化草原N素限制后,P有效性会成为荒漠草本一年生植物生长的重要限制性因子。
植物地上部组织的C/N和C/P反映了植物在吸收营养元素时对C的同化能力,其不仅取决于土壤中养分的有效性,在荒漠化草原其同时还取决于土壤水分的有效性[39,44]。本研究中3种植物地上部的C/N和C/P在施N后整体上是下降的,且施N水平越高下降趋势越显著(图3),这表明荒漠化草原N素的输入会降低植物的C同化能力。从3种植物根部的C/N和C/P来看,一年生植物茵陈蒿和多年生非禾本科植物狗娃花在所有处理下其值均远高于地上部,多年生禾本科植物无芒隐子草则在中、高肥处理下也会表现出这种趋势(图3)。然而,从N/P来看,3种植物在所有处理下均未表现出显著的器官差异,这意味着荒漠化草原外源N素的输入会加强草本植物体内C向根部的分配,其可能是为了增加植物对P的获取和吸收,亦可能是为了维持和拥有更加庞大发达的根系系统。
4 结论
在荒漠化草原,外源N素的添加显著增加了土壤及3种不同功能型草本植物体内全N的含量,且施肥水平越高效果越显著。外源N素添加后,其会促进多年生草本植物尤其是非禾本科植物对P的吸收,但会降低除多年生禾本科植物根部外3种植物体内的C含量。相应地,外源N素添加降低了植物体内的C/N和地上部的C/P,N/P的增加一年生植物更明显,且所有化学计量比的变化随施肥水平越高变化趋势越显著。另外,外源N素添加后3种植物根部的C/N和C/P总是高于地上部,但它们的N/P并未表现出显著的器官差异。可见,不同功能型草本植物对养分的利用存在明显的差异。外源N素的输入会促进荒漠化草原草本植物的生长,但当N素的限制性作用解除后一年生草本植物的生长会受到P素的限制,同时荒漠草本植物的C同化能力也会减弱,但根部的C分配作用会加强。本研究结果可以为大气N沉降增加背景下基于草本植物群落养分利用策略和C分配机制维持荒漠化草原生态系统的稳定性和保障其可持续发展提供重要的理论支撑。
