不同地理类型青杨派古树扦插苗对干旱胁迫的响应*
2021-06-24潘立陆燕元
潘立,陆燕元
(1.西南林业大学 会计学院,云南 昆明 650224;2.国家林业局西南地区生物多样性保育重点实验室,西南林业大学,云南 昆明 650224)
川西高原位于青藏高原东南部,地形地貌复杂,气候环境多样,冰川活动,地理条件独特,是青杨派杨树适宜的生长地,也是我国及全球的杨树(Populus)分布中心之一[1],蕴藏着丰富的古杨树资源[2]。这些珍稀古树是在多个生态环境因子和人为因子的长期综合作用下生存下来的,适应所处地区的气候、土壤等环境条件,抗逆能力较强,是最地道的乡土树种。对它们进行深入研究和利用,有利于广大高原地区的造林绿化,在防止耕地和草地退化,提供牧畜饲料,涵养水源,改善生态环境和发展少数民族地区经济等方面都具有十分重要的意义,同时,也可为杨树育种提供新的种质资源。
干旱是世界上危害最为严重的自然灾害之一,其出现次数、持续时间、影响范围、造成的损失等均居各种自然灾害之首[3]。近40 a来,随着大气中温室气体含量的升高,我国气候总体出现变暖变干的趋势[4-5]。这种趋势对我国农业、林业生产以及整个生态环境造成了极大的不利影响[6]。西南地区属于干旱半干旱气候区,干湿季分明,时常遭受季节性干旱的影响,而且近年来干旱频繁发生,持续的时间及程度均较严重。在生态环境十分脆弱的情况下生存下来的古杨树历经百年乃至千年的自然环境考验,对所处地区的气候与土壤等环境条件的抗逆境能力较强,利用这些古树资源来研究其应对干旱的生理生化机制,从而为西南地区选择抗旱性较强的古杨树种提供理论依据和参考。
在本区域不同的地理环境中,不同种类的杨树对气候环境表现出明显的差异,据此划分出不同的地理类型[7]。三脉青杨(Populustrinervis)、乡城杨(P.xiangchengensis)和康定杨(P.kangdingensis)分别是干热河谷型、温湿型及寒湿型等3种地理类型的典型代表,是全国乃至世界少有的分布于低纬度高海拔地区的宝贵杨树资源,在高山造林、高原地区绿化树种选择中有极大的优势[8-9]。贾晨等在对川西高原的几种乡土杨树树种研究中发现,三脉青杨、乡城杨和康定杨在年高生长、胸径生长及材积生长过程中均有较大优势,造林效果显著[10-11]。然而,其对环境适应的内在机理仍缺乏研究,因此,本试验以胸径≥1 m且树龄在300~500 a的这3种古杨树扦插幼苗为研究对象,研究它们在干旱胁迫下生理生化及分子差异响应,以揭示其对干旱环境胁迫的适应机理,为该地区生态环境建设中抗旱性较强树种的筛选提供依据,促进乡土古杨树资源的利用及开发。
1 试验材料与方法
1.1 试验材料与培养
本试验于2016年对云南、四川、西藏等西南地区的古杨树资源调查中选取胸径≥1m,树龄在300~500 a生的三脉青杨、乡城杨和康定杨3种青杨派古杨树为对象,分别剪取阳面上部枝条(直径约2 cm,长度约150 cm)20个,带回西南林业大学全自动智能温室的试验地中,剪成长30 cm的枝段,消毒、抹芽处理后进行扦插。2018年5月剪取2 a生的健壮枝条进行扦插,在成活后按照日常进行水肥管理。于2019年6月,每种选择10个健壮、无病虫害、粗度、高度及生长势大体一致的枝条,剪枝每段约30 cm,在含有1 000 mg/L生根粉和稀释2 000倍多菌灵的水中泡足14 h后,扦插到装满匀质红壤珍珠岩的塑料盆中进行培育。培育期间在温室环境中生长,待苗木成活后计算扦插成活率,生长健壮、稳定后开始试验处理。
1.2 自然干旱胁迫试验
干旱胁迫试验在温室中进行。温室条件为光照时间18 h,湿度为30%~55%,温度为16~25 ℃。干旱胁迫采用自然干旱法:即当植株长至60~80 cm时,试验前一次性浇足水,之后不再浇水,直至植株干枯死亡,隔天进行相关指标的测定,整个试验过程持续12~14 d。
1.3 相关指标测定方法
1.3.1 土壤含水量的测定
开始自然干旱胁迫后,以第0 d作为对照,采用TDR 200土壤水分测量仪(美国Spectrum)结合称重的方法测定土壤中的相对含水量,每个树种每个处理测定3盆,每盆测定3次,取平均值作为数据分析。
1.3.2 叶片相对含水量(RWC)的测定
按照高俊风[12]的方法测定,从干旱第0 d开始剪取枝条顶端下数第3~4片生长良好的成熟叶片,迅速放入锡箔纸中,用分析天平称出鲜重(Wf),将样品完全浸入蒸馏水中24 h,取出用吸水纸擦干样品表面水分,称出饱和重(Wt)。然后再放入烘箱,于105 ℃下放置0.5 h杀死叶片,后于80 ℃下烘干至恒重,称出干重Wd,根据下列公式计算出叶片相对含水量:
RWC=(Wf-Wd)/(Wt-Wd)×100%
1.3.3 光合系统参数的测定
采用便携式光合仪测定系统〔CIRAS-3光合仪(美国)PP SYSTEMS〕测定光合系统参数。于上午9:00—11:00进行测定,选取从枝条顶端下数第3~4片生长良好的成熟叶,测定净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、细胞间CO2浓度(Ci)、水分利用效率(EWU)等参数。为了保证测量在近似于理想光合作用状态下进行,测定时将叶面温度控制为25 ℃,光照强度控制为1 400 μmol/(m2·s),相对湿度控制为50%左右,CO2浓度控制为(350 ± 5)μmol/mol。每盆测定3次,隔天进行测定,直到叶片干枯旱死、净光合速率为负值时停止测定。
1.3.4 抗氧化酶系统的测定
每隔2 d,10:30进行一次统一采样,分别剪取处理苗木第3、4、5完全展开叶的混合样,经液氮速冻后置于-80 ℃冰箱中,用于超氧化物歧化酶活性、过氧化物酶活性、抗坏血酸过氧化物酶活性、丙二醛含量以及相关抗旱基因表达量的测定。
超氧化物歧化酶(SOD)用氮蓝四唑(NBT)光还原法[13-15]测定。抗坏血酸过氧化物酶(APX)测定参照Manon[16]、 沈文飚等[17]的方法,利用APX 在H2O2存在的条件下使抗坏血酸量减少的原理测定酶活性。过氧化物酶(POD)根据Chance和Maehly[18]的方法,利用过氧化物酶能促使愈创木酚氧化生成棕红色4-邻甲氨基苯酚,在470 nm 处有特征吸收峰,在一定范围内其颜色深浅与4-邻甲氨基苯酚的量成比例关系,从而间接测定过氧化物酶的活性。
1.3.5 活性氧自由基指标测定及膜脂过氧化物质含量测定
膜脂过氧化物(MDA)的测定方法参照Hodges[19]、孙群[20]和赵世杰等[21]等所描述的硫代巴比妥酸法。过氧化氢(H2O2)含量测定参照Mukherjee和 Choudhuri的方法[22]。H2O2可与氯化钛(TiCl4)反应生成过氧化物-钛复合物黄色沉淀,待黄色沉淀被H2SO4溶解后,溶液呈黄色,在415 nm处有最大吸收。超氧阴离子产生速率(O2-.)含量测定测定参照王爱国等[21]的方法。细胞膜透性用 DDS-11A 电导率仪测定电导率,用相对电导率(%)表示。
1.3.6 抗逆基因SOD,lea2、lea3组基因的瞬时表达量测定
采样 每隔2 d ,10:30进行采样一次,分别剪取处理苗木顶端往下第3、4、5片完全展开叶的混合叶样,经液氮速冻后置于-80 ℃冰箱中,采用Real-time PCR法检测cDNA样本中SOD、lea2、lea3基因相对含量〔生工生物工程(上海)股份有限公司〕。
RNA提取 SK8661 柱式植物总RNA抽提纯化试剂盒;
RNA样品的浓度和质量监测 电泳检测-RNA电泳结果(1.5%琼脂糖,1×TAE电泳缓冲液),观察并拍照。
反转录 第一链cDNA合成试剂盒(RevertAid Premium Reverse Transcriptase,Thermo ScientificTMEP0733), RNA按照800 ng反转。
引物序列(Primer Premier 5.0软件设计) 选取在植物中比较保守的基因β-actin作为内参基因,SOD,lea2、lea3组基因做为目的基因(表1)。

表1 引物及其序列Tab.1 Primers and their seqences
荧光定量PCR检测 将cDNA样品稀释10倍作为模板上机检测,定量PCR试剂采用SG Fast qPCR Master Mix(2X)(BBI)(B639271),定量PCR仪用LightCycler 480 (美国Roche)。
1.4 数据分析
统计所测数据,使用Excel 2010软件进行计算,Sigmaplot 10.0软件作图,由SPSS软件(20.0版本)进行数据统计分析,采用Duncan法进行多重比较。
2 结果与分析
2.1 自然干旱胁迫对土壤含水量的影响
土壤含水量是影响植物生长发育的关键因素。土壤水分亏缺时,植物的许多功能都会受到限制。
由图1可以看出,自然干旱胁迫对不同种古杨树的土壤含水量都有明显影响。随着自然干旱胁迫处理时间的增加,土壤含水量总体呈下降趋势,在干旱胁迫2~10 d,土壤含水量急剧下降,到第10 d以后土壤含水量处于一个较低水平,康定杨日均土壤含水量变化最快,降至16.4%,接下来土壤含水量变化变慢,但最后均已降至10%以下。
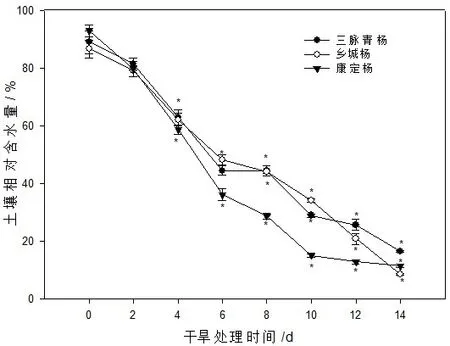
图1 自然干旱胁迫对土壤含水量的影响注:*表示各自的处理组与对照组间存在显著差异 (P<0.05),下同。Fig.1 Effects of natural drought stress on soil water content
2.2 自然干旱胁迫对古杨树扦插苗叶片生长的影响
土壤干旱持续时间对古杨树幼苗形态产生显著影响。
以三脉青杨为例(图2),停止浇水初期(0~6 d),植株总体形态变化并不明显;处理10 d后,伴随土壤含水量的显著下降,土壤表面看起来已经很干,轻微结块,此时植株形态开始明显变化,首先是叶片出现轻微下垂,随后茎尖萎蔫,叶片完全下垂,最后叶片干枯,整个植株呈枯死状态,此时表层土壤已完全变干,用硬物已经插不进去,但叶片没有掉落;在后期的试验发现,植株叶片干枯1个月后,幼苗一旦重新浇水,干枯的叶片会很快掉落,然后又长出新叶,说明古杨树幼苗对干旱逆境有很好的适应性,其恢复能力也很强。遭受干旱胁迫后,乡城杨和康定杨形态上的变化趋势与三脉青杨的一致,只是在关键时间节点上有轻微差异,如康定杨出现萎蔫的时间会提前1 d(图3),而乡城杨则往后延长1 d(图4),存活的时间较长。

图2 自然干旱胁迫对三脉青杨植株形态的影响Fig.2 Effects of natural drought stress on the plant morphology of P.trinervis

图3 自然干旱胁迫对康定杨植株形态的影响Fig.3 Effects of natural drought stress on the plant morphology of P.kangdingensis

图4 自然干旱胁迫对乡城杨植株形态的影响Fig.4 Effects of natural drought stress on the plant morphology of P.xiangchengensis
2.3 自然干旱胁迫对古杨树叶片相对含水量(RWC)的影响
叶片相对含水量的高低能够反映植物的生长代谢情况。在干旱条件下,叶片相对含水量高的植物拥有更旺盛的生命功能,对干旱适应能力更强[6]。从图5可知,随着自然干旱胁迫的持续,3种古杨树的叶片含水量均呈现出持续下降的趋势。自然干旱胁迫初期(0~6 d),3种杨树的叶片相对含水量的下降速度均较慢,与对照相比均无显著差异。之后,随干旱胁迫时间的延长,3种杨树的叶片相对含水量均显著下降,其中三脉青杨的降幅最为显著,至干旱后期时其叶片RWC仅为对照的63%,而乡城杨和康定杨分别为对照的77%和78%,表明随着自然干旱胁迫时间的持续,乡城杨和康定杨叶片保水能力较强,对干旱的适应能力较强,而三脉青杨叶片保水能力较弱。
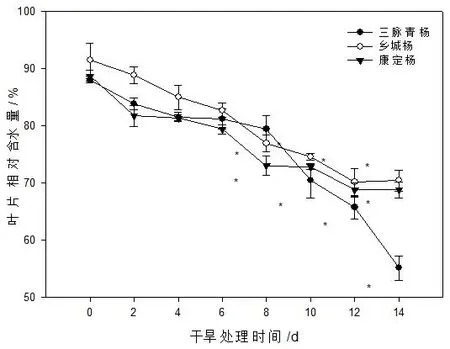
图5 土壤自然干旱对古杨树扦插苗叶片RWC的影响Fig.5 Effect of soil natural drought on RWC of ancient poplar cutting seedling leaves
2.4 自然干旱胁迫对古杨树光合特性的影响
如图6所示,干旱胁迫对古杨树扦插苗叶片光合系统产生重要影响。叶片净光合速率Pn随干旱胁迫时间的持续呈下降趋势,但在干旱中前期(1~8 d)时降幅较为缓慢。在水分充足条件下,相对于乡城杨和康定杨,虽然三脉青杨叶片Pn较低,但受到干旱胁迫后却比较稳定,降幅较为缓慢,例如干旱第9 d,与对照相比,其降幅仅为17%,而乡城杨和康定杨21%分别为47%。干旱第10 d后, 3种杨树的Pn均显著下降,第12 d时,三脉青杨的Pn将为负值,呼吸作用大于光合作用,而康定杨和乡城杨的光合作用则可以分别维持至12和15d(图6A)。气孔导度(Gs)的变化趋势与Pn略微相似,随干旱胁迫持续基本呈逐渐下降的趋势,但康定杨在干旱初期(1~2 d)显著下降后在中期略有上升,在干旱第9 d是才显著下降,而乡城杨和康定杨一直呈平缓下降的趋势(图6B)。胞间二氧化碳的浓度(Ci)受气孔导度和体内光合活性的综合影响。在干旱中前期(1~11 d),古杨树扦插幼苗叶片Ci的变化不大,略有升降的起伏,3种杨树之间的差异也不明显;干旱后期(12 d),三脉青杨和康定杨的Ci显著上升,分别为对照的190%和149%,与对照差异显著,而乡城杨直至干旱胁迫的第15 d其Ci值升至对照的117%(图6C)。叶片蒸腾速率(Tr)与气孔导度密切相关,因为气孔的开合程度影响植物体内水分的进出及气体交换。由图6 D可以看出,干旱胁迫后,Tr的变化趋势与Gs基本相似,都是先降后升再降。一般来说,一定程度的干旱可以促进植物水分利用效率(EWU)的提高。在本试验中,干旱胁迫处理第1 d时,3种古杨树的叶片的EWU轻微下降然后上升,乡城杨和康定杨的EWU在干旱胁迫第2 d达到最高值,但与对照相比均没达到显著性差异,而三脉青杨受干旱胁迫后叶片EWU逐渐上升,至第5~6 d时达到最大值,随后开始显著下降。总体上来说,乡城杨的EWU至干旱后期变化较为平缓,干旱胁迫第15 d时降幅为对照的63%,而三脉青杨分别为193%和134%,与对照的差异极为显著(图6E)。
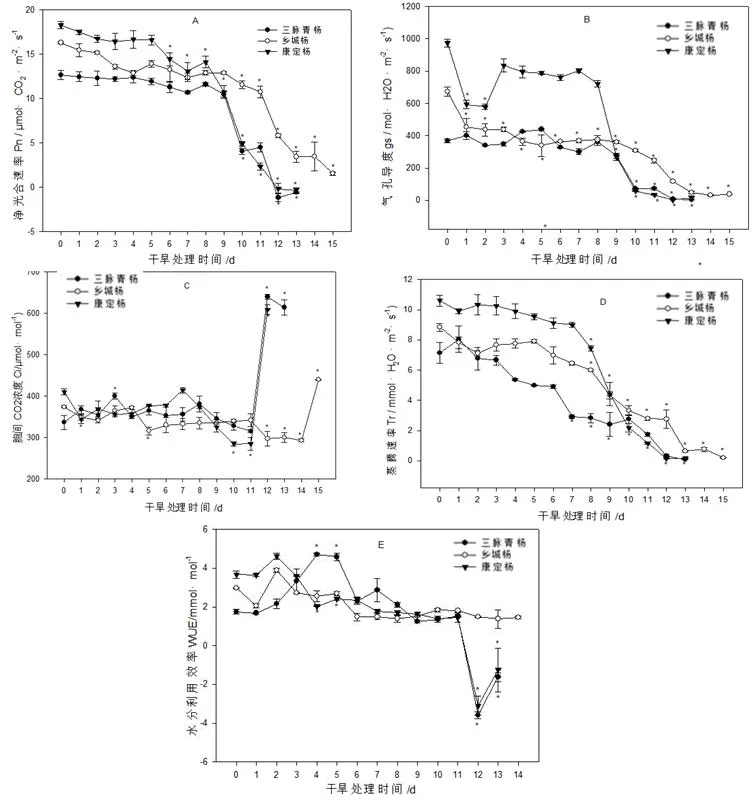
图6 土壤自然干旱对对古杨树扦插苗叶片光合特性的影响Fig.6 Effects of natural drought on photosynthetic characteristics of ancient poplar cuttings
2.5 自然干旱胁迫对古杨树抗氧化酶系统的影响
土壤自然干旱处理对古杨树3个种的叶片超氧化物歧化酶SOD活性变化产生不同影响(图7)。随干旱胁迫时间的持续,三脉青杨叶片SOD呈现升—降—升—降的双峰值趋势,在干旱的第2 d和4 d分别出现次大和最大峰值,涨幅分别为37%和32%,与对照差异显著,干旱后期略有上升,随后降至对照水平。乡城杨叶片SOD受到干旱胁迫后逐渐上升,第10 d时达到最大值,涨幅为181%,后期略有下降,但与对照相比,涨幅仍然较大,为147%,与对照达到极显著差异水平(P<0.001)。受到干旱胁迫后,康定杨叶片SOD总体上变化不明显,至干旱第14 d时升至最大值,涨幅为23%,与对照差异不明显。抗坏血酸过氧化物酶(APX)在土壤自然干旱过程中的变化趋势与SOD不同,在3个树种中的表现也不同。土壤自然干旱处理下,三脉青杨叶片APX先显著升后降再上升,至干旱后期(第14 d)时达到最大值,涨幅为430%,与对照相比差异极为显著。而乡城杨和康定杨在干旱胁迫的影响下,APX活性均逐渐下降,乡城杨在干旱第14 d时略有回升,但没有恢复到对照水平,在整个干旱过程中乡城杨和康定杨的APX活性降幅与对照相比没有达到显著性差异,两个种之间差异也不显著。过氧化物酶POD受干旱的影响与SOD相似,呈升—降—升—降的双峰变化趋势,但变化幅度较小。其中,康定杨POD的变化趋势最为显著,在干旱胁迫第4d和第10d,其值达到最高值,涨幅为110%,与对照差异显著。三脉青杨POD酶活性最小,在干旱第10 d达到最大值是涨幅仅为对照的35%,而乡城杨叶片POD活性在干旱的第8 d和10 d涨幅分别为对照的54%和47%左右,差异显著。

图7 土壤自然干旱对古杨树扦插苗叶片抗氧化酶系统的影响Fig.7 Effect of soil natural drought on antioxidant enzyme system of ancient poplar cutting
2.6 自然干旱胁迫对古杨树叶片活性氧含量的影响
丙二醛MDA是膜脂过氧化的产物,含量越高表明膜脂受到的伤害越严重。如图8所示,受旱后,三脉青杨和乡城杨叶片MDA含量逐渐上升,到干旱后期达最大值,涨幅分别为184%和70%,前者与对照达到及显著差异,后者差异显著;康定杨则先升后降再升,干旱后期涨幅为277%,差异极为显著。这些结果表明受干旱胁迫后,3个古杨树扦插苗的膜脂均受到较大的破坏,其中康定杨受损最为严重,而乡城杨相对较轻。细胞膜是活细胞与环境之间的界面和屏障,各种不良环境对细胞的影响往往首先作用于细胞膜,改变其透性, 而相对电导率是衡量细胞膜透性的重要指标,其值越大,表示电解质的渗漏量越多,细胞膜受害程度越重。如图8所示,干旱胁迫持续时间对不同种古杨树的细胞膜相对透性产生很大影响。土壤自然干旱处理后的1~8 d,3个树种叶片细胞膜相对透性均呈先上升后下降的趋势,但变化幅度不大,3种杨树之间也没有明显的差异;干旱第10 d,该值开始呈显著上升的趋势,干旱第14 d均达到最大值,与对照相比,三脉青杨、乡城杨和康定杨分别上升了264%,224%和184%,差异极为显著。表明在一定的干旱范围内,古杨树通过自身的调节,可以减轻细胞膜受干旱的伤害,一旦超过一定的范围,植物各个功能失调,细胞膜相对透性增大,电解质流失严重。

图8 土壤自然干旱对古杨树扦插苗叶片细胞膜的影响Fig.8 Effect of soil natural drought on cell membrane system of ancient poplar cutting
过氧化氢(H2O2)是活性氧的一种,多种外界刺激或细胞内信号分子都可以导致细胞内H2O2的积累。如图9所示,自然干旱处理下,三脉青杨和康定杨过氧化氢(H2O2)逐渐升高,中期略有下降,后期又上升;而乡城杨在干旱前期0~6 d无明显变化,8 d时明显上升;三脉青杨、康定杨和乡城杨在干旱第14 d时均达到最大值,上升幅度分别问其对照的 192%,201%和202%。由此可见,不同种的古杨树叶片过氧化氢含量对干旱处理的反应在时间和积累量上存在差异。
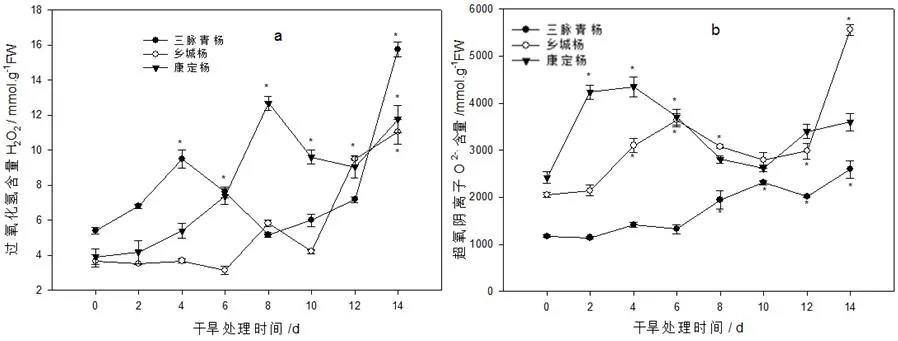
图9 土壤自然干旱对古杨树扦插苗叶片活性氧含量的影响Fig.9 Active oxygen content of natural soil drought to the ancient poplar cutting seedlings
超氧阴离子具有独特的生理功能,当其处于非平衡浓度时,则会引起组织损伤。由图9可以看出,在自然干旱胁迫下,三脉青杨、康定杨和乡城杨3个树种的超氧阴离子O2-.变化趋势基本一致,先升后降再升,干旱后期上升幅度分别为120%,170%和49%,与对照差异显著,乡城杨后期积累的O2-.最多,而康定杨最低。
2.7 自然干旱胁迫对古杨树抗旱基因表达的影响
3种古杨树叶片的SOD基因瞬时表达量在干旱初期(1~6 d)时均逐渐上升,三脉青杨从第8 d开始显著下降,直至干旱后期(第14 d)康定杨才开始下降,而乡城杨一直呈上升的趋势。表明干旱胁迫激发了SOD基因的瞬时表达量,尤其在干旱的初期和中期,而乡城杨在后期SOD基因顺势表达量增量更为明显(图10)。干旱胁迫对3种古杨树叶片Lea2瞬时表达量影响显著。干旱初期,3种杨树叶片Lea2瞬时表达量随干旱胁迫时间的持续而呈显著上升的趋势,三脉青杨和乡城杨在干旱胁迫的第6 d升至最高值,而康定杨则在第8 d时升至最大值,随后均显著下降,到干旱后期基本回落至对照水平。3种古杨树扦插苗叶片Lea3瞬时表达量受到干旱胁迫后变化趋势与Lea2变化基本相似,均是随干旱胁迫的加剧呈先升后降的趋势,其中乡城杨的变化幅度有所不同,即干旱后期Lea3瞬时表达量降幅较低,与对照相比差异仍比较显著。
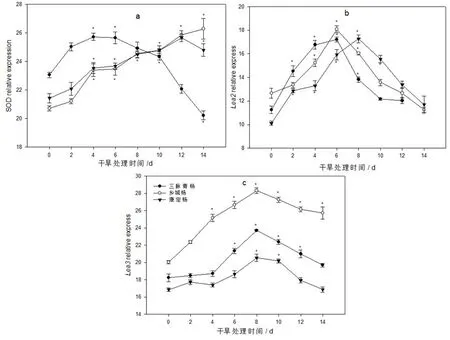
图10 土壤自然干旱对古杨树扦插苗叶片抗旱基因表达的影响Fig.10 Effect of soil natural drought on Drought resistance gene expression of ancient poplar cutting
3 讨论与结论
3.1 讨论
随着全球气候变化的加剧,水分短缺问题更加突出,植物生长、产量受到显著影响。到目前为止已有很多关于杨树对干旱胁迫的形态和生理适应的报道[25-27]。这些研究表明水分亏缺下植物可以通过改变形态结构和生长模式,调整各生理指标如光合参数特性、抗氧化酶系统、抗性基因表达等来适应干旱胁迫[28-30]。本研究发现,在干旱胁迫下,古杨树生长停滞、叶片相对含水量、净光合速率、呼吸速率、蒸腾速率和胞间CO2等气体交换指标显著降低,这与许多以前的研究相一致[31-32],但不同地理类型的古杨树在干旱胁迫下存在一系列生理和生化响应上的差异。
自然干旱胁迫后,土壤相对含水量直线下降,干旱处理14 d时3种杨树的土壤相对含水量均降至10%以下,其中寒湿型康定杨的日均土壤含水量降幅最显著。植物叶片相对含水量与土壤中有效水分含量密切相关[30]。本研究中,在自然干旱胁迫前期,3种古杨树幼苗叶片RWC变化较为平缓,而后期,三脉青杨RWC下降的幅度和速度更显著,而乡城杨则较为缓慢,表明其能更好保持体内良好的水分状况,对干旱的适应能力更强。这可能由于三脉青杨是长期处于干热河谷气候区,为适应干热的气候,气孔调节更为敏感和灵活,以避免叶温过高灼伤叶片或过度散失水分。
水分充足条件下,寒湿型康定杨光合参数均较高,而干热河谷型的三脉青杨最低。干旱胁迫后,3种地理类型的古杨树扦插苗叶片的光合作用均受到明显抑制,具体表现为净光合速率(Pn)显著下降,而叶片气孔导度(Gs)也同时下降,从而导致叶片叶绿体中CO2含量下降(由Ci值降低可以看出)。这可能是由于干旱胁迫下光合作用酶、叶绿素类及叶绿体结构等受到影响,从而抑制了光合系统的作用。此外,干旱胁迫会显著影响植物的光合电子传递[33-34]。干旱胁迫后,康定杨叶片中的Pn、Gs及Ci等下降幅度均显著高于其它两种,表明寒湿型的康定杨对干旱的适应能力较弱;而乡城杨降幅较低,且在极端干旱下(干旱处理9 d后)还能维系光合系统的稳定,表明乡城杨比其它两种杨树更适合于在极端干旱的环境下生存,因此更能解释其分布范围更广的原因,这在前人的研究上也有类似的报道[33-35]。
环境胁迫能够诱使活性氧自由基(ROS)的产生,使蛋白、膜脂发生过氧化,导致膜结构的变化,从而改变细胞膜的透性[32]。作为细胞膜破环程度的指标MDA含量和相对电导率的升高意味着膜脂过氧化的加剧[7],类似的研究也在橄榄树(OleaeuropaeaL.)中有体现[36]。尽管在干旱胁迫下3种古杨树细胞膜都遭到了破坏,但康定杨叶片中MDA和EL增幅更大,表明其受到的破坏程度更高,这一方面与细胞中积累的活性氧自由基的浓度和类型直接相关,比如受旱后,三脉青杨和康定杨H2O2积累浓度较高,而乡城杨积累的O2-.浓度较高,另一方面也与植物中抗氧化物酶的活性有关。研究表明,具有较高的抗氧化酶活性的植物,能够保护其自身免受由于环境胁迫积累的ROS的伤害,控制有害脂肪酸的积累[5]。SOD主要岐化细胞质、叶绿体和线粒体中的O2-.为O2和H2O2,是抵御ROS伤害的第一道防线。SOD在三脉青杨中活性最强,有效抑制了O2-.的积累。POD主要分布在非原质体空间和液泡中,APX分布最广泛,它在细胞溶质、叶绿体、线粒体、过氧化物酶体和非原质体空间中都被检测到[17,30]。在干旱胁迫下,三脉青杨中除POD外,SOD,APX活性均高于其它两种,说明三脉青杨清除ROS的能力较强,可有效减轻其对叶片细胞的伤害程度,类似结果在P.euphratica中也有表现[5,16,32]。
参与干旱胁迫相关的基因根据作用方式的不同可以分为两类:一类是直接参与植物抗旱能力提高的功能基因,此类基因的表达产物能维持细胞各种理化代谢活动的正常进行;另一类是起调节作用的蛋白基因,主要对信号传导和基因表达的过程进行调节,包括渗调蛋白、Lea蛋白等直接保护细胞免受水分胁迫损伤的功能蛋白基因和超氧化物酶歧化酶、抗坏血酸等毒性降解酶基因[28,34]。根据已有的研究结果[37-38]从结构上看,Lea基因在干旱胁迫下可能与抗氧化,膜系统、蛋白质的保护等功能有关,一是通过直接作用,还有一些是起分子保护屏障的作用。本研究中,通过荧光定量PCR分析,Lea2、Lea3组基因在干旱胁迫下在mRNA水平的瞬时表达量明显升高,说明了这2个基因在三脉青杨耐干旱胁迫上起了显著的作用,且两基因在干旱胁迫下,表达量的升高和降低的时间近乎同步,表明两基因在干旱胁迫下对植物应急保护机制的启动都发挥着重要的作用,这个研究结果与鲁松[37]在4个青杨(P.cathayanaRehd.)种群Lea3基因表达对干旱和低温响应的研究结果基本一致。此外,乡城杨中Lea3基因瞬时表达量增加得更显著,表明Lea3基因在乡城杨中应对干旱逆境的作用要大于Lea2基因,但长时间胁迫时两基因的表达量都明显下降,说明植物依靠其他方式应对长期干旱胁迫,如抗氧化系统酶活性刺激升高,渗透调节物质合成增加等[34]。受旱后,康定杨和乡城杨中叶片SOD基因瞬时表达量呈持续升高,而三脉青杨则先上升后下降,表明不同的抗旱基因在不同的地理类型中发挥的作用不同,这与青扦(Piceawilsonii)、海岸松(Pinuspinaster)、小麦(Triticumaestivum)等的试验结果基本类似[38-41]。
3.2 结论
综上所述,不同地理类型的古杨树对干旱的差异响应反映它们对当地气候的适应机制。受到干旱胁迫后,三脉青杨反应迅速,抗氧化酶APX和SOD活性快速升高,抗逆基因SOD瞬时表达量也显著增加,但干旱强度加剧后,三脉青杨植株受损程度最重,表明干热河谷型三脉青杨在中轻度的干旱胁迫下适应能力较好,但在严重干旱条件下适应能力较弱。而温湿型乡城杨受旱后抗逆基因Lea3和SOD瞬时表达量均同时升高,维持相对稳定的光合能力,表明其对干旱的适应的综合能力较强,因此可以解释为何乡城杨的分布范围要远远大于其它两种。在高海拔地区,一般土壤水分含量较高而且降水量充足,树木遭受的胁迫主要是霜冻和低温,所以长期生长在高海拔区域的寒湿型康定杨对低温胁迫适应能力较强,而对干旱胁迫的耐性较弱。
