铅镉污染矿区优势植物叶表型变异分析及综合评价
2021-06-23曹晶潇陆素芬韦岩松韦联强黄启堂
曹晶潇, 陆素芬, 韦岩松, 韦联强, 黄启堂
(河池学院 化学与生物工程学院, 河池 546300)
长期以来,陆生植物的表型可塑性受到植物生态学界的持续关注[1]。异质性是自然系统的基本特征,表型可塑性是植物适应异质生境的重要生态对策[2-3]。随着人类经济社会的发展,土壤-植物系统中重金属污染问题日益加剧,土壤中重金属的类型及浓度也表现出异质性斑块状分布格局[4]。学者们就异质重金属环境中植物的生长发育及适应性开展了一系列研究,如异质性铅污染环境中蛇莓的研究,异质性铜污染环境中苦草的研究,异质性镉环境中积雪草的研究[5-7];但大多数集中在对入侵植物的研究上,对非入侵植物研究较少。禾本科植物是一类广泛分布的植物,具有生长量大、抗逆能力强等特征,可以弥补现有土壤污染修复植物的缺点和不足,是较理想的植物修复资源[8]。目前对该科植物进行的研究多集中在地域分布、气候和土壤理化因子所带来的遗传多样性方面[9-10],对土壤重金属铅(Pb)、镉(Cd)污染的地区仍缺乏足够的表型可塑性研究。植物叶片在长期进化过程中对环境变化敏感且对气候变化的响应表现出表型可塑性[11],叶性状作为植物的重要特性之一,直接影响到植物的基本行为和功能,能够反映植物适应环境变化所形成的生存对策[12]。
广西西江上游河池地区铅锌作业密集,包括Pb、Cd在内的重金属污染历史遗留问题突出。在已发布的报道中,南丹、环江等地区土壤Pb、Cd污染已经严重影响人体健康[13-14]。因此,研究以南丹县车河、茶山、大厂镇等尾矿库周边自然生长的3种优势植物粽叶芦[Thysanolaenalatifolia(Roxb.)Honda]、类芦[Neyraudiareynaudiana(kunth.)Keng]和芒草(MiscanthussinensisAnderss)为研究对象,重点分析叶性状的变异情况,以及在重金属Pb、Cd异质环境中的适应性。运用统计学分析方法阐明不同植物叶表型变异来源及综合适应性,为矿区土壤重金属污染的生态恢复、植物资源开发、利用和保护提供支撑。
1 材料与方法
1.1 采样点概况
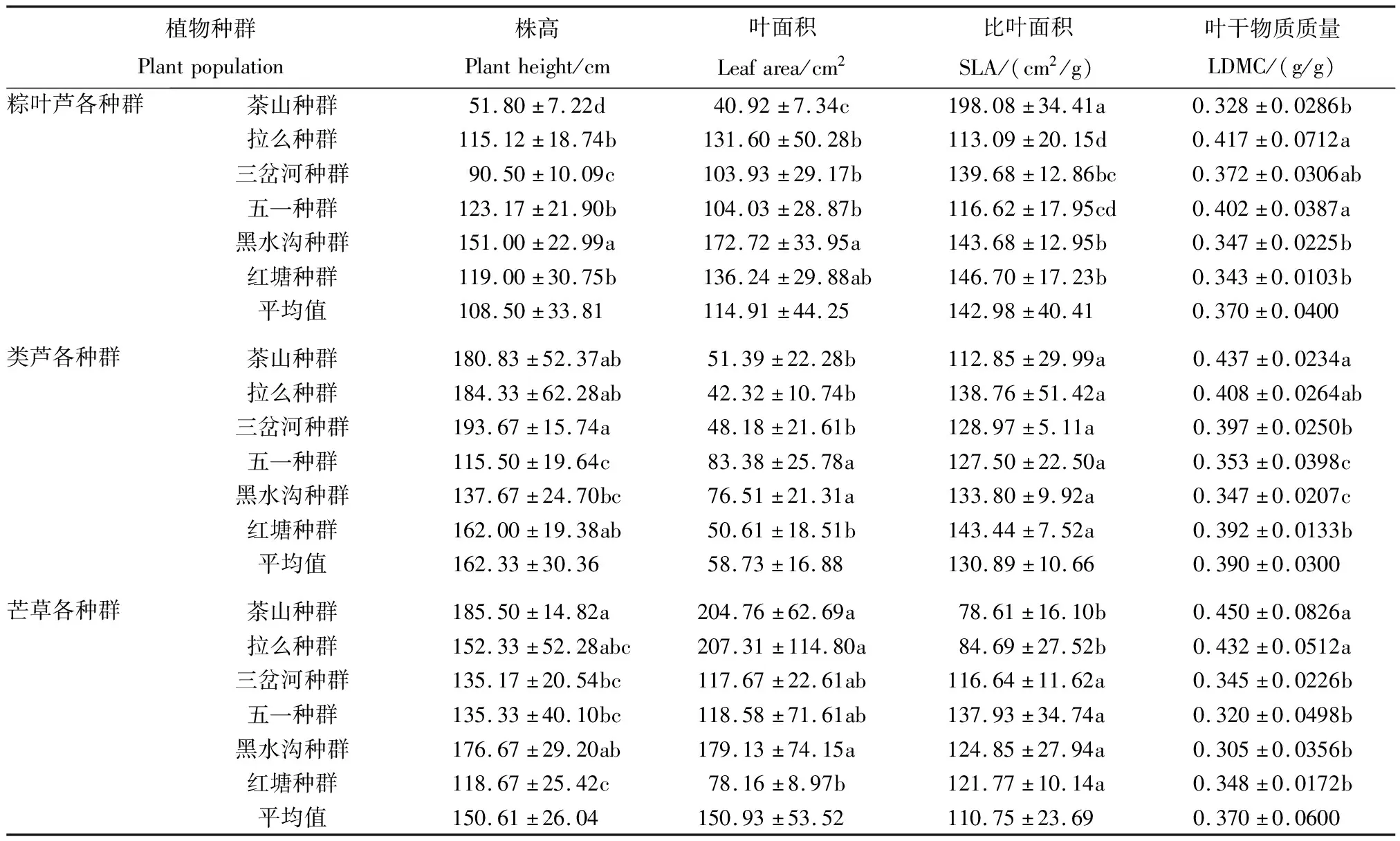
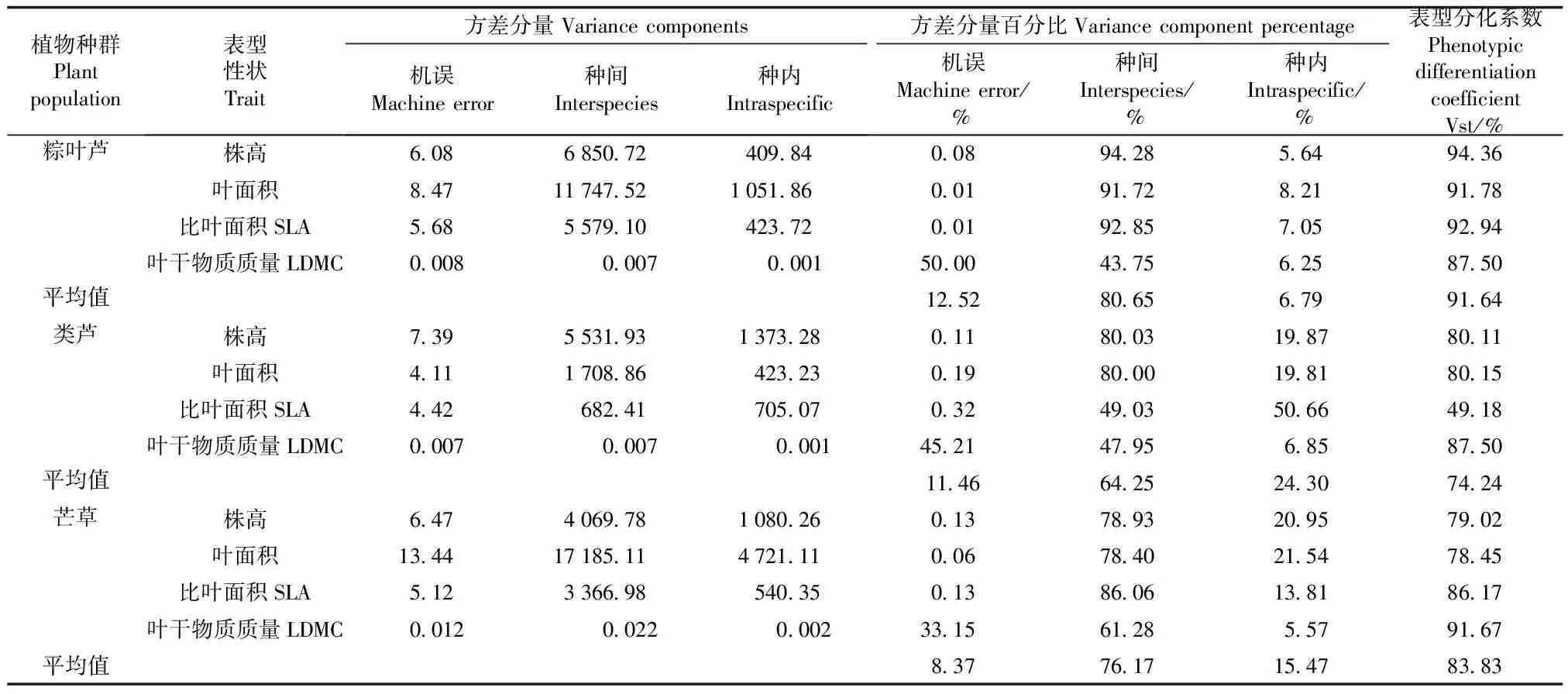
研究区域位于南丹县大厂、车河镇矿区带,土壤Pb、Cd 均值高达5 856和91.87 mg/kg[15],矿区主要出产锑、锌、铜等有色金属。采样区域常见植物有棕叶芦、类芦、芒草、百背枫、蜈蚣草、五月艾等,多分布在尾矿库采样区域的斜坡草垛上。各采样点的基本情况见表1。该地区土壤中的Pb、Cd含量均显著高于《土壤环境质量-农用地土壤污染风险管控标准(试行)(GB 15618—2018)》中对应风险筛选值(pH≤5.5、6.5 表1 采样点土壤理化性质 植物样品采集:于2018 年 7月,在矿区采摘3种植物(棕叶芦、类芦、芒草)各6个种群,每个植物种群随机选取外观无明显病虫害6株植株,选取3个完全伸展没有明显虫害的叶片,总计324片,剪下置于装有干燥剂的自封袋中,编号放入保温箱(4 ℃,内部有冰袋)中保持黑暗环境,带回实验室进行分析。 土壤样品的采集:采用“S”型土壤采样布点法取各植物种群生长地表层土壤混合样,带回实验室后铺开放置在室内,风干。采用四分法取样分别过100目、60目和20目筛,分别装封于自封袋中,贴上标签贮存在干燥环境中,进行后续重金属Pb、Cd以及相关理化性质的测定,土壤理化性质测定结果见表 1。 先用去离子水清洗叶片表面灰尘,再将装植株的自封袋中充满水,置于 5 ℃黑暗环境下储藏 12 h,取出后迅速吸干叶片表面水分,天平称重其读数即为叶片的饱和鲜重;采用美国 CID公司生产的 CI-300手持式激光叶面积仪对其叶面积进行测定;将叶片放入60 ℃的烘箱烘干48 h后,取出称重即为干重。植物比叶面积(SLA)和叶干物质质量(LDMC)以及变异系数的计算公式见文献[16];采集过程中,利用不锈钢直尺分别对3种植物各6个种群、每个种群各6株植株株高进行测量,重复3次,测量精确到0.1 cm。 3种植物(粽叶芦、类芦、芒草)各种群表型变异特征采用巢式方差分析,计算各植物种间表型分化系数(Vst)和变异系数,具体计算公式见文献[17];Vst表示种群间变异占遗传总变异的百分比,表明表型变异在种群间贡献的大小。 运用SPSS 22.0软件进行种群间、种群内方差分析、皮尔逊相关性分析,通过F检验的Duncan统计辨识种群间的差异性。 对 3种植物各 6个种群 4个表型性状指标的平均值、标准差及多重比较结果见表 2。粽叶芦各种群表型性状:株高除红塘、五一种群之间差异不显著外,其余的种群间差异显著;叶面积指标拉么、三岔河、五一种群之间无显著差异,茶山种群与其他 5个种群间差异显著;SLA、LDMC 2个指标中黑水沟、红塘种群不存在显著差异。类芦各种群:五一种群株高与其他种群间存在显著差异,叶面积、LDMC指标中五一、黑水沟种群与其他种群差异显著,SLA指标各种群无明显差异。芒草各种群:红塘种群的株高、叶面积与其他 5个种群差异明显,茶山、拉么种群的 SLA、LDMC与其他种群差异明显。 表2 3种植物各种群表型性状分析 3种植物各6个种群表型性状指标在种群内、种群间方差分量和表型分化系数情况见表3。就各性状的平均方差分量而言,3种植物(粽叶芦、类芦、芒草)的表型性状种群间的平均方差分量大于种群内,说明其表型性状的种群间变异远大于种群内变异;3种植物各种群的表型分化系数结果显示:3种植物的表型变异种群间的变异都大于种群内的变异,即变异主要存在于不同种群间,种群内变异保持得较为稳定;3种植物(粽叶芦、类芦、芒草)平均表型分化系数分别为 91.64%、72.24%和83.83%。 表3 3种植物表型性状的方差分析及表型分化系数分析 3 种植物各种群表型性状变异系数见表 4。粽叶芦各表型性状种群内平均变异系数为 16.10%,变异幅度为 8.86%~25.43%。株高、叶面积 2个指标的变异系数大于种群内部平均变异系数,SLA、LDMC变异系数小于种群内部平均变异系数;由粽叶芦不同种群间所有指标的平均变异系数可知,拉么、五一种群这2个种群的变异系数均大于平均变异系数,表明拉么种群、五一种群的表型变异和多样性较为丰富;红塘、茶山种群的变异系数比较接近平均变异系数,表明这2个种群的多样性大体一致;三岔河、黑水沟种群的变异系数小于粽叶芦平均变异系数,表明这2个种群的表型变异较弱,多样性也较低。 表4 3种植物各种群表型性状变异系数分析 类芦各种群表型变异性状的种内平均变异系数为 19.31%,变异幅度为 6.46%~34.82%。株高、叶面积 2个指标的变异系数大于种群内部平均变异系数,SLA、LDMC变异系数小于种群内部平均变异系数;类芦各种群间所有指标的平均变异系数(表4),茶山、拉么种群 2个种群的变异系数均大于平均变异系数,表明这2个种群的表型变异和多样性较为丰富;三岔河、五一、黑水沟、红塘种群的变异系数小于类芦各表型平均变异系数,表明这4个种群的表型变异较弱,多样性也较低。 芒草各种群表型性状在种群内平均变异系数为 22.13%,变异幅度为 11.49%~36.39%。株叶面积的变异系数大于种群内部平均变异系数,株高、SLA、LDMC变异系数均小于种群内部平均变异系;芒草不同种群间所有指标的平均变异系数可知,拉么、五一、黑水沟 3个种群的变异系数均大于平均变异系数,表明这3种群的表型变异和多样性较为丰富;茶山、三岔河、红塘种群的变异系数小于芒草平均变异系数(22.13%),表明这 3个种群的表型变异较弱,多样性也较低。3种植物各种群表型性状平均变异系数大小排序均为叶面积>株高> SLA> LDMC,说明所测性状中LDMC这一性状比较稳定。 利用所测土壤因子与3种植物表型性状进行相关性分析,结果(表5)表明:粽叶芦各种群中株高与土壤含水量呈极显著负相关,叶干物质质量与土壤含水量呈显著负相关。土壤因子对粽叶芦表型性状影响程度有一定差异,按照相关系数均值大小依次排列为:土壤总磷>含水量>pH值>总氮>总Pb>全钾>有机质>总Cd;土壤因子对类芦表型性状影响按照相关系数均值大小依次排列为:土壤pH值>全钾>含水量>有机质>总Cd>总磷>总氮>总Pb;芒草各种群株高与土壤pH值呈极显著负相关、与土壤总磷呈显著负相关,土壤因子对芒草表型性状影响按照相关系数均值大小依次排列为:土壤pH值>总Cd>有机质>总Pb>总氮>全钾>总磷>含水量。 表5 3种植物表型性状与土壤因子间的相关系数分析 种群是生物适应环境的基本单位。一般来说,种群内的变异体现种群的稳定性,分布在群体间的变异真正反映了群体在不同环境中的适应状况,其大小在某种程度上说明了该生物对不同环境适应的广泛程度,即种群间变异越大,该生物适应的环境越广[18-19]。3种植物(粽叶芦、类芦、芒草)表型分化系数分别为91.64%、72.24%和83.83%,高于珙桐(37%)[20]、北沙柳(26.54%)[21]等自然定居在未受Pb、Cd污染土壤中的植物表型分化系数,与红椿(80.73%)[22]、夏蜡梅(89.30%)[23]等植物接近。与自然定居未遭受重金属污染的植物地理居群相比,3种矿区植物种群间表型分化水平高,为了应对土壤中Pb、Cd含量的差异,形成了稳定表型遗传特征。原因可能是本研究中的3种禾本科植物地下茎丰富,无性系繁殖发达,相比起异花授粉的植物而言,种群间交流较为容易,种群更新较为快速;其次,地理隔离会造成种群间交流不频繁[22],野外调查发现,3种植物种群分布距离较近,且由于采样区域平均海拔较低,各种群间没有高山阻挡,相反那些叶表型变异主要来源于种群内的植物,由于种群分布距离远,地理隔离使得种群间表型变异保持在相对较低水平,而促进了种群内部的遗传分化。因此,与那些叶表型分化系数接近的植物种群相似[22-23],3种植物种群间的环境异质性程度大于种群内,种群间变异是3种植物表型变异的主要来源。 变异系数是衡量物种表型性状变异程度的重要指标,值越大,说明物种表型变异幅度越大,可以使种群整体更加适应环境的变化[24]。研究中的3种优势植物的变异系数变化范围较大(表4),说明它们在自然条件下都具有极强的生存和适应能力,其中,芒草的变异系数最大(表4),就矿区这种重金属污染环境而言芒草的适应能力较其他2种植物强,这与田美虹等[25]研究结果类似。 综上,研究中的3种优势植物各表型性状的多样性水平高,种群间表型多样性大于种群内,表型变异丰富,反映了3种植物具有广泛的环境适应性。 植物在不同的生态环境中,往往会以不同的形态适应不同环境,这对提高该物种的抗逆性和适应环境的能力十分有利[26]。3种植物表型性状变异分析结果(表2)显示相同植物不同种群间性状差异显著,说明它们对异质环境的趋异适应。研究发现粽叶芦、芒草表型性状与土壤pH值、含水量、TP存在显著相关性,推测长期生长在重金属污染环境中的植物形成了较强的抗逆性,因此重金属含量并不是影响矿区优势植物适应能力的关键因素,相反在重金属污染下,土壤本身的pH值是影响矿区优势植物适应能力的关键因素。这可能是因为土壤pH值是土壤化学性质的综合反映,土壤中各营养元素的转化、释放,镁、铁、钙等微量元素的有效性,有机质的合成与分解、微生物的活动以及重金属生物有效性都与土壤pH值有关[27]。因此,在考虑对矿区重金属污染进行植被修复时,比起采用化学方法降低土壤中重金属总量而言,调整土壤酸碱性以及引种抗重金属能力强且具有土壤改良作用的植物更为重要。类芦的表型性状与8个土壤因子均为无显著相关性,说明生态环境因子对其叶表型影响不大,决定其表型的主要因素可能是其自身遗传因素。 SLA和LDMC是叶片最为关键的性状因子,SLA与植物同化率有着较为密切的关系,一般而言,SLA 值越低表示该植物本身对环境资源的利用能力越强,对所获得资源的保存能力也越强,能够更好地适应贫瘠的环境[28],LDMC能够很好地反映植物对养分的保有能力[29]。对外界恶劣环境抵抗能力较强的植物,往往具有较高的资源利用效率(SLA小、LDMC较高),且对环境养分具有较好的保存能力,可通过慢速的养分循环来适应环境;而资源利用能力较差的植物(SLA较大、LDMC较低),对外界不良环境的抵抗能力也较差,此类植物通常具有较高的光合生产能力,可通过快速的养分循环来求得生存[30]。SLA、LDMC两者体现了植物为获得最大化碳收获所采取的生存适应策略,是植物适应相同或不同生境所体现出的关键特征,能综合反映植物利用资源的能力,具有重要的生态学意义[31]。本研究中的3种优势植物SLA大小排序为粽叶芦>类芦>芒草,芒草的SLA值最小,表明芒草在该地区利用环境的能力较强,在矿区土壤重金属污染适应性方面表现出优势。就本研究中的3种优势植物而言,芒草表现出相对最低水平的SLA和中等水平的LDMC;类芦表现出中等水平的SLA和最大LDMC;粽叶芦表现出最高水平的SLA和中等水平的LDMC;3种植物都表现出对矿区重金属污染环境的趋同适应。3种矿区优势植物LDMC值平均值(0.37 g/g)大于滇中岩溶地区草本植物(0.27 g/g)[28]和科尔沁沙地植物(0.27 g/g)[32]。研究结果从侧面说明土壤重金属污染地区植物抵御外界干扰能力强于其他地区植物,当受到重金属胁迫时仍能表现出对养分较高的保有能力,维持体内碳同化,保证自身的正常生长,更好地适应污染环境,这对土壤重金属污染生态恢复提供了良好的植物资源支撑。 综合而言,芒草具有最高的资源利用效率,对外界不良环境的抵抗能力较强,类芦次之,粽叶芦在3种植物中抵抗不良环境能力最差;芒草在矿区土壤重金属污染环境中表现出适应性上的优势。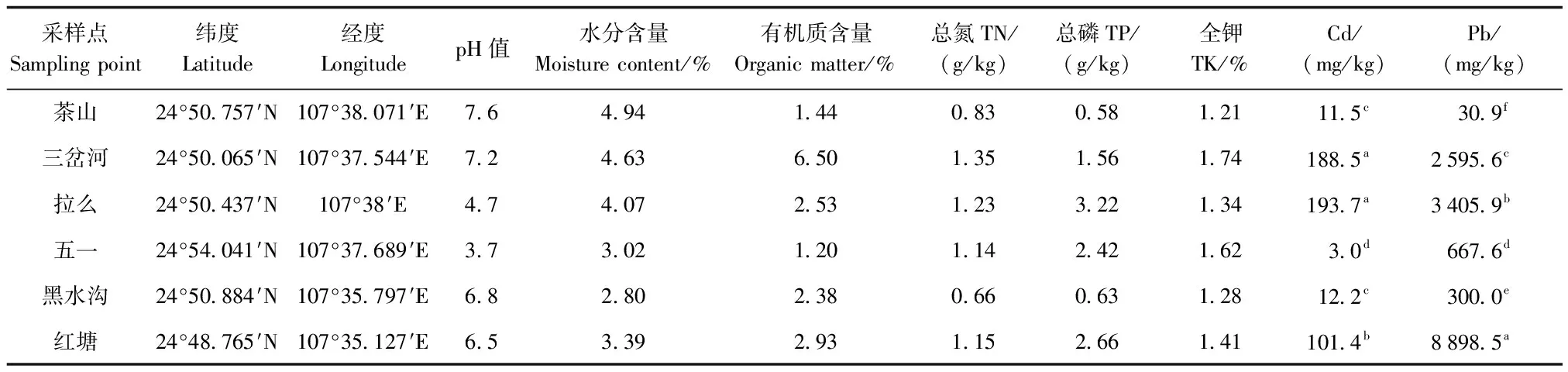
1.2 样品采集与处理
1.3 表型性状的测定
1.4 数据处理
2 结果与分析
2.1 3种植物各种群表型性状特征
2.2 3种植物各种群表型性状变异来源分析
2.3 3种植物各种群表型性状变异特征分析

2.4 3种植物表型性状与土壤因子相关性

3 讨论与结论
3.1 矿区3种优势植物的表型变异来源
3.2 矿区3种优势植物叶表型性状与土壤理化因子的关系
3.3 矿区优势植物适应环境能力比较
