Effects of long-term straw incorporation on nematode community composition and metabolic footprint in a rice-wheat cropping system
2021-06-24CHENYunfengXlAXiangeHUChengLlUDonghaiQlAOYanLlShuanglaiFANXianpeng
CHEN Yun-feng,XlA Xian-ge,HU Cheng,LlU Dong-hai,QlAO Yan,Ll Shuang-lai,FAN Xian-peng
Key Laboratory of Fertilization from Agricultural Wastes,Ministry of Agriculture and Rural Affairs/Institute of Plant Protection and Soil Fertilizer,Hubei Academy of Agricultural Sciences,Wuhan 430064,P.R.China
Abstract Soil nematode communities can provide valuable information about the structure and functions of soil food webs,and are sensitive to agricultural practices,including short-term straw incorporation. However,currently,such effects under longterm straw incorporation conditions at different fertility levels are largely unknown. Thus,we conducted a 13-year ongoing experiment to evaluate the effects of long-term straw incorporation on the structure and functions of the soil food web in low and high fertility soils through analyzing its effects on nematode communities,food web indices and metabolic footprints. Four treatments were included:straw removal (-S) under non-fertilized (-NPK) or fertilized (+NPK) conditions;and straw incorporation (+S) under -NPK or +NPK conditions. Soil samples from a 0-20 cm depth layer were collected when wheat and rice were harvested. Compared with straw removal,straw incorporation increased the abundances of total nematodes,bacterivores,plant-parasites and omnivores-predators,as well the relative abundances of omnivores-predators with increases of 73.06,89.29,95.31,238.98,and 114.61% in -NPK soils and 16.23,2.23,19.01,141.38,and 90.23% in +NPK soils,respectively. Regardless of sampling times and fertilization effects,straw incorporation increased the diversity and community stability of nematodes,as indicated by the Shannon-Weaver diversity index and maturity index. Enrichment and structure index did not show significant responses to straw incorporation,but a slight increase was observed in the structure index. The analysis of nematode metabolic footprints showed that straw incorporation increased the plant-parasite footprint and structure footprint by 97.27 and 305.39% in -NPK soils and by 11.29 and 149.56% in +NPK soils,but did not significantly influence enrichment,bacterivore and fungivore footprints. In conclusion,long-term straw incorporation,particularly under a low fertility level,favored the soil nematodes and regulated the soil food web mainly via a top-down effect.
Keywords:soil nematodes,community composition,metabolic footprint,straw incorporation,rice-wheat cropping system,top-down effect
1.lntroduction
The annual crop straw production in China has been estimated to be nearly 1 billion tonnes,accounting for 25% of the total straw production in the world (Tianet al.2018).Currently,the utilization of crop straw in China has been mainly focused on forage,straw return (mainly including straw incorporation and mulching),energy,base material for edible mushrooms,and industrial processing,such as papermaking,packing material,artificial boards,etc. Of the five types of crop straw utilization,straw return accounted for nearly half of the total straw utilization (Shiet al.2017),and has been regarded as the most important alternative pathway to burning straw,which has led to serious environmental pollution in recent decades in China. The rice-wheat rotation system is the most dominant cropping system at the middle and lower flow regions of Yangtze River Plain in Central China,and the yearly high straw yields of rice and wheat straw are about 9.0 and 4.5 t ha-1,respectively (Chenet al.2017). Since over 60% of rice and wheat straws were directly incorporated into the soil in the 2010s (Liu and Li 2017),the rice-wheat cropping system has contributed substantially to the environmental and economic benefits.
Crop straw is a valuable resource with a high carbonto-nitrogen ratio (C/N ratio),and is rich in mineral nutrients with respect to potassium (K). Hence,straw return has been applied to improve the soil nutrients and productivity (Chen Z Met al.2018;Songet al.2018),and to provide substrates for below-ground soil food webs. Straw decomposition is a complex process predominantly mediated by soil microorganisms (Zhao and Zhang 2018). Currently,most studies have focused on how straw return affects the soil microbial community (Zhaoet al.2016,2017,2019;Chenet al.2017;Chen Y Fet al.2018;Zhao and Zhang 2018;Baiet al.2019;Kouet al.2020). Specifically,both short-term (≤5-year) and long-term straw returns have increased the soil bacterial and fungal biomass,but they have not shown the consistent influences on microbial diversity.
The carbon (C) and energy derived from the incorporated straws deliver all along the trophic levels,and affect not only soil microbes but also the other soil organisms,including nematodes. In soils,nematodes are present at all trophic levels and have diverse feeding preferences and life-history strategies. They could serve as a model soil food web. A series of evaluative indices based on the relative abundances of nematode feeding and life-history groups,such as maturity index (MI),basal index (BI),enrichment index (EI),structure index (SI) and channel index (CI),have been developed to infer the conditions and the environment of soil food webs (Bongers 1990;Ferriset al.2001;Bongiornoet al.2019). Furthermore,for providing information about the magnitude of ecosystem functions (e.g.,utilization of C) and services (e.g.,nutrient mineralization),Ferris (2010) developed the nematode metabolic footprint (NMF). The NMF refers to the amount of C used for growth and reproduction of nematodes and for respiration by different trophic groups,which can provide more detailed interpretation on the energy channels,nutrient release and top-down regulatory functions.
The quality and quantity of exogenous organic materials regulate the soil food web structure and function (Holtkampet al.2008). The readily-available nutrients of organic materials are most likely to be exploited by the organisms within the bacterial channel,such as bacteria,protozoa and bacterivore nematodes. Recalcitrant resources are most likely to be exploited by the organisms within the fungal channel,such as fungi and fungivore nematodes. Crop straw can provide simultaneously readily-available and recalcitrant resources,and the quantity of substrates entering the soil food web in a specific agricultural ecosystem is determined by the amount of straw return,returning time,soil characteristics and agricultural managements (Zhao and Zhang 2018). Many researchers have reported the utilization of nematode community analysis to indicate the effects of straw return on the structure and functions of soil food webs (Ferris and Matutue 2003;Ferriset al.2004;Liet al.2009,2018;Zhang X Ket al.2012;Yeet al.2013;Huaet al.2014;Liuet al.2015;Mouet al.2017;Chen Y Fet al.2018;Kouet al.2020). However,more work is needed. Firstly,other than straw return increasing the abundance of nematodes,most results relative to the nematode functional groups and ecological indices have shown considerable discrepancy even among the similar studies. This discrepancy is probably due to the fact that most of these studies were based on a short-term field experiment. The quality and quantity of substrates for the soil food web were not stable in a short-term period since the decomposition rate of straw is relatively low (Yanet al.2019). Secondly,most of these studies were conducted in soils amended with mineral fertilizer,i.e.,at high fertility level. Because the bottom-up effect and top-down effect (predation and trophic cascade) were found to be weaker in high background fertility soils than in low background fertility soils (Lenoiret al.2007;Chenet al.2017),changes in some nematode communities resulting from straw return might be not observed. In addition,mineral fertilizers at higher concentrations are toxic to the sensitive organisms which inhabit those with small volumes of water (Ferriset al.2012),and these unfavorable effects could offset or even suppress the positive influences of straw return. Thirdly,until now,relatively few studies (Zhang X Ket al.2012;Zhonget al.2017;Kouet al.2020) have examined the nematode metabolic footprints under straw return conditions,which leads to deficiencies relating to the effects of straw return on soil food web energy channels and regulation modes.
The different findings made in various previous studies indicate that the effects of straw return on soil nematode communities need to be investigated in more detail. Thus,this study aimed to determine the effects of long-term straw incorporation under non-fertilized and fertilized conditions on:(1) nematode communities;(2) structure and function of soil food webviaevaluation with nematode fauna analysis;and (3) the regulation mode of the soil food web. We assumed that the nematode abundance and the functions of the soil food web could be influenced positively by bottomup control due to the addition of organic substrates into soil with the straw incorporation,and the C entering the fungal channel could be increased because of the higher C/N ratio of the straw,which might be relatively favorable to fungi. Also,we assumed that the degree of improvements could be higher in the non-fertilized soils than in fertilized soils due to low background fertility. Herein,we report our findings from a 13-year field experiment under a rice-wheat cropping system in Central China.
2.Materials and methods
2.1.Site description
The study site was at Qianjiang Scientific Observing and Experimental Station of Agro-Environment and Arable Land Conservation (112°38´13.21´´E,30° 22´54.07´´N),Ministry of Agriculture and Rural Affairs of China,Hubei Province,China. The soil at this site is classified as Fluo-aquic soil. The long-term experimental field was initiated in the rice season in June 2005,and had been rotated with rice (OryzasativaL.) and wheat (TriticumaestivumL.). Rice was planted from June to October while winter wheat was planted from November to May. At the beginning of the experiment,the soil characteristics were determined as follows:pH,7.1;soil organic carbon content,11.9 g kg-1;total nitrogen (N),1.53 g kg-1;available N (AN),121 mg kg-1;available phosphorus (AP),19.2 mg kg-1;available potassium (AK),59.1 mg kg-1;and bulk density,1.20 g cm-3.
2.2.Experimental design and management practices
The experiment was designed with four treatments and four replications,laid out in a randomized complete block design with 16 plots,and each plot was 5.0 m×4.0 m in size. The four treatments included:(1) straw removal (-S) without nitrogen-phosphorus-potassium (NPK) fertilizer,-S-NPK;(2) straw removal plus NPK fertilizer,-S+NPK;(3) straw incorporation (+S) without NPK fertilizer,+S-NPK;and (4) straw incorporation plus NPK fertilizer,+S+NPK. The experiment was similar to a split-plot experiment,with straw considered as the main plot-factor and fertilization considered as the sup-plot factor. The two treatments of straw removal in -NPK and +NPK soils were considered as Control 1 and Control 2,respectively (Appendix A).
Application rates of crop straw and chemical fertilizers for the four treatments are listed in Appendix A. In each plot,the stubbles those were less than 10 cm and roots were left in the field and the crop straws were harvested. Prior to returning the straw,the rice straw was cut into < 5-cm lengths while the wheat straw was cut into 6-10 cm lengths and piled at 40-80 cm high. For every 20 cm height,the straw was inoculated with 5% (w/w) effective microorganisms (composed withBacillussubtilis,AspergillusnigerandTrichoderma),1% (w/w) urea,5% (w/w) rice bran,0.25% (w/w) sugar and water. After 14 days for rice straw or 21-28 days for wheat straw,the decomposed straw was incorporated into the soil. In the rice-growing season,the N fertilizer was applied at sowing and tillering stages at the rates of 60 and 40% before 2012,and at the sowing,tillering and jointing stages at the rates of 60,20 and 20% from 2012. Urea,superphosphate and potassium chloride were used as the sources of N,P and K,respectively. During the wheat-growing season,the N fertilizer was applied at sowing and tillering stages at the rates of 60 and 40% in all years. The P2O5and K2O were applied as base fertilizers in both rice and wheat seasons.
2.3.Soil sampling and analysis
Soil samples were collected on 26 May 2018 after harvesting wheat and on 7 October 2018 after harvesting rice. In each plot,10-15 soil cores (2.0 cm in diameter) at 0-20 cm depth layer were mixed as one composite sample and passed through a 2-cm sieve to remove stones and plant debris. Sub-samples of 200 g were air-dried and used for subsequent analyses of pH and the contents of soil organic carbon (SOC),AN,ammonium N (NH4+-N),nitrate N (NO3--N),AP and AK. Sub-samples of 300 g were stored at 4°C and used for subsequent analyses of nematodes.
The soil pH and the contents of SOC,AN,AP and AK were determined using the conventional methods (Bao 2005). NH4+-N and NO3--N were extracted with 1 mol L-1KCL solution and then detected with a Continuous Flow Aanlyzer (Technicon Corporation,NY,USA).
Nematodes were extracted from 100 g fresh soil by a modified cotton-wool filter method (Lianget al.2009).After counting the total number of free-living nematodes in a sample,100 individuals per sample were randomly selected and identified to the genus level using a compound microscope under 100× or 400× magnifications. According to the feeding habits,nematodes were assigned into four functional groups:bacterivores (BF),fungivores (FF),plantparasites (PP) and omnivores-predators (OP) (Yeateset al.1993).
2.4.Ecological indices
A number of descriptive and evaluative indices for soil nematodes were used in this study.
Shannon-Weaver diversity index (H´)H´=-∑pi(lnpi),wherepiis the proportion of theith nematode genus.
Maturity index (Ml)MI=∑vipi,whereviis the colonizerpersister (cp) value,assigned by Bongers (1990),for freeliving nematodes of theith genus.
Enrichment index (El) and structure index (Sl)Both EI and SI are calculated from weighted basal,enrichment and structural components (b,e,s) of the nematode assemblage:b=(Ba2+Fu2)×W2,where W2=0.8;e=(Ba1×W1)+(Fu2×W2),where W1=3.2;s=(Ban×Wn+Fun×Wn+Omn×Wn+Prn×Wn),wheren=3-5,W3=0.8,W4=3.2,W5=5.0. Ba,Fu,Om,and Pr indicate the abundances of bacterivores,fungivores,omnivores and predaceous nematodes,respectively,andnindicates the cp value of the nematode taxa. Both EI and SI were calculated from the weighted faunal components:EI=100×[e/(e+b)],SI=100×[s/(b+s)] (Ferriset al.2001).
Nematode metabolic footprint (NMF)NMF=∑{Nt×[0.1×(Wt/mt)+0.273×(Wt0.75)]} (Ferris 2010),where Nt,Wt,and mtrepresent the number,body weight and cp values of genust,respectively,while the Wtis available at http://nemaplex.ucdavis.edu/. The bacterivore footprint (BFfoot),fungivore footprint (FFfoot) and plant-parasite footprint (PPfoot) represent the metabolic footprints of these three trophic groups. The enrichment footprint (Efoot) and structure footprint (Sfoot) represent the metabolic footprints of lower (cp 1-2) and higher (cp 3-5) trophic levels,respectively. Functional metabolic footprints (FMP) were calculated as (Fs×Fe)/2 with complex μg2units,where Fs is the sum of standardized C labile by structural indicator taxa and Fe is the sum of standardized C labile by enrichment indicator taxa (Ferris 2010).
2.5.Data analysis
In this study,there were three types of effects:straw incorporation,fertilization and sampling time. However,only the straw incorporation effect was considered. Independent Student’st-test was used to compare the differences in the measured variables between -S and +S treatments under -NPK and +NPK soils at the sampling times of May and October. Two-way analysis of variance (ANOVA) was used to test the straw incorporation effect regardless of sampling time effect,while three-way ANOVA was used to examine the straw incorporation effect regardless of sampling time and fertilization effects. Prior to statistical analyses,log10(x+1),1/xor square root transformation was used for the original data to satisfy the equal variance assumption.
Plevels of 0.10 or less (0.05,0.01,and 0.001) were regarded as statistically significant. Thet-test,two-way and three-way ANOVA were conducted with SPSS 19.0 (SPSS Inc.,Chicago,IL,USA). The relationships between nematode genera and soil properties were examined based on redundancy analysis (RDA) using CANOCO for Win dows 4.5 package (Microcomputer Power,Ithaca,NY,USA).Monte Carlo permutation tests were applied to calculate statistical significance (n=499).
3.Results
3.1.Soil physicochemical properties and yield of wheat and rice
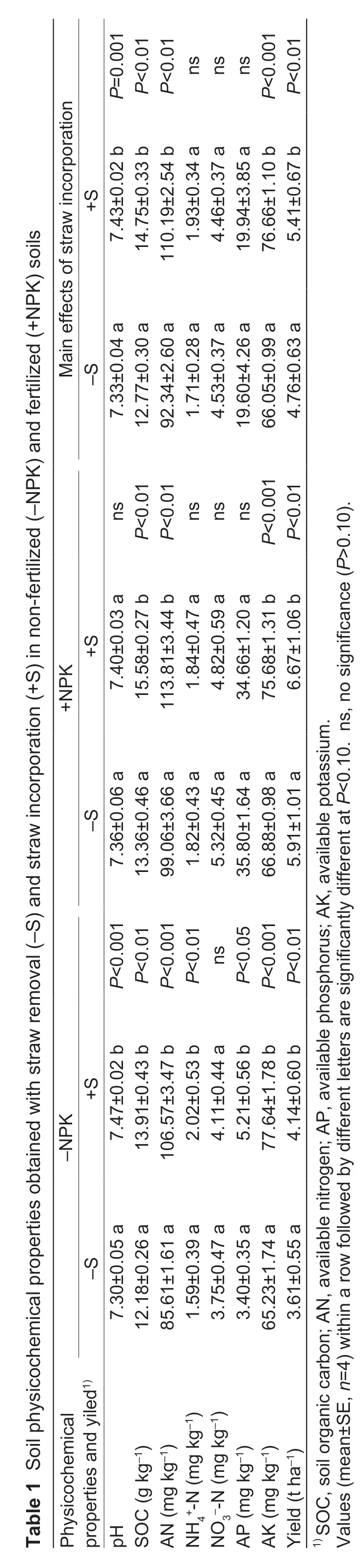
In the low fertility soils,i.e.,-NPK soils,straw incorporation significantly increased most of the selected physicochemical properties (pH,SOC,AN,AK and wheat yield of the sampling of May,and SOC,AN,NH4+-N,AP,AK and rice yield of the sampling of October),and tended to increase other selected properties as well (Appendix B). Regardless of sampling time effect,the pH,SOC,AN,NH4+-N,AP,AK,and crop yield were increased by 2.21,14.20,24.28,26.90,53.39,19.03 and 14.51%,respectively (P<0.001,P<0.01,P<0.001,P<0.01,P<0.05,P<0.001,andP<0.01,respectively) (Table 1).
In the high fertility soils,i.e.,+NPK soils,the straw incorporation effects were similar to those of low fertility soils (-NPK) at both sampling times (Appendix B). Regardless of sampling time effects,straw incorporation increased the contents of SOC,AN,and AK and yields by 16.62,14.89,13.17 and 12.97%,respectively (P<0.01,P<0.01,P<0.001,andP<0.01,respectively) (Table 1). In addition,the degrees of improvement (except for SOC) were higher in -NPK soils than in +NPK soils.
Regardless of sampling time and fertilization effects,except for NO3--N and AP,straw incorporation increased the pH,SOC,AN,NH4+-N,AK and crop yield by 1.35,15.46,19.33,13.10,16.06 and 13.56%,respectively (P=0.001,P<0.001,P<0.001,P<0.10,P<0.001,andP<0.001,respectively) (Table 1).
3.2.Nematode abundances and community compositions
The abundances of total nematodes and four trophic groups are listed in Table 2 and Appendix C. In the -NPK soils,the abundances of total nematodes,bacterivores,plant parasites and omnivores-predators were significantly higher in the +S treatment than in the -S treatment by 80.30,103.36,104.75,and 315.82% at the sampling of May,respectively (P<0.01,P<0.01,P<0.01,andP<0.01,respectively). The same trends were observed at the sampling of October,but the increases in these parameters were not statistically significant. Regardless of the sampling time effect,straw incorporation increased the total nematodes,bacterivores,plant parasites and omnivores-predators by 73.06,89.29,95.31 and 238.98%,respectively (P=0.001,P<0.001,P<0.01,andP<0.01,respectively).

In the +NPK soils,straw incorporation effects on omnivores-predators in May and on fungivores in October were observed,which were increased by 143.26 and 108.93%,respectively (P<0.05 andP<0.05,respectively). Regardless of sampling time effect,straw incorporation tended to increase the total nematodes,bacterivores and plantparasites,and significantly increased the fungivores and omnivores-predators by 12.57 and 141.38%,respectively (P<0.01 andP<0.01,respectively). The degrees of improvements in total nematodes,bacterivores,plant parasites and omnivorespredators were higher in -NPK soils than in +NPK soils.
Regardless of sampling time and fertilization effects,straw incorporation increased the abundances of total nematodes,bacterivores,plant parasites and omnivores-predators by 75.19,89.29,95.31 and 238.98%,respectively (P<0.01,P<0.01,P<0.01,andP<0.001,respectively).
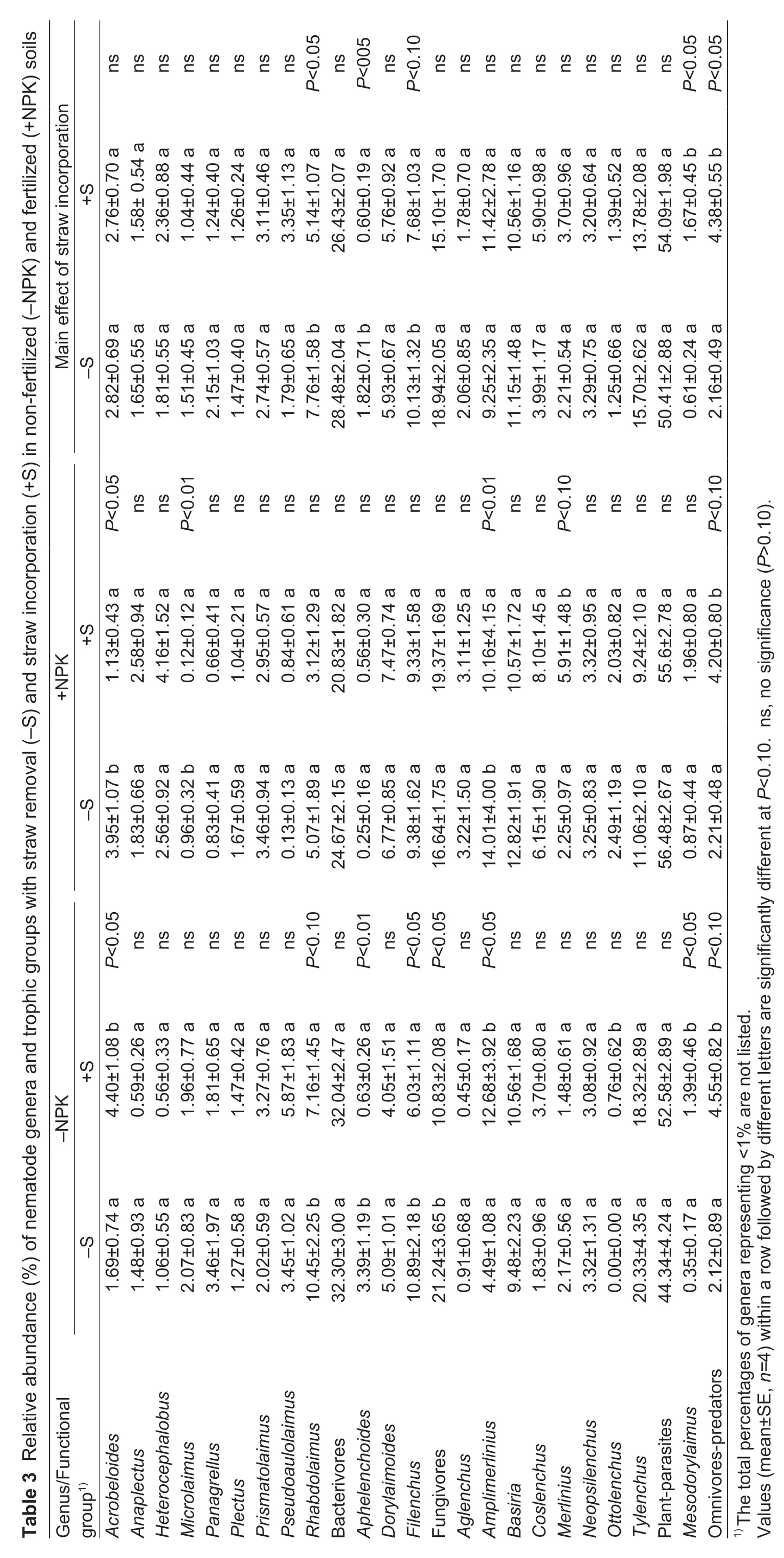
A total of 48 nematode genera were observed at the two sampling times,and only those genera with more than 1% relative abundance are listed in Table 3 and Appendix D. Among them,20 genera were bacterivores,five were fungivores,13 were plant parasites and 10 were omnivorespredators. Of all the nematodes,Tylenchus,BasiriaandAmplimerliniuswere found to be the dominant genera,with relative abundances of 14.74,10.86 and 10.33%,respectively.
Of the four nematode groups,the relative abundance of plant-parasites was the highest (52.25%),followed by that of bacterivores (27.46%). Regardless of sampling time effect,straw incorporation increased the proportion of omnivorespredators by 114.61% in -NPK soils and by 90.23% in +NPK soils,respectively (P<0.10 andP<0.10,respectively). On the other hand,straw incorporation decreased the proportion of fungivores by 49.02% in -NPK soils (P<0.05). Regardless of sampling time and fertilization effects,straw incorporation increased the proportion of omnivorespredators by 102.16% (P<0.05).
3.3.The ecological indices of nematodes
The effects of straw incorporation on the diversity and functional indices of soil nematodes are listed in Table 4 and Appendix E. In the -NPK soils,straw incorporation significantly increased the MI value by 8.39% (P<0.10),whereas it significantly decreased the EI value by 35.73% in May (P<0.05). In October,the effects of straw incorporation on all indices were not significant.Regardless of sampling time effect,straw incorporation increased all indices,except EI,but the increasing effects were not statistically significant.
In the +NPK soils,no significant effects of straw incorporation on any of the indices were observed at either sampling time,but straw incorporation tended to increase the SI value. Also,no significant effects of straw incorporation on any of the indices were observed regardless of sampling time effect.However,straw incorporation significantly increased the values of H´ and MI by 4.49 and 4.35%,respectively (P<0.10 andP<0.10) regardless of sampling time and fertilization effects.
3.4.Nematode metabolic footprints
The effects of straw incorporation on nematode metabolic footprints are listed in Table 5 and Appendix F. In the -NPK soils,straw incorporation significantly increased the values of both PPfoot and Sfoot by 87.05 and 347.92% (P<0.01 andP<0.05),but significantly decreased the value of Efoot by 52.56% in May (P<0.05). In October,straw incorporation increased both PPfoot and Sfoot by 133.40 and 181.83%,respectively (P<0.05 andP<0.10). Regardless of sampling time effect,straw incorporation significantly increased the values of PPfoot and Sfoot by 97.27 and 305.39% (P<0.01 andP<0.01),but significantly decreased the value of Efoot by 39.53% (P<0.05).

Table 4 Nematode ecological indices with straw removal (-S) and straw incorporation (+S) in non-fertilized (-NPK) and fertilized (+NPK) soils
In the +NPK soils,no significant effects of straw incorporation were observed at the sampling of May,but straw incorporation in October significantly increased FFfoot and Sfoot by 124.13 and 145.92% (P<0.05 andP<0.05),respectively. Regardless of sampling time effect,straw incorporation significantly increased the values of FFfoot and Sfoot by 16.19 and 61.17% (P<0.01 andP<0.01).The degree of Sfoot improvement was higher in the -NPK soils than in the +NPK soils. Regardless of sampling time and fertilization effects,straw incorporation significantly increased the values of PPfoot and Sfoot by 11.29 and 149.56% (P<0.10 andP<0.001).
Of the three channel metabolic footprints (BFfoot,FFfoot and PPfoot),PPfoot accounted for 44.51%,followed by BFfoot (38.82%). The functional metabolic footprint of nematodes is illustrated in Fig.1. Under the -NPK and +NPK conditions,the FMF was greater in the +S treatment than in the -S treatment at both sampling seasons. In addition,the increasing rates were much greater in the -NPK soils (92.91%) than in the +NPK soils (15.20%).
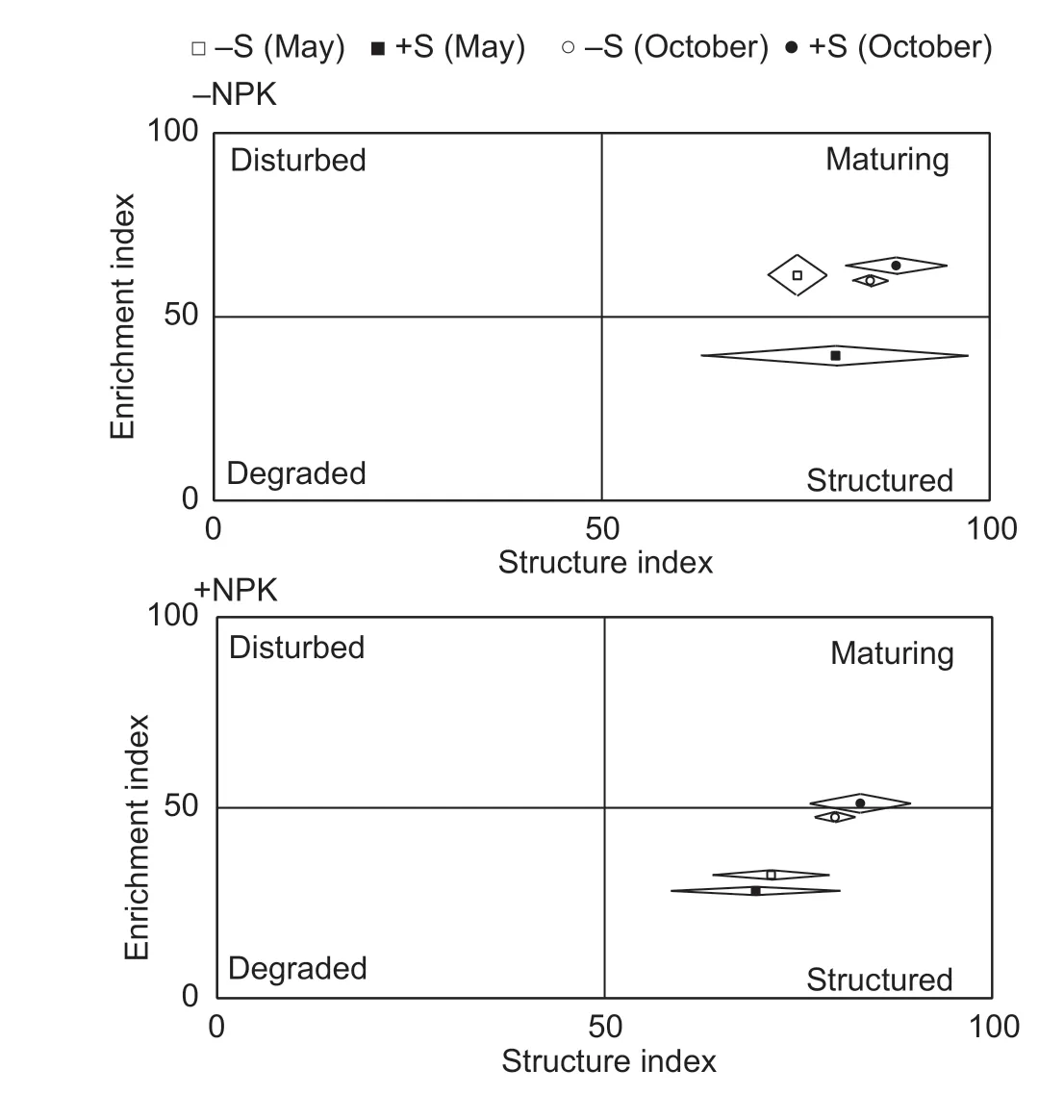
Fig.1 Functional metabolic footprints of nematode commnuities subject to straw removal (-S) and straw return (+S) in (A) non-fertilized (-NPK) and (B) fertilized (+NPK) soils at the sampling of May and October. The horizontal axis of each footprint represents the structure footprint and is calculated as SI-0.5Fs/k and SI+0.5Fs/k,and the vertical axis of each footprint represents the enrichment footprint and is calcaulated as EI+0.5Fe/k and EI-0.5Fe/k. The functional metabolic footprint is described by the sequentially joining points:(SI-0.5Fs/k,EI),(SI,EI+0.5Fe/k),(SI+0.5Fs/k,EI),(SI,EI-0.5Fe/k),and (SI,EI) (central point). The k value is 7.1 in May and 4.0 in October. SI,structure index;EI,enrichment index.
3.5.Redundancy analysis of nematode genera in response to soil physiochemical properties
Redundancy analysis (RA) revealed that NH4+-N was the most important factor influencing the distribution of nematode genera,followed by pH (Fig.2). The eigenvalues were 0.239 (F=6.88,P=0.002) and 0.158 (F=5.97,P=0.002) for the first axis and all canonical axes,respectively,and the first two axes could account for 73.8% of the variation.
4.Discussion
4.1.Soil physicochemical properties and crop yield
After decomposition,the incorporated straws return large amounts of C and K,and portions of P and N into the soils,leading to an increase in soil nutrients and crop yields (Liuet al.2014). This study revealed that the contents of SOC,AN,AK and crop yields were significantly increased by straw incorporation under the fertilized condition (Table 1).These results are consistent with those reported in previous long-term studies (Baiet al.2015;Zhaoet al.2016;Wanget al.2018;Puet al.2019). In addition,the degrees of improvement in soil nutrients and crop yields were higher at low fertility than at high fertility (Appendix A),probably due to the law of diminishing marginal returns. In agreement with these results,Zhang Y Let al.(2012) compared the effects of straw return on soil physicochemical properties and wheat yield under NP fertilizer and NPK conditions,and found that the degrees of improvement of most selected properties were higher under NP conditions than under NPK conditions.
4.2.Nematode abundances and community compositions
The C and energy enter the soil food web and,thereby,are delivered along the trophic levels and affect the abundances of total nematodes and trophic groups present at different trophic levels. In low fertility soils,straw incorporation had significantly positive effects on the abundances of total nematodes and trophic groups,except for fungivores (Table 2;Appendix C),consistent with the findings made in most previous short-term studies (Liet al.2009,2018;Zhang X Ket al.2012;Mouet al.2017;Chen Y Fet al.2018).Generally,micro-feeding nematodes (bacterivores and fungivores) and omnivores-predators have been regarded as the beneficial nematodes because they can stimulate soil nitrogen mineralization and suppress the pests (Ferriset al.2004). Therefore,in this study,straw incorporation contributed to the improvement of soil function. However,plant-parasites,as harmful nematodes,are generally hidden in crop straws and can enter into soils during straw return,resulting in a potential risk to crop growth. Our results implied that straw incorporation also had negative effects on crops. In high fertility soils,the differences in nematode abundance between straw removal and straw incorporation were similar to those of low fertility soils,but the degrees of improvement were lower,probably due to the reason that the resource supply rate is higher in low fertility soils than in high fertility soils. Similarly,Zhang X Ket al.(2012) reported that the degrees of improvement of total nematodes,plant-parasites and omnivores-predators resulting from wheat straw incorporation were higher in a convention tillage system (low fertility) than in a non-tillage system (high fertility). Zhaoet al.(2019) also observed similar trends on bacterial and fungal biomass under an incubation experiment. Notably,the abundances of total nematodes and trophic groups were higher in fertilized soils than in non-fertilized soils,revealing that the application rate of mineral fertilizer is appropriate in this study and does not cause any negative influences on nematodes.

Of the four trophic groups,plant-parasites was the most abundant group. This is in agreement with the findings in several previous reports (Zhang X Ket al.2012;Mouet al.2017;Chen Y Fet al.2018;Liet al.2018). However,a number of other studies showed that bacterivores was the most abundant group (Liet al.2009;Yeet al.2013;Liuet al.2015). These inconsistent results implied that straw incorporation did not affect the intrinsic characteristics of the soil nematode community. An RDA analysis showed that NH4+-N was the relatively important factor contributing to the distribution profile of nematode genera (Fig.2),probably due to the large input of straw-C,which needs N input to balance the C/N ratio for the growth of soil microorganisms. Zhaoet al.(2019) also found NH4+-N mainly influenced the distribution of bacterial community composition in the straw amendment treatments. Notably,the difference in concentration of NH4+-N between -NPK and +NPK soils under straw incorporation was not significant (Table 1;Appendix B),but there was a significant difference in the nematode communities between them. This result implied that NH4+-N probably was not the leading factor affecting nematode communities. A possible reason is related to the sampling time. Normally,the chemical fertilizer,especially N fertilizer,is dissolved rapidly in the soil water and the effects of fertilizer on soil and plant would last 7-30 days. However,the sampling time in this study was after wheat or rice harvest,which could result in the NH4+-N difference between -NPK and +NPK soils not being observed. Wanget al.(2020) found that the NH4+-N difference between no fertilizer and N fertilizer treatments was significant in the early growth stage of maize but not significant in the late stage.
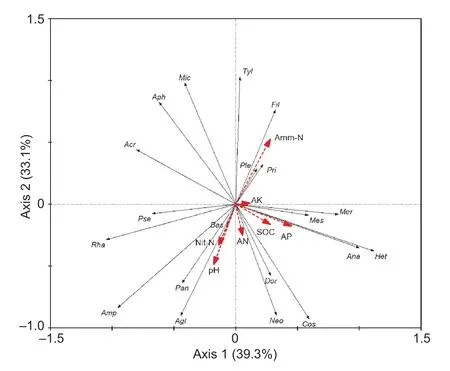
Fig.2 Redundancy analysis (RDA) of the relationship between nematode genera and soil physicochemical properties. SOC,soil organic carbon;AN,available nitrogen;Amm-N,ammonium N;Nit-N,nitrate nitrogen;AP,available phosphorus;AK,available potassium;Acr,Acrobeloides;Ana,Anaplectus;Het,Heterocephalobus;Mic,Microlaimus;Pan,Panagrellus;Ple,Plectus;Pri,Prismatolaimus;Pse,Pseudoaulolaimus;Rha,Rhabdolaimus;Aph,Aphelenchoides;Dor,Dorylaimoides;Fil,Filenchus;Agl,Aglenchus;Amp,Amplimerlinius;Bas,Basiria;Cos,Coslenchus;Mer,Merlinius;Neo,Neopsilenchus;Ott,Ottolenchus;Tyl,Tylenchus;Mes,Mesodorylaimus.
4.3.Ecological indices of nematode
The values of diversity index (H´) suggest that straw incorporation increased the diversity of the nematode community regardless of sampling and fertilization effects (Table 4),consistent with those reported in previous studies (Liet al.2009,2018;Zhang X Ket al.2012;Mouet al.2017).This may be because the incorporated straws change the relative abundances of different SOC factions (Zhaoet al.2016;Chen Z Met al.2018),which,in turn,change the abundance and distribution at the genus level.
MI is a weighted mean of cp value for all individual nonplant-feeding nematodes,and a high MI value indicates a stable environment (Bongers 1990;Bongiornoet al.2019). In the present study,we observed that long-term straw incorporation increased the MI value regardless of sampling and fertilization effects,indicating that straw incorporation drives the soil food web toward a relatively stable environment. This result is in accordance with previous studies (Liet al.2009;Zhang X Ket al.2012;Mouet al.2017;Liet al.2018).
The EI measures the resource availability and the responses of primary decomposers to resources. SI measures the degree of trophic links,or soil food web complexity. The combination of both EI and SI can be used to infer soil conditions (Ferriset al.2001;Bongiornoet al.2019). In the present study,we observed that straw incorporation did not impose positive influences on EI but tended to increase SI (Table 4),suggesting that the straw incorporation does not increase the bottom-up constraint but tends to increase the soil food web connection and complexity. These results are different from those reported by some short-term studies,which showed positive effects of straw incorporation on EI and SI (Liet al.2009;Zhang X Ket al.2012). In the EI-SI graph,all the treatments were located in the maturing or structured quadrants (Fig.1),suggesting that straw incorporation imposes a low or moderate disturbance on the soil food web,which may be the reason explaining why straw incorporation does not significantly affect EI and SI. Additionally,the treatments in high fertility soils were preferentially located in the structured quadrant,suggesting that the disturbances are lower at high fertility soils than at low fertility soils. Similar trends were also observed by Zhang X Ket al.(2012).
4.4.Metabolic footprints of nematodes
Metabolic footprints of bacterivores,fungivores and plantparasites indicate the amounts of C and energy entering the soil food web through their respective channels (Bhusalet al.2015). Of the three energy channels,plant-parasite footprints were the highest at the two sampling times (Table 5;Appendix F),indicating a greater flow rate of resources into the food web through the plant channel.Additionally,straw incorporation significantly increased plant-parasite footprints,particularly in non-fertilized soils,suggesting that straw incorporation increases the assimilation of plant-derived C. In additional to its effects on plant-parasite footprints,straw incorporation also positively influenced the fungivore footprint with significant increases being observed in non-fertilized soils (Table 5),suggesting that long-term incorporated straw-C,especially at low fertility conditions,increases the size of the fungal channel. This may be because the high C/N ratio of straw tends to shift the size of the fungal energy channel dominated by fungi and fungal feeders,such as fungivore nematodes (Holtkampet al.2008;Zhanget al.2016;Guanet al.2018).
The enrichment footprint represents the ecosystem service of nutrient mineralization (Ferriset al.2012). In the present study,we observed that straw incorporation did not cause obvious effects on the enrichment footprint in either non-fertilized or fertilized soils,indicating that straw incorporation does not increase the magnitude of nutrient mineralization. Generally,the long-term (12-year) straw incorporation results in saturation of soil C (Liuet al.2014).Therefore,the mineralization rates of young straw-C and the incorporated straw-C of every crop season are probably slow. Zhang X Ket al.(2012) demonstrated that short-term straw incorporation increased the enrichment footprint. The structural footprint represents top-down regulatory function in the soil food web (Ferris 2010). In this study,we observed that straw incorporation significantly enhanced the structural footprint in both low and high fertility soils,suggesting that straw incorporation increases the top-down pressure on food web structure. A greater functional footprint indicates a higher C used in production and a higher total rate of CO2evolution for the nematodes (Zhonget al.2017). Straw incorporation increased the functional footprint and greater improvements were found in non-fertilized soils (Fig.1),indicating that straw incorporation improves the C utilization by nematodes,especially under low fertility conditions.
4.5.How did long-term straw incorporation regulate the soil food web?
Generally,the size and activity of the soil food web are regulated simultaneously by resources,predation and/or trophic cascade effects,i.e.,bottom-up (donor) and topdown controllers (Ferriset al.2001;Lenoiret al.2007).A number of nematode ecological indices can be used to indicate the top-down effect,such as the absolute and relative abundances of omnivores-predators,MI,SI and structural footprint (Ferriset al.2001,2012;Chenet al.2019). In this study,we observed that all these indices were increased significantly by straw incorporation (Tables 2-5;Appendices C-F),particularly in non-fertilized soils,suggesting that the top-down effect resulting from straw incorporation is strong. This may be due to the fact that long-term straw incorporation increases the proportions of large and small macro-aggregates (Zhaoet al.2018),and thereby provides bigger pores favorable to the growth of nematodes,especially of omnivores-predators with larger body diameters. On the other hand,several indices could be used to indicate a bottom-up effect,such as absolute and relative abundances of bacterivores and fungivores,metabolic footprints of bacterivores and fungivores,or the enrichment index (Ferriset al.2001,2012;Chenet al.2019).However,these indices,except bacterivore abundance,were not sensitive to straw incorporation (Tables 2-5;Appendices C-F),indicating that the bottom-up effects of straw incorporation are inconspicuous. Numerous studies demonstrated that short-term and long-term straw return increased the abundance/biomass of the microorganisms (Zhaoet al.2016,2017,2019;Chenet al.2017;Chen Y Fet al.2018;Zhao and Zhang 2018;Baiet al.2019),and showed a strong bottom-up effect on the soil food web. However,this effect cannot be extended to nematodes,which is probably due to the face that the strong top-down effect suppresses the opportunist nematodes that respond to resources quickly. In this study,the top-down effect was mainly exerted by omnivores-predators. However,some predators that were not included in the present study,such as mites and collembola,could also impose the top-down effect. Other scientists also found similar results. For example,Holtkampet al.(2008) found that the biomasses of soil microorganisms at the entry level of the soil food web were increased during ecosystem development after land abandonment,but did not increase at higher trophic levels. Zhanget al.(2016) also found that wheat straw significantly increased the biomasses of bacteria and fungi in the maize growth season in a 23-year fertilization experiment,but not the biomass of micro-feeding nematodes.
5.Conclusion
We observed that long-term straw incorporation increased the abundances of total nematodes and trophic groups of bacterivores,plant-parasites and omnivores-predators under both non-fertilizer and fertilizer conditions,and that greater improvements were observed in non-fertilized soils. Nematode community analysis showed that straw incorporation increased the nematode diversity and community stability regardless of sampling and fertilization effects. Nematode footprints revealed that straw incorporation increased the resource assimilation of the plant channel and the top-down pressure on the soil food web especially in non-fertilized soils. Among the driving forces of nematode communities,a top-down effect mediated by straw incorporation was relatively more important.
Acknowledgements
This study was funded by the Open Project of Key Laboratory of the Ministry of Agriculture and Rural Affairs,China (KLFAW201705),the National Natural Science Foundation of China (31870501),the Technological Achievements Cultivation Project of Hubei Academy of Agricultural Sciences (2017CGPY01) and the Key Project of Technological Innovation in Hubei Province,China (2018ABA091).
Declaration of competing interest
The authors declare that they have no conflict of interest.
Appendicesassociated with this paper are available on http://www.ChinaAgriSci.com/V2/En/appendix.htm
杂志排行

Journal of Integrative Agriculture的其它文章
- Adoption of small-scale irrigation technologies and its impact on land productivity:Evidence from Rwanda
- African swine fever and meat prices fluctuation:An empirical study in China based on TVP-VAR model
- The impacts of oxytetracycline on humification during manure composting can be alleviated by adjusting initial moisture contents as illustrated by NMR
- Functional diversity of soil microbial communities in response to supplementing 50% of the mineral N fertilizer with organic fertilizer in an oat field
- Nitrogen acquisition,fixation and transfer in maize/alfalfa intercrops are increased through root contact and morphological responses to interspecies competition
- Kaempferol inhibits Pseudorabies virus replication in vitro through regulation of MAPKs and NF-κB signaling pathways