基于生物量的香蕉叶片主要几何属性研究
2021-06-23刘永霞何应对连子豪王丽霞殷晓敏王必尊曹宏鑫
刘永霞,何应对,连子豪,王丽霞,殷晓敏,王必尊,曹宏鑫
(1 中国热带农业科学院海口实验站,海口,570102;2 江苏省农业科学院农业信息研究所,南京,210014)
香蕉是世界上主要的热带经济作物,主要种植区域分布在南北纬20°之间。 2018年全球香蕉种植面积565.4万hm2,产量达11 456万t;我国香蕉种植面积33.19万hm2,位居世界第六,产量1 122.17万t,其中总产量及单产均居世界第二。我国是主要的香蕉生产国,也是香蕉消费国,2016—2017年,进口额都稳定在5.8亿美元左右,2018年中国香蕉进口增长54%,达到8.96亿美元,创历史新高[1-2]。
叶片是植物主要组成器官,其空间散布状态决定了植物地上部的形态结构,进而影响植物的光截获且决定了植物最终的光合产量。20世纪80年代开始,作物功能结构模型(FSPMs)是将生长模型功能与形态模型结构有机融合产生的新一代模型[3],许多学者建立了水稻、小麦、玉米和棉花等多种作物的叶片形态及群体结构特征[4-10],然而这些模型是纯形态结构模型,不具有植物生理学机制,不能很好模拟作物生长。国际上,小麦、水稻、玉米、棉花等作物以L-PEACH[11]、GREEN-LAB[12]和LIGNUM[13]等为代表的功能—结构植物模型研究有:Cieslak等[14]应用L-studio系统软件基于生物量和冠层光合作用完成了猕猴桃藤分支结构的三维重建。Kang等[15]以向日葵生长模型SUNFLO为基础,结合GreenLab中生物量分配模块开发了向日葵功能结构模型SUNLAB。Cao等[16]基于生物量研究了水稻出苗期地上部器官的生长规律。张文宇等[17]基于油菜的生物量构建了叶长、最大叶宽和叶柄长模型。张伟欣等[18]通过分析油菜生物量与器官形态间的关系,构建了基于生物量的油菜越冬前植株地上部、分枝各器官形态结构模型。
关于香蕉功能—结构模型的研究甚少,在前期研究中我们团队建立了巴西蕉叶面积、叶长、叶宽及长宽乘积间的关系模型[19];而丰锋等[20]、安佳佳等[21]也在巴西蕉叶片形状指标与产量之间的关系上,建立回归模型。相关研究中缺少以生物量作为基础的形态结构模型,因此,我们通过分析香蕉叶片主要形态参数与生物量的定量关系,建立香蕉叶片主要形态结构参数模型,以期促进香蕉功能—结构模型的研究和应用。
1 材料与方法
1.1 材料
试验于2018年7月至2020年7月在海南省澄迈县福山镇丰西村实施,试验香蕉园面积2 hm2。供试土壤pH值5.01,有机质含量3.12%,全氮1.256 g/kg,有效磷26.69 mg/kg,速效钾141 mg/kg。试验设0、300、400、500、600 g/kg等5个N水平施肥处理,化肥施用时间及比例参照匡石滋等[22]方法。香蕉品种为巴西蕉,每处理100株,移栽前每株施腐熟羊粪有机肥20 kg,移栽1个月后即9月1日进行不同施肥处理,施肥量按苗期∶营养生长期∶生殖生长期∶壮果期以2∶3∶3∶2的比例分配,分别定量于每月离根周围20 cm左右开穴施用,其他管理均按照正常大田管理标准。
1.2 主要测定指标
香蕉苗田间定植后,待幼苗长出第一片新叶作为植株第1叶,用记号笔标记植株和叶位,每15 d每处理随机选取3株带回实验室测定。用软尺或直尺测量主茎每个完全展开的功能叶最大叶长(叶片基部至顶端长度,结果以平均值表示),单位cm;最大叶宽(叶片最宽处的宽度,结果以平均值表示),单位cm;干重,105 ℃下杀青30 min,再80 ℃烘至恒重后称重,单位g。
1.3 数据处理
用SPSS 22.0、Sigmaplot14.0及Excel 2017软件处理数据。
1.3.1 巴西蕉果实形态参数与产量间的关系模型 为了模型的通用性,2018年7月至2019年7月施肥处理的数据用于分析香蕉叶片主要形态参数,2019年7月至2020年7月处理的数据用于模型检验。
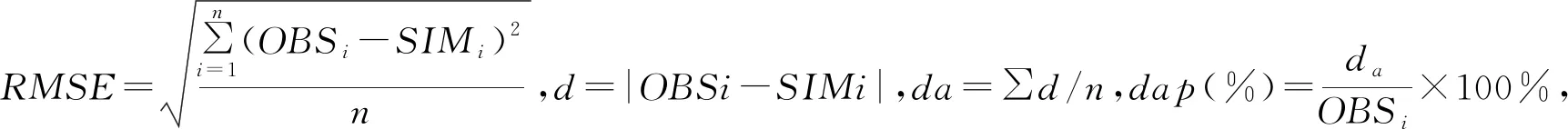
2 结果与分析
2.1 不同氮水平处理的不同叶位叶片最大叶长、叶宽、干重比较
香蕉叶片对其营养生长和果实产量非常重要,是整个植株生长的基础,对于提高果实产量作用较大。试验结果看出,不同氮水平处理下相同叶位叶片最大叶长、叶宽及干重都是随着施氮量的增加而增加,对照的最大叶长、叶宽及干重几乎都是显著低于施肥处理(见表1至表3)。

表1 不同氮水平处理的巴西蕉幼苗不同叶位叶片最大叶长比较

续表1

表2 不同氮水平处理的巴西蕉幼苗不同叶位叶片最大叶宽比较
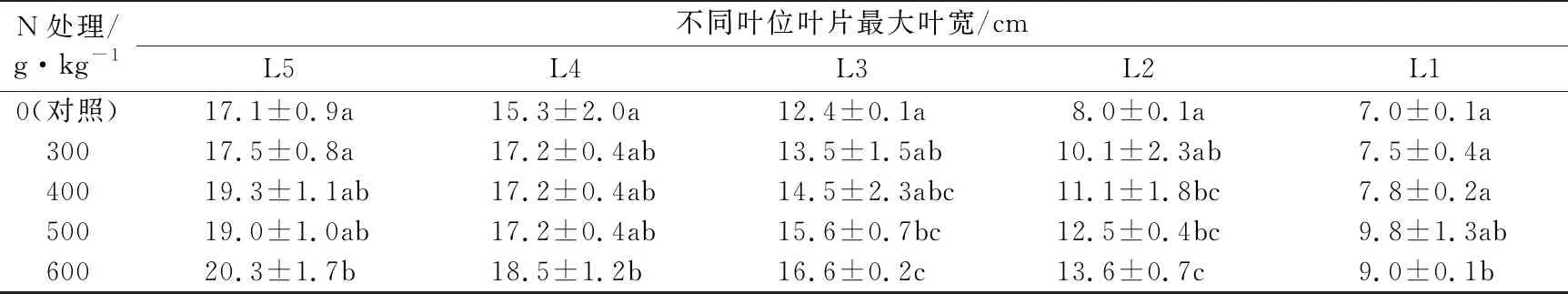
续表2
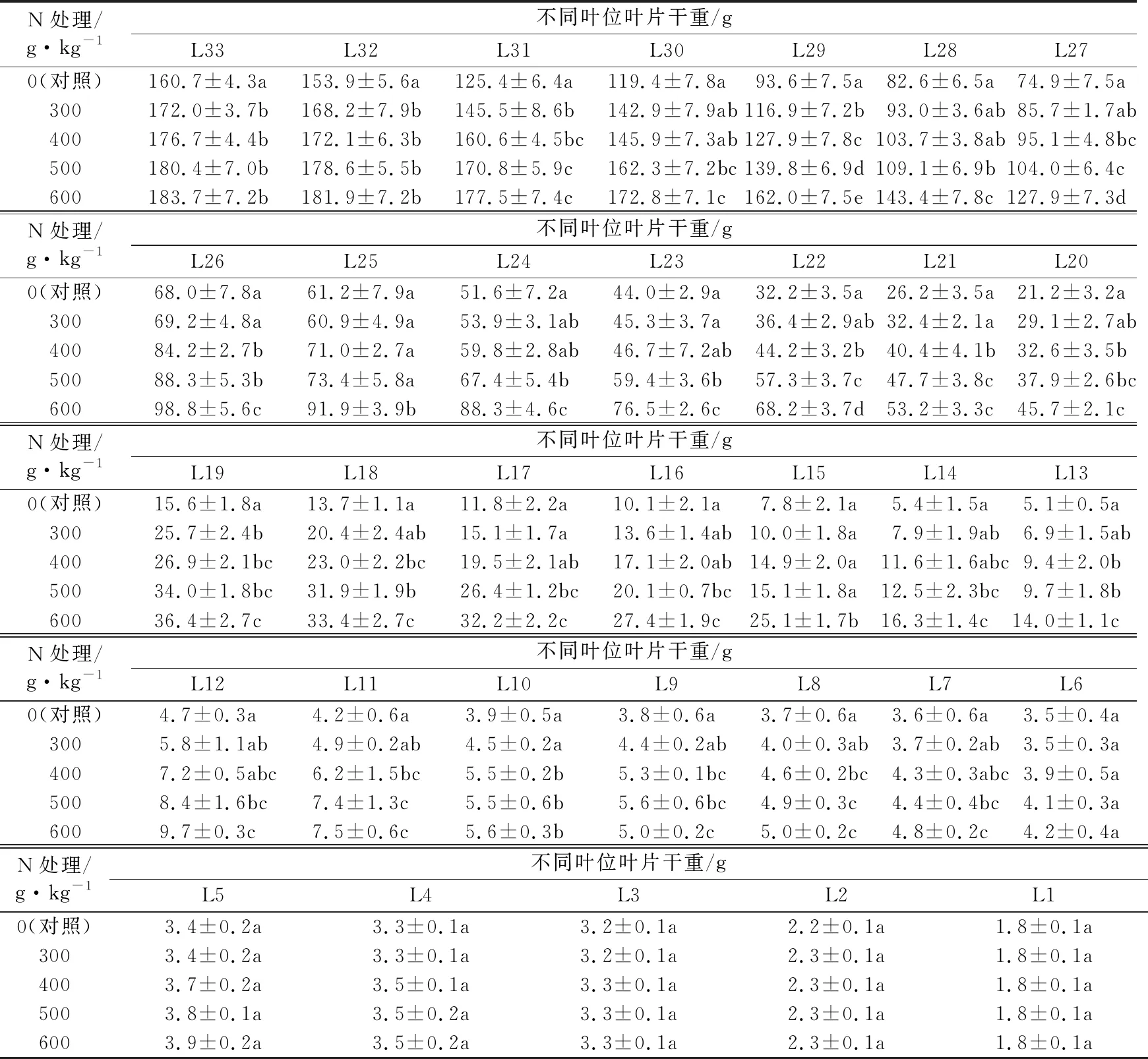
表3 不同氮水平处理的巴西蕉幼苗不同叶位叶片干重比较
2.2 香蕉叶片比叶重、最大叶长及最大叶宽的模型构建
比叶重[10](Specific blade weight,SLW)是衡量叶片光合作用性能的参数之一,用单位叶面积的叶片干重来表示。因其测定方法简便、精确,常被用作品种或者品系间的比较,也是品种的重要遗传参数。香蕉大田定植后会抽出叶片33片左右,香蕉叶片面积148~12 748 cm2,叶片干重1.8~230 g,比叶重50~82 g/cm2。由图1可知,叶片比叶重随着叶位升高先降低后增加。本文中的比叶重用2段函数来表示,数据分析结果表明,第i片叶的比叶重由如下公式计算:
SLW(i)=LDW(i)/ALB(i)(1≤i≤33),SLW(i)= c0+c1LP(i)( 1≤i≤20),SLW(i)= c2+c3LP(i) (21≤i≤33),式中,SLW(i)表示第i片叶的比叶重,LDW(i)表示第i片叶的干重(g),ALB(i)表示第i片叶的叶面积(cm2);LP(i)表示第i片叶的叶位。
叶面积可以借鉴刘永霞等[23]建立的巴西蕉叶面积与叶长的关系:ALB(i)=-38.177+0.778LL(i)×LW(i) (1≤i≤24),ALB(i)=-76.014+0.767LL(i)×LW(i)(25≤i≤33), 式中,LL(i)表示第i片叶的叶长;LW(i)表示第i片叶的最大叶宽。
叶片叶长随着生物量的增加而增长,处理间无显著性差异(见图1),不同叶片叶长与叶片生物量的关系可表示为:LL(i)=a0+a1LDW(i)+a2LDW(i)2(1≤i≤33)。

图1 不同氮水平处理的巴西蕉比叶重随叶位及叶长随叶干重的变化规律
不同叶位叶片最大叶长与叶宽的关系见图2,叶片叶长随着叶宽的增加而增长,处理间无显著性差异,其关系可表示为:LW(i)= b0+ b1LL(i)+ b2LL(i)2(1≤i≤33) ,式中,b0、b1、b2模型参数为模型参数。

图2 不同氮水平处理的巴西蕉叶片叶宽随叶长的变化规律
上述公式中,a0、a1、a2、b0、b1、b2、c0、c1、c2、c3均为模型参数,模型参数均由试验数据获得(见表4)。

表4 各模型参数值及其统计检验
由以上模型及结合刘永霞等[23]建立的叶面积模型,在已知巴西蕉某叶片的叶位、叶片生物量及已知参数a0、a1、a2、a3、b0、b1、b2、c0、c1、c2、c3后,就可以得到此株不同叶位的叶片的比叶重、最大叶片长度、最大叶片宽度和叶面积。
2.3 模型检验
采用2019—2020年独立试验资料对所建巴西蕉叶片形态参数模型进行检验,结果表明,叶片形态结构参数叶长、叶宽,及比叶重的实测值与模拟值的吻合程度较好(见图3、图4和表5),各模型r值在0.91~0.95之间(p<0.001),各模型的dap均小于15%,表明模型精度较好。
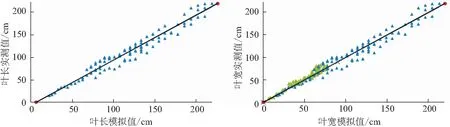
图3 2019—2020年巴西蕉叶长和叶宽测量值与模拟值比较

图4 2019—2020年巴西蕉比叶重测量值与模拟值比较
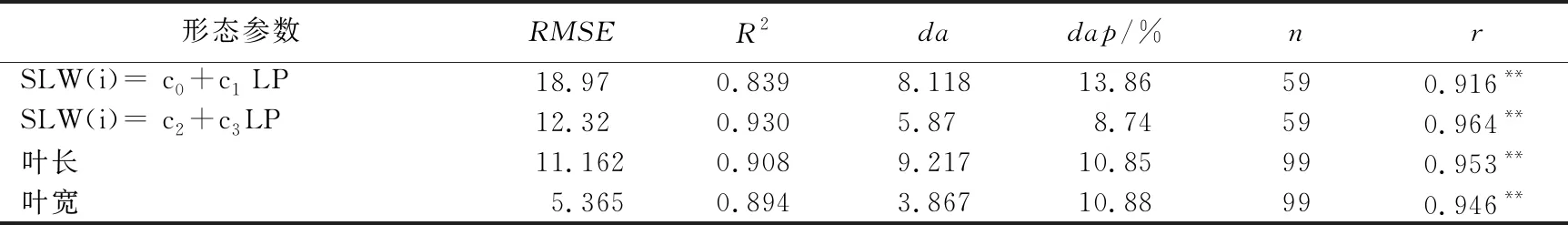
表5 巴西蕉叶片主要形态结构模型观察值与模拟值比较的统计参数
3 结论与讨论
本文分析了巴西蕉比叶重与叶位、主茎不同叶位最大叶片长与生物量、最大叶长与最大叶宽的关系,构建了基于生物量的香蕉叶片主要几何属性模型:比叶重与叶位的关系可用线性关系,叶长与叶重、叶宽与叶长的关系可用二次曲线方程表达。采用独立的试验资料检验模型,比叶重与叶位的模拟值与实测值的根均方差分别为18.97和12.32;叶长与叶重、叶宽与叶长的模拟值与实测值的根均方差分别为11.16和5.36,各模型的dap<15%,所建模型精确度较高,具有一定的机理性和普适性。
本研究通过分析香蕉叶片形态参数与生物量的定量关系,将香蕉生长模型和形态模型结合,建立了基于生物量的香蕉叶片形态结构参数模型,通过生物量、比叶重等指标将作物生理过程与作物形成模型相结合,不断强化对作物生理机制、品种基因型与环境响应研究[23],这也是香蕉功能—结构模型的重要内容之一。
氮素是调控植物叶片光合能力的最有效因子之一,适量施用氮肥可增加叶片含氮量,提高叶片的光合速率,延缓叶片光合功能的衰老进程[24]。本试验中随着氮素水平的提高,香蕉同叶位的最大叶长、叶宽及干重均有不同程度的增加,大田香蕉叶色也要更绿一些,也说明施氮增加了香蕉单株叶绿素含量,从而增加了单株光合作用效率,叶片生物量干重随供氮水平的升高而升高。这与王晓琳研究[24]相同,氮元素会使植物体可溶性蛋白含量增加,进而促使核酮糖1,5-二磷酸羧化酶含量的增加及羧化能力的提高,从而提高光合速率。但氮素不足和过量均不利于植物生物量的积累,氮素不足导致叶片光能捕获、电子传递和RuBP 羧化速率受抑制,氮素过量会引起叶片光合作用的气孔限制,影响PSII光化效率,进而抑制光合作用效率。
比叶重对叶片厚度、形状、质量有直接的影响,在一定程度上反应了叶片截获光的能力,也用来反映叶片的季节性变化。康博文等[25]研究表明刺槐、山杨、黄蔷薇、狼牙刺、柠条和紫穗槐比叶重变化趋势是单峰式,即生长初期和末期低,中期即7月比叶重最高;而本研究的香蕉则相反,即生长初期和末期高,中期即1月是峰谷,比叶重最低。分析其原因,刺槐、山杨、黄蔷薇、狼牙刺、柠条和紫穗槐的生长季节是5—10月,在北方,7月正好是北方生长季中光照最强、日照时间最长、气温最高的月份;而香蕉生长期11个月,从第一年7月到第二年6月,1月海南省澄迈县温度最低、光照减弱、日照时间减少、光合作用能力降低,因此香蕉1月的比叶重最低。卫新菊等[26]研究表明,施氮提高了苜蓿现蕾期的比叶重,本文中低氮处理的香蕉比叶重低于高氮处理,研究结果一致。另外,香蕉叶片的光合作用与比叶重、生物量的积累有直接关系,下一步拟将香蕉光合作用作为模型研究的重点之一,进一步研究光分布—光合作用—同化物的分配的关系,以增强香蕉功能—结构模型生理机制。
生物量的形成受品种和环境因素如光照、温度、水分、施肥等综合因素的影响[17],而生物量作为香蕉生长发育的一个重要指标,对香蕉植株形态具有重要作用,是香蕉生长模型和形态模型有机结合的桥梁。本研究结果表明,香蕉叶片长度和生物量关系存在二次曲线关系,这与陈昱利等[27]研究的冬小麦叶长和生物量存在线性关系有所不同,这也反映了不同品种的特性。此外,香蕉植株形态结构除了受氮素影响外,还受品种、水分、种植期、种植密度等多种因素影响,下一步拟将品种、水分和种植密度作为重点,研究此环境中的形态结构参数模型。香蕉叶片还包含叶柄及叶鞘,我们只研究了叶片的长、宽、叶面积、比叶重,下一步拟将叶柄、叶鞘形态及其生物量也纳入香蕉功能—结构模型,以完善香蕉叶片形态结构模型。
