空心莲子草入侵群落的昆虫丰富度和分布状况及其影响因素
2021-06-22吴昊王明霞李迎港
吴昊 ,王明霞,李迎港
1.信阳师范学院生命科学学院,河南 信阳 464000;2.中国科学院武汉植物园,湖北 武汉 430074
昆虫作为自然界数量最多、分布最广的动物群体,是全球生物多样性的重要组分;其生长发育及分布格局受到温度、湿度和地形等各类环境因子的影响(Dudgeon,2012;金倩等,2019)。前人研究发现,帽儿山溪流水生昆虫群落结构的主导性因子为海拔、水温、总磷和氨态氮(刘曼红等,2014);贺兰山地表甲虫丰富度与土壤含水量呈正相关、与海拔呈负相关(王敏等,2020);青藏高原鳞翅目昆虫的系统发育多样性随降雨量上升而增加(金倩等,2018)。此外,植食性昆虫还从寄主植物中获取营养物质以维持其种群规模(García-Robledo et al.,2011),故栖息地植物群落类型及其种群特征也是影响昆虫数量和分布的重要因素(Wardle,2006;王珊珊等,2012)。例如,浙江清凉峰的不同植物群落中步甲物种多样性明显表现为落叶阔叶林>常绿阔叶林>针阔混交林(郭瑞等,2016);对于武汉市湿地公园而言,千屈菜(Lythrumsalicaria)、再力花(Thaliadealbata)等群落中的昆虫多样性较高,而香蒲(Typhaorientalis)群落的昆虫种数最低(王珊珊等,2012)。探讨异质性生境中昆虫分布与其栖息地植物群落之间的关系,有助于深度理解物种共存及生物多样性维持机制。
近年来,迅速的全球环境变化导致生物入侵日益加剧,严重制约经济发展和环境质量,尤其威胁生态系统稳定性及生物多样性(吴昊等,2014)。入侵植物不仅对本土种产生强烈的种间竞争,还可通过改变多营养级中的食物资源种类、调节本土植物营养和化学防御水平、改变植物群落空间结构和化学信号特征等方式间接地影响昆虫分布及种群数量(Bezemer et al.,2014;王毅等,2019)。张红玉(2013)发现,紫茎泽兰(Ageratinaadenophora)入侵导致依赖于本土植物的昆虫种群减少或丧失适宜的栖息生境,其还可通过形成单优势种群落及强烈的化感作用抑制天敌昆虫自然生长。蒋智林等(2017)发现,随着紫茎泽兰入侵程度增加,群落中昆虫的多样性指数先降低后升高,而其优势度指数先升高后降低,且紫茎泽兰入侵群落昆虫多样性显著低于对照区禾本科植物群落。互花米草(Spartina alterniflora)入侵降低了盐沼湿地昆虫多样性,并显著改变昆虫功能群组成特征,导致群落中膜翅目植食性昆虫的数量和种类明显降低、昆虫分散程度增加(潘卉等,2017)。相对于本土种而言,外来入侵植物与本土昆虫的协同进化时间较短,这种时空尺度上的“非对称性”(non-parallel)对于预测入侵后效及其生物防治具有重要指导意义(Cronin et al.,2015),但目前关于大尺度异质性生境中的“入侵植物-昆虫”相关性依然亟待研究。
空心莲子草(Alternantheraphiloxeroides)又名水花生,为苋科莲子草属杂草,原产于南美洲,由于其具备强大的克隆增殖能力及水、陆两境适应性,已广泛入侵中国南方、北方20余省,给经济建设、生态环境及本土植物多样性造成极大威胁(Wu et al.,2017;吴昊等,2020)。前人围绕空心莲子草入侵植物群落结构特征、环境因子对空心莲子草天敌昆虫的影响等方面开展了大量的研究,例如,陆生型及水生型空心莲子草群落的主要伴生植物分别为马唐(Digitariasanguinalis)和双穗雀稗(Paspalumpaspaloides),且陆生入侵群落的植物多样性显著高于水生(Wu et al.,2017);气候变暖导致空心莲子草向高纬度扩散,从而加剧该入侵种对其专食性昆虫莲草直胸跳甲(Agasicleshygrophila)的“天敌逃逸”及莲草直胸跳甲对本土植物的“非靶标效应”(Lu et al.,2015);入侵群落中莲草直胸跳甲数量的主导性因子为气温、纬度和海拔(吴昊等,2019)。但大空间尺度上空心莲子草群落的昆虫发生状况及决定因素尚未见报道。本研究拟通过大尺度野外调查,阐明无机环境、空心莲子草入侵群落特征对昆虫丰富度分布的影响,以期为空心莲子草防治及其天敌资源利用提供参考。
1 研究方法
1.1 野外调查
于2019年7—8月在中国南方的湖北、湖南、广东、广西和江西等5省内空心莲子草连续入侵面积>100 m2的区域设置调查样地(介于 21°N—31°N)。每隔2个纬度梯度设置5个陆生样地,并在同纬度段生境相似的区域设置5个水生样地。共设置样地50个,分布于5省11市,其中水、陆样地各25个(具体分布如表1所示)。研究样地横跨亚热带大陆性季风气候、亚热带季风性湿润气候和亚热带季风气候等多个气候区,降水充沛、光照充足,年均气温20.34 ℃,年均降雨量1514 mm(中央气象台国家气象中心,http://www.nmc.cn),土壤质地以水稻土、红壤和砂岩土等为主。空心莲子草入侵群落分布于撂荒地、废弃工地、乡径、水田、池塘和河岸带等多类生境中。
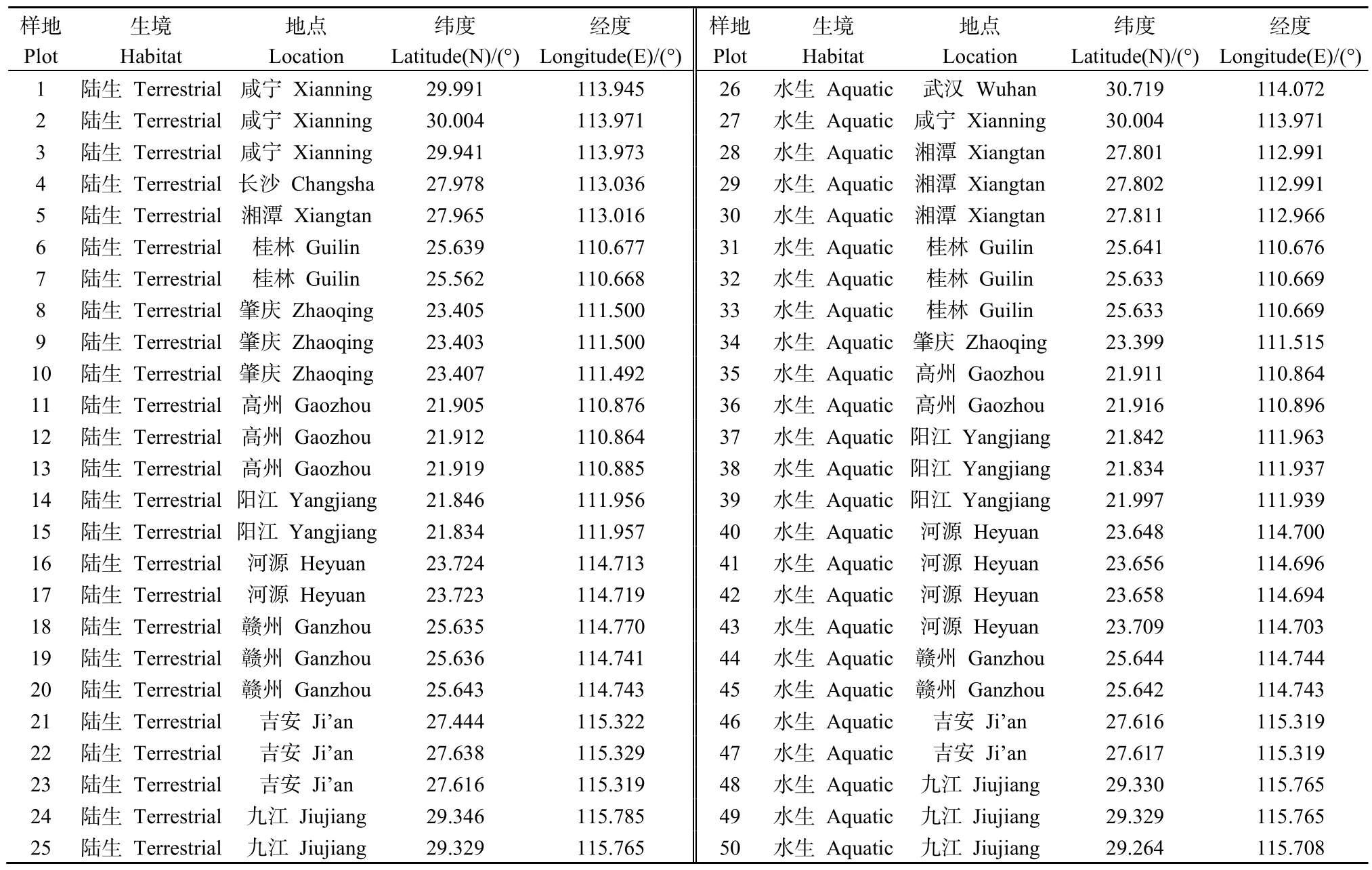
表1 空心莲子草入侵群落样地分布概况Table 1 Distributions of sampling plots in Alternantheraphiloxeroides invaded communities
每个样地面积为10 m×10 m。对于陆生生境及水田、河岸交错带等水生生境,在样地中央均匀布设3条10 m长的样带,每条样带上均匀设置5个面积为0.5 m×0.5 m的小样方。而对于池塘、水库和河流等水位较深的生境,为了便于调查,选取靠近岸边的水生型空心莲子草毯状斑块,沿水平方向连续设置3条10 m长的样带进行调查。调查时,记录各小样方中植物的物种名称、株高、盖度和多度等指标。植物调查结束后,使用可伸缩式捕虫网(直径长30 cm,网深度60 cm)沿各样地中任意一条样带横扫10网(来回扫一次算1网),将捕获的昆虫装进昆虫瓶。所有收集到的昆虫带回实验室后,利用《中国昆虫生态大图鉴》 (张巍巍等,2011)、《农业昆虫鉴定》(李照会,2002)和中国动物主题数据库(http://www.zoology.csdb.cn)、国家动物标本资源库(http://museum.ioz.ac.cn)以及美国佐治亚大学病虫害物种在线图像库(http://www.ipmimages.org)等资料按其形态特征进行物种鉴定,并统计样地中昆虫丰富度(总物种数)及各类昆虫的数量。
1.2 环境测定
利用手持式GPS定位仪(Garmin eTrex20,美国)测量样地的经度、纬度和海拔。在植物及昆虫调查结束后,对于陆生样地,沿其任意一条对角线均匀设置3个取样点,采集0—20 cm土层土壤样品约250 g,装进塑封袋,置于车载冰箱(Mobicool CF-50DC,德国)冷冻保存,及时带回实验室后利用全自动化学分析仪(Easychem Plus,意大利)测定土壤中氨态氮(N-NH4+)和硝态氮(N-NO3−)含量。对于水生样地,利用 YSI便携式水质分析仪(Professional Plus,美国)测量水质中的氨态氮和硝态氮含量。在中央气象台国家气象中心网站(http://www.nmc.cn/)获取各样地所在城市近50年的平均气温及年均降雨量数据。
1.3 数据计算
植物重要值(Importance value,IV)计算公式如下所示:

群落中某种植物总重要值即为所有样地中该植物的相对重要值之和;植物群落α-物种多样性指数计算公式如下:

式中,S为样地内的所有植物的总种数,Pi为第i种植物的相对重要值(吴昊等,2020)。
1.4 数据分析
使用SPSS 16.0软件(SPSS Inc,美国)对样地的7项环境因子(经度、纬度、海拔、氨态氮、硝态氮、年均温、年均降雨量)、空心莲子草4项种群特征(株高、盖度、多度、重要值)和群落的4项多样性指数(Patrick、Shannon-Wiener、Simpson和Pielou指数)分别与样地中昆虫丰富度进行曲线回归分析(Curve Estimation),以SPSS 16.0提供的线性、多项式、指数等11类方程建立回归模型,选择回归显著且拟合系数最高者进行讨论。利用 SPSS 16.0对水、陆两境昆虫丰富度进行独立样本t检验(Independent-samplettest)。
对于水、陆两境空心莲子草样地群落,分别建立25×7的环境因子矩阵、25×8的植物群落特征矩阵(种群形态、物种多样性指数),以及 25×19和25×15的昆虫数量矩阵,利用数量生态学软件Canoco 4.5(Microcomputer Power,美国)提供的典范对应分析(Canonical Correspondence Analysis,CCA)探讨无机环境和空心莲子入侵对异质性生境中昆虫分布状况的影响,并绘制二维排序图。利用基于499次置换的Monte Carlo前置性检验判断自变量与排序轴相关系数的显著性。CCA是一种基于单峰模型的约束性数量排序方法,其能够很好地反映多变量的综合效应并筛选出主导性因子。
2 结果与分析
2.1 昆虫分类特征
50个调查样地分布于5省11市,7—8两个月共采集昆虫标本402头(陆生样地151头、水生样地251头),鉴定为25种(表2)。其中,陆生样地昆虫8目19科19属19种,丰富度较高的为鞘翅目9科9属9种、双翅目6科6属6种,虫体数量较高的物种为莲草直胸跳甲(33头)和东亚飞蝗(33头);水生样地昆虫7目15科15属15种,丰富度较高也为鞘翅目和双翅目,均为4科4属4种,莲草直胸跳甲的虫体数量最大(134头)。水、陆两境共存的昆虫物种有9种,分别为莲草直胸跳甲、东亚飞蝗、淡胸藜龟甲、向日葵斑螟、短尾黄蟌、马铃薯瓢虫、白额高脚蛛、家蝇和黑尾黑麻蝇。独立样本t检验表明,水、陆两境的昆虫丰富度无显著性差异(t=1.206,P=0.234)。
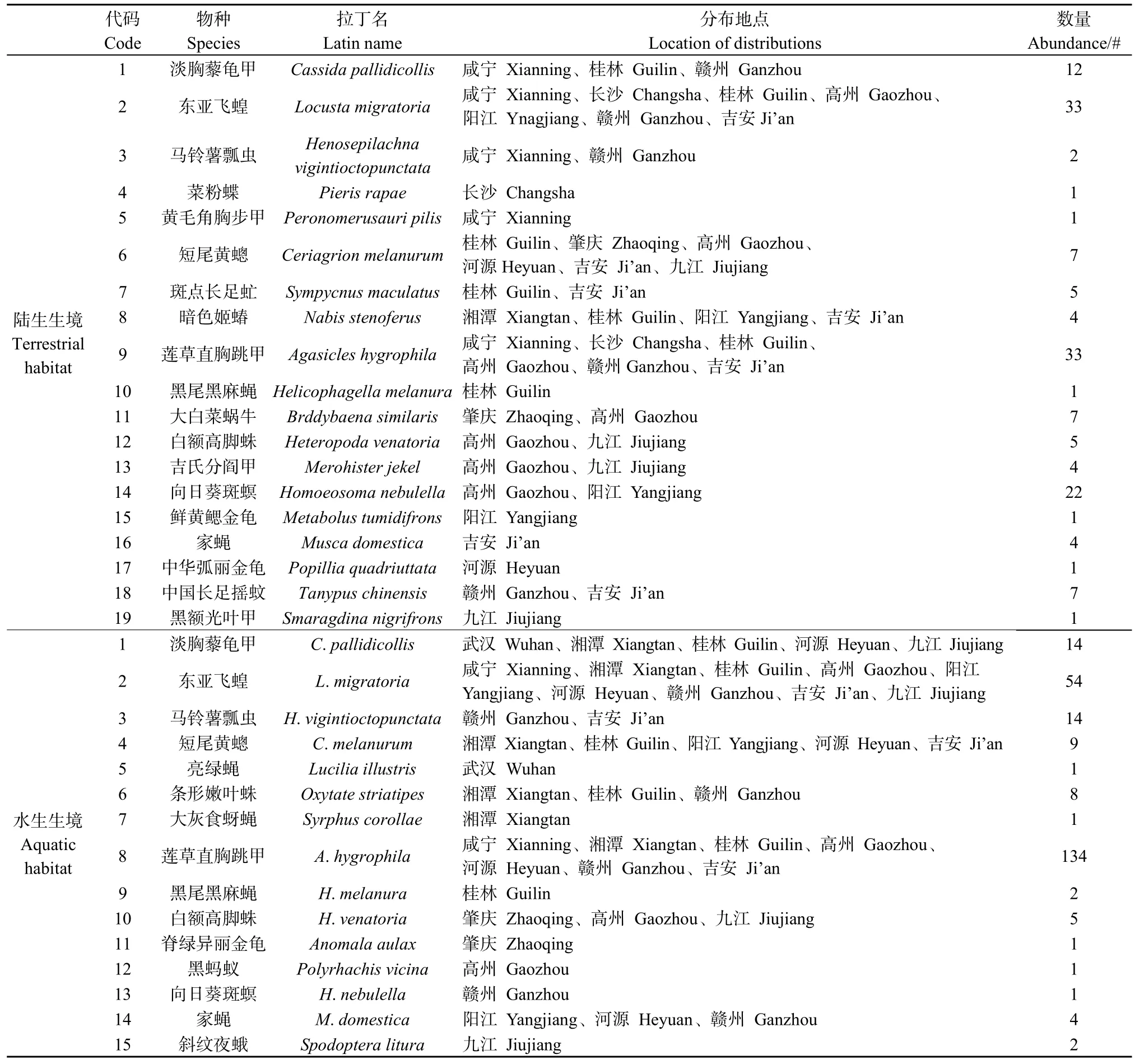
表2 异质生境中空心莲子草群落的昆虫群落结构Table 2 Insect communities’ structures of A.philoxeroides communities in heterogeneous habitats
2.2 环境因子及植物群落特征对昆虫丰富度的影响
回归分析表明,陆生样地昆虫丰富度与土壤硝态氮呈显著的线性拟合关系(r=0.247,P=0.012),其随硝态氮升高而明显增加;水生样地昆虫丰富度与纬度和年均气温均呈极显著的二项式拟合关系(r=0.373,P=0.006;r=0.471,P=0.001),与海拔呈极显著的三项式拟合关系(r=0.416,P=0.009),其随着纬度、年均气温和海拔的增加均表现为“先上升—后下降”趋势(图1)。水、陆两境空心莲子草种群特征(株高、盖度、多度和重要值)与昆虫丰富度间均无显著性关系。陆生样地中,昆虫丰富度与群落 Patrick丰富度指数之间具有显著的三项式拟合关系(r=0.359,P=0.023),其随Patrick指数增加呈“上升—下降—上升”波动趋势;昆虫丰富度与群落 Pielou均匀度指数呈显著的线性拟合关系(r=0.159,P=0.048),其随 Pielou指数上升而明显下降(图2)。水生样地的4个物种多样性指数与其昆虫丰富度之间均无显著性关系。
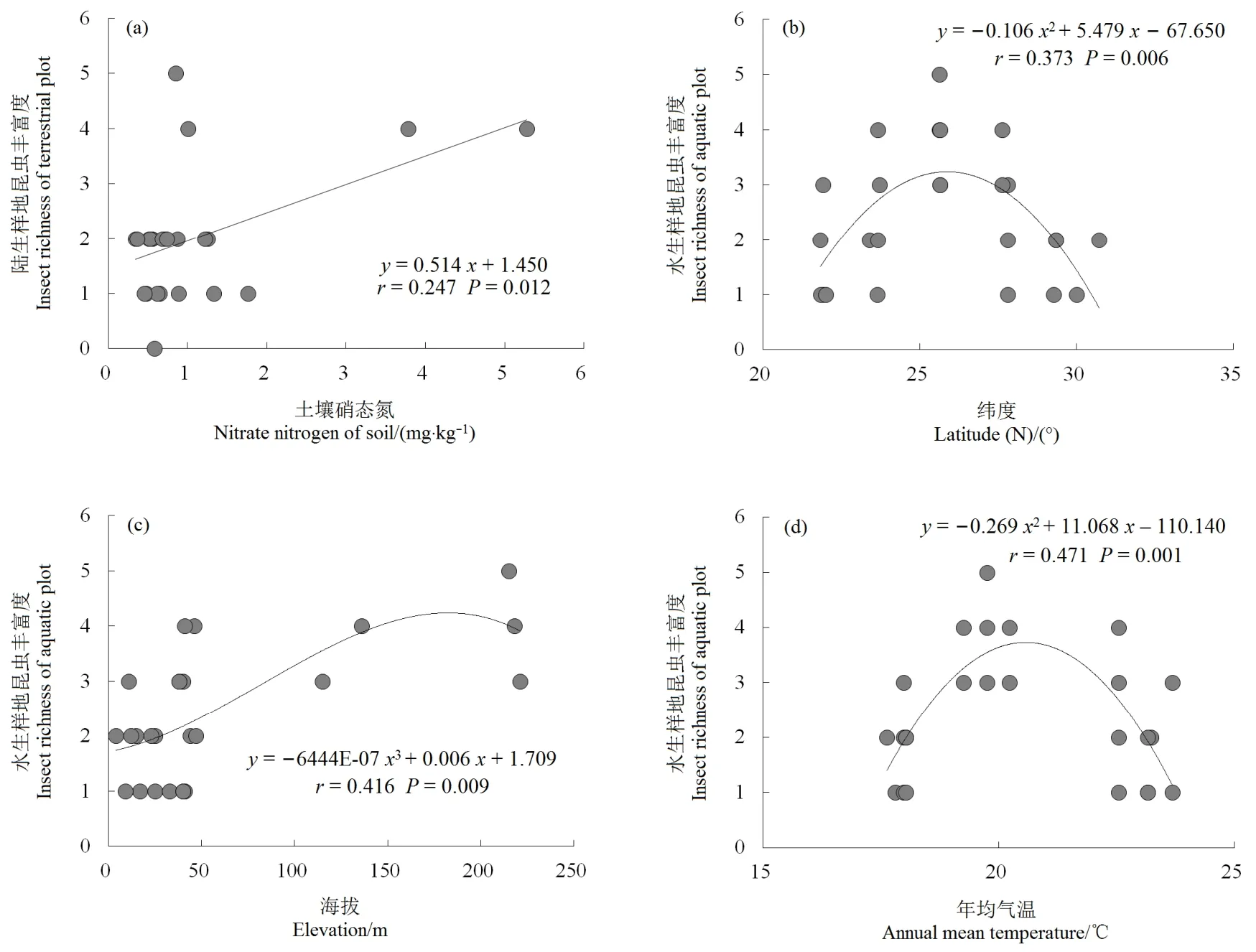
图1 环境因子与昆虫丰富度的回归分析Fig.1 Regression analysis between environmental factors and insect richness
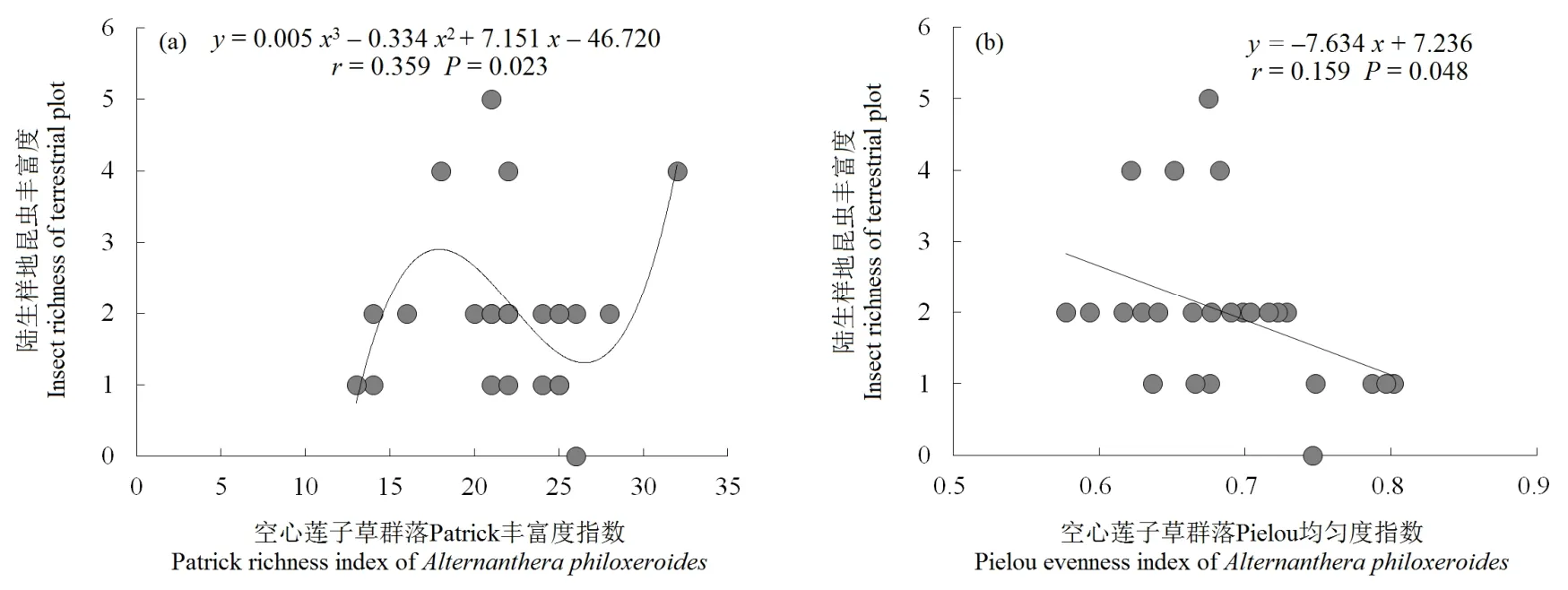
图2 入侵群落物种多样性指数与昆虫丰富度的回归拟合Fig.2 Regression analysis between species diversity indices and insect richness in invaded communities
2.3 环境因子对昆虫分布的影响
CCA排序轴计算结果如表3所示,陆生及水生样地前两个排序轴对“环境因子-昆虫分布”关系解释的累计贡献率分别为 44.9%(1轴 24.0%,2轴20.9%)和55.5%(1轴35.2%,2轴20.3%),采用前两轴数据作CCA二维排序图(图3)。
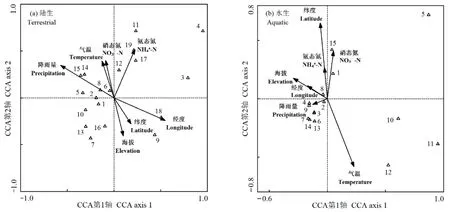
图3 陆生(a)及水生(b)样地中昆虫分布与环境因子的CCA二维排序图Fig.3 Two-dimensional CCA ordination diagram of insect distributions along environmental factors in terrestrial (a) and aquatic (b) plots
结合表3及图3a可知,决定陆生样地昆虫分布的主导性环境因子为年均降雨量、经度和氨态氮。其中,对CCA 1轴起到显著影响作用的因子为年均降雨量(r= −0.572,P<0.01)和经度(r= −0.549,P<0.01),决定 CCA 2轴的主导性因子为氨态氮(r=0.518,P<0.05)。即沿着CCA 1轴从左往右,经度逐渐增加、降雨量逐渐减少;沿着CCA 2轴由下往上,土壤氨态氮含量逐渐增加。陆生样地的昆虫物种沿环境梯度的 CCA排序较为分散,其分布受到多个环境因子的影响。如:中国长足摇蚊(代码18)和莲草直胸跳甲(代码 9)趋向分布于经度较高的区域;向日葵斑螟(代码14)和鲜黄鳃金龟(代码15)趋向分布于年均降雨量较大的区域;大白菜蜗牛(代码11)、中华弧丽金龟(代码17)和黑额光叶甲(代码19)的分布与土壤氨态氮含量具有密切正向关系,而斑点长足虻(代码 7)则趋向分布于氨态氮含量较低的区域。马铃薯瓢虫(代码 3)和菜粉蝶(代码 4)远离排序图中心,表明这两种昆虫物种的分布极少受无机环境因子的影响。
7项环境因子均未对水生样地CCA 1轴产生显著性影响;决定CCA 2轴的因子为纬度(r=0.651,P<0.01)和年均气温(r=−0.584,P<0.01)(表 3、图3b)。即沿着CCA 2轴由下往上,纬度逐渐上升、年均气温逐渐降低。相对于陆生而言,水生样地的昆虫分布较为集中。黑蚂蚁(代码12)趋向分布于低纬度的温暖地带,斜纹夜蛾(代码15)趋向分布于温度较低的高纬度区域;亮绿蝇(代码5)、白额高脚蛛(代码10)和脊绿异丽金龟(代码11)受环境影响较小;多数昆虫趋向分布于中等环境梯度区域。水、陆两境中,东亚飞蝗(代码2)均位于排序图中心位置,表明其生态幅宽、生态适应性强,可在空心莲子草入侵群落中广泛存在。
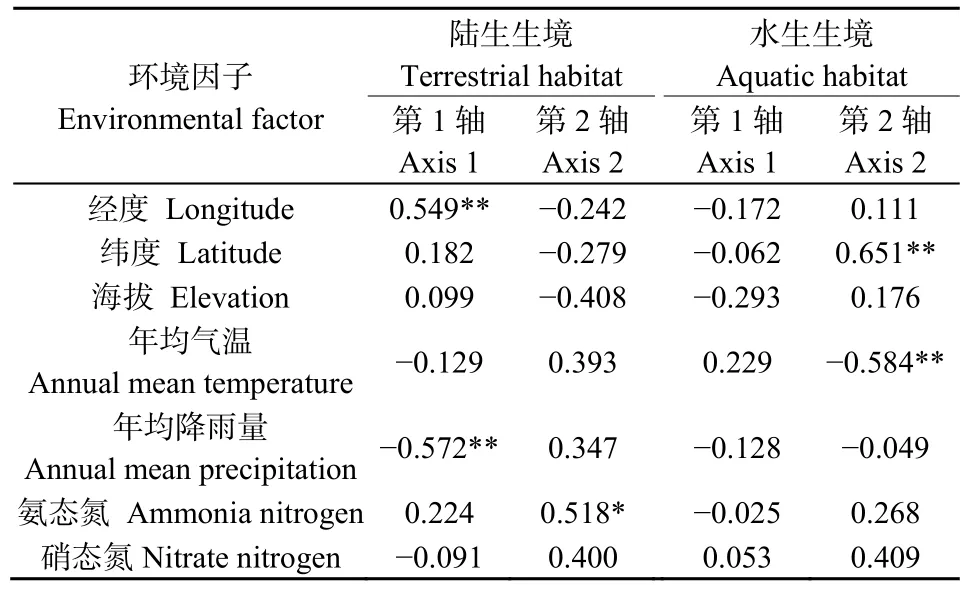
表3 环境因子与CCA前两个排序轴的相关系数(r)Table 3 Correlation coefficient (r) between environmental factors and CCA first two ordination axes
2.4 植物群落对昆虫分布的影响
CCA排序轴计算结果如表4所示,陆生及水生样地前两个排序轴对“昆虫分布-植物群落特征”关系解释的累计贡献率分别为39.7%(1轴22.1%,2轴17.6%)和60.1%(1轴38.0%,2轴22.1%),采用前两轴的数据进行CCA二维排序如图4所示。
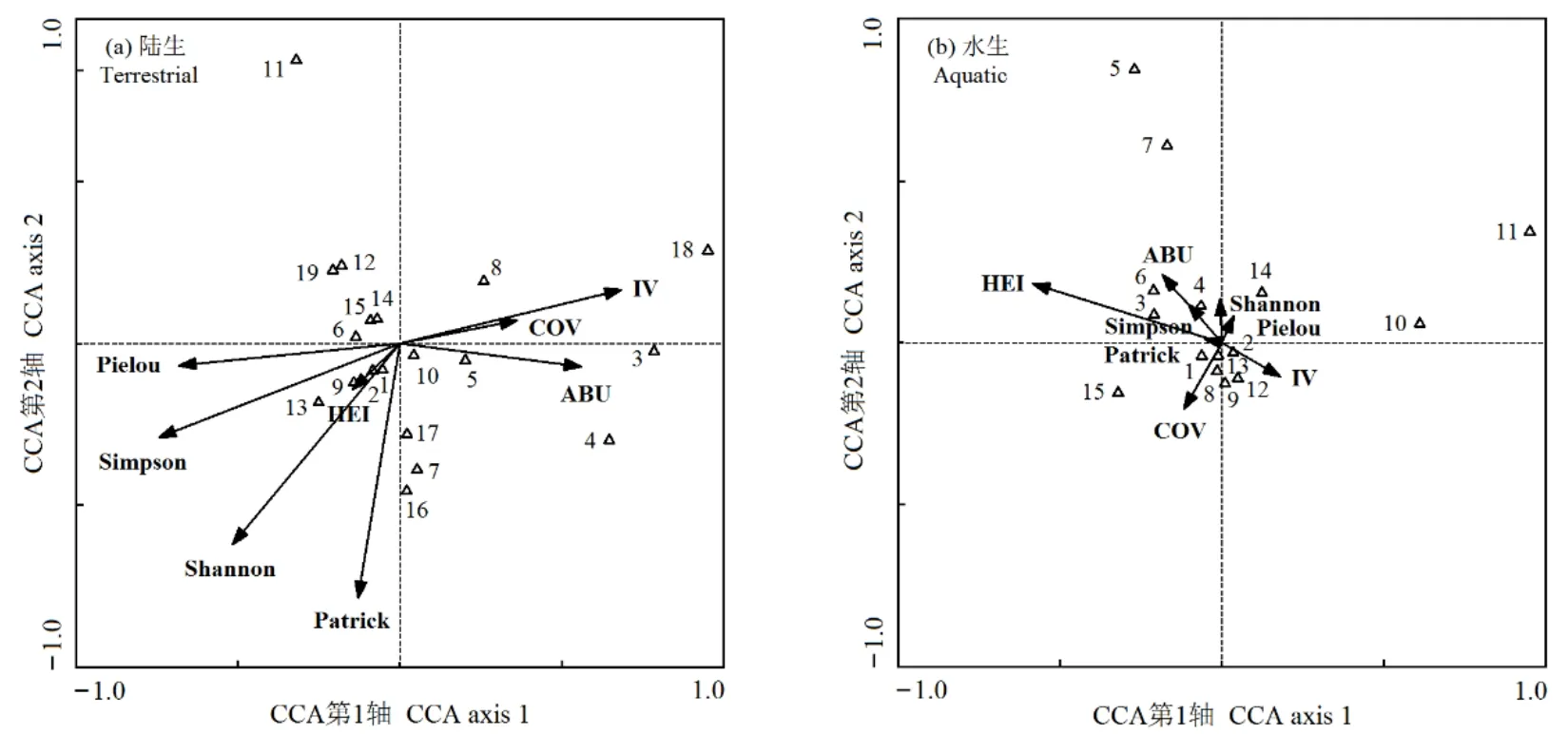
图4 陆生(a)及水生(b)生境中昆虫分布与植物群落特征的CCA二维排序图Fig.4 Two-dimensional CCA ordination diagram of insect distributions along plant community traits in terrestrial (a) and aquatic (b) habitats
物种多样性指数及空心莲子草多度和重要值均对陆生样地的昆虫分布产生了显著性影响(表 4和图4a)。其中,CCA 1轴的主导性因子为Simpson多样性指数(r= −0.744,P<0.01)、Pielou均匀度指数(r= −0.683,P<0.01)、Shannon-Wiener多样性指数(r= −0.517,P<0.05)、空心莲子草重要值(r=0.683,P<0.01)和空心莲子草多度(r=0.558,P<0.01);CCA 2轴的主导性因子为Patrick丰富度指数(r= −0.786,P<0.01)和 Shannon-Wiener多样性指数(r= −0.620,P<0.01)。马铃薯瓢虫(代码 3)、菜粉蝶(代码4)和中国长足摇蚊(代码18)倾向分布于空心莲子草入侵态势严重的区域;而包括优势物种莲草直胸跳甲(代码 9)在内的大多数昆虫趋向分布于中等植物多样性水平的区域;大白菜蜗牛(代码11)的分布受植物群落特征影响极小。
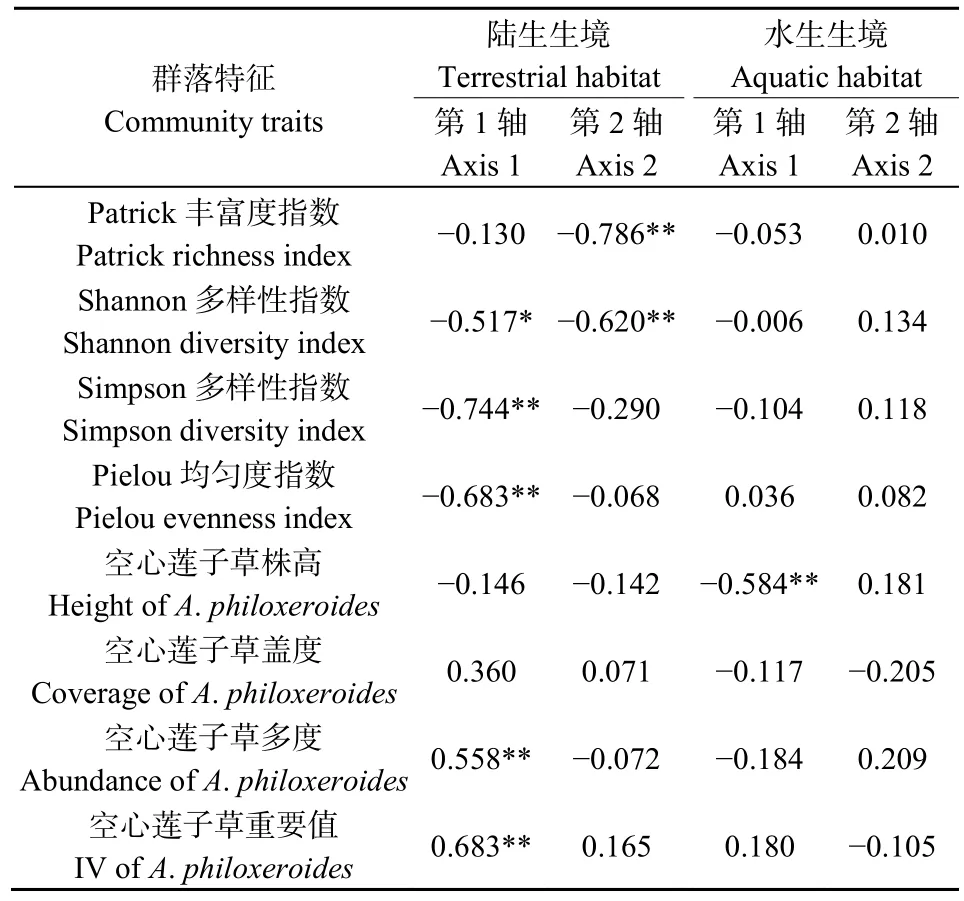
表4 植物群落特征与CCA前两个排序轴的相关系数(r)Table 4 Correlation coefficient (r) between plantcommunity traits and CCA first two ordination axes
对于水生样地,仅空心莲子草株高对CCA 1轴起到显著决定作用(r= −0.584,P<0.01),其余因子均未对 CCA排序轴产生显著影响。水生样地昆虫分布的集中程度极高,均趋向分布于中等株高的空心莲子草群落中。亮绿蝇(代码5)、大灰食蚜蝇(代码7)和脊绿异丽金龟(代码11)等3种昆虫的分布基本不受植物群落特征影响。
3 讨论
3.1 陆生环境中昆虫与降雨量、经度和土壤氮素的关系
昆虫的生长发育及其行为、分布与环境密切相关,植物群落中的昆虫丰富度状况一定程度上反映了昆虫种群之间以及“环境-植物-昆虫”间的互作关系(王毅等,2019)。本研究发现,年均降雨量、经度和土壤氨态氮是决定陆生空心莲子草群落中昆虫结构与分布的主导因素。随着降雨量升高,土壤相对湿度增加,这将对陆生生境中昆虫的取食强度、幼虫存活率、生态位分化程度及种群分布模式造成显著影响(顾永征等,2010;贺奇等,2013)。经度间接影响昆虫分布,其常与降水量、气温、区域性干旱指数等因素具有共变效应(吴昊等,2019),且在中国境内,自西向东随着经度增加,商业贸易活动渐趋频繁、氮沉降增加,这有助于空心莲子草入侵蔓延,也为生防昆虫莲草直胸跳甲的种群扩增提供了理想栖息地(Lv et al.,2007;吴昊等,2014),但高经度地区入侵植物形成的“生态避难所”也会导致摇蚊等害虫大量滋生(Wu et al.,2019)。陆生昆虫丰富度与土壤硝态氮呈显著正相关(图1),是由于与氨态氮相比,大多数陆生植物以硝态氮作为其主要氮源,且硝态氮在土壤中的移动速度较快,植物能更加迅速地吸收利用硝态氮以促进自身生长,从而为植食性昆虫提供充足的食物来源及优良的栖息环境(Wu et al.,2019;宋明华等,2020)。氨态氮累积容易导致植物光合电子传递链断裂及其膜系统的功能障碍(Wu et al.,2017),使得陆生CCA排序图中大多数昆虫趋向分布于中、低氨态氮含量的生境中(图3a)。
3.2 水生环境中昆虫与温度、纬度和海拔的关系
水生样地昆虫丰富度随年均气温、纬度的增加均呈“先上升—后下降”单峰回归模式(图1),这与其CCA排序结果高度吻合(图3b)。昆虫生长发育在一定范围内随温度的升高而加快,但气温过高则会制约昆虫性腺的发育及能量代谢,导致昆虫性比偏移、产卵数量和卵化率降低(李建宇等,2013;蒋丰泽等,2015)。纬度等大尺度空间上的地理因子本身不会直接影响水生生物,其通过引起温度和降水的变化间接作用于水域系统(Dudgeon,2012)。低纬度地区的过高温度导致昆虫生殖率下降、虫体内有益共生菌解体,而高纬度带的低温则破坏昆虫的滞育节律(辛蓓等,2019)。回归分析中,水生样地昆虫丰富度随海拔上升而增加,是由于海拔升高导致区域尺度下的水温、降水等因素波动明显,这种变化带来的生境异质性为更多具有独立生态功能的昆虫提供了栖息空间(魏慧玉等,2020)。相对于陆生生境而言,本研究中水生生境的昆虫分布受无机环境影响较弱,可能是由于陆生群落在很大程度上受到土地利用类型的影响,并且人类活动极大地干扰了陆生昆虫的自然分布(李嘉昊等,2018);而河流、湖泊等生境具有良好的水体连通性使得其营养元素迁移性及周转率均较高,缓解了水环境对植物群落及昆虫种群的限制作用(Alahuhta et al.,2018)。
3.3 昆虫与植物多样性的关系
本研究发现,陆生样地昆虫丰富度随植物群落Patrick指数增加呈“上升—下降—上升”波动趋势。这是因为在Patrick指数增加初期,较高的植物丰富度能够支持更加多样化的昆虫群落(朱慧等,2008),但当虫口密度增大到一定程度时,种间竞争加剧导致虫体死亡率明显增高,其丰富度下降(冀佳悦等,2014)。随着Patrick指数继续增加,本土植物占据更多生态位,其对空心莲子草入侵的生物抗性增强,即降低了莲草直胸跳甲的产卵场地及食物来源,从而削弱优势种莲草直胸跳甲对其他昆虫的竞争,使得群落中昆虫丰富度有所上升(Byun et al.,2015;吴昊等,2019)。本研究中,陆生昆虫丰富度与Pielou均匀度指数呈相关,是由于植物分布越均匀,则其为昆虫生存营造的生境异质性越低,均质化的环境不利于维持昆虫多样性(Meziere et al.,2015)。陆生CCA排序图中,马铃薯瓢虫、菜粉蝶趋向分布于Pielou指数低且空心莲子草入侵态势高的区域。前人研究表明,马铃薯瓢虫具有很强的寄主选择性,喜欢取食茄科及苋科植物(刘永超等,2018),空心莲子草或是其理想食物来源之一;菜粉蝶等蝶类昆虫的分布与城市距离关系密切,距主城区越远,其种群密度越大(和秋菊等,2008),而空心莲子草群落常大面积发生于远离城区的农田和撂荒地(吴昊等,2020),可为菜粉蝶提供良好的栖息地。水生 CCA排序图中,大部分昆虫趋向分布于中等空心莲子草株高的群落中,可能是由于水生型中等高度空心莲子草的茎节距离更利于栖息在其上的昆虫产卵、孵化及幼虫发育(马瑞燕等,2004)。
3.4 环境变化下的“入侵植物-昆虫”互作态势
全球环境变化将导致部分生态幅较广的昆虫的潜在分布区逐步扩大,而对生境变化敏感的昆虫物种则可能受到限制(Hof et al.,2015)。前人研究表明,增温可削弱本土植物对于空心莲子草入侵的生物抗性,使得空心莲子草在气候变暖情形下通过生物量累积、增强种间竞争力等途径加剧入侵态势(吴昊等,2020);但高温不利于生防昆虫莲草直胸跳甲的生存,且导致使其幼虫化蛹率显著降低(胡中昀等,2014)。此外,全球高纬度、高海拔地区的增温幅度更加明显,故空心莲子草将会在这些潜在适生区内由于“天敌逃逸”以及莲草直胸跳甲对本土植物“非靶标效应”的增强而进一步入侵扩散(Lu et al.,2015),因此,应更加关注空心莲子草群落中昆虫丰富度变化及筛选生防昆虫资源。值得注意的是,本研究发现中国长足摇蚊趋向分布于空心莲子草入侵程度较高的区域,蚊子的大量发生势必威胁人类健康,故未来研究中关于“入侵植物-媒介昆虫-人类健康”级联关系也应引起足够重视。
4 结论
(1)陆生空心莲子草群落昆虫丰富度的主导性因子为土壤硝态氮和植物多样性,昆虫分布的主导性因子为降雨量、经度和植物群落特征。水生空心莲子草群落昆虫丰富度的主导性因子为纬度、气温和海拔,昆虫分布的主导性因子为空心莲子草株高。
(2)对于陆生空心莲子草入侵严重的群落,可人为补植本土植物,通过适当提高植物多样性维持多种昆虫共存,以实现“以虫治草”绿色防控;同时可在西部低经度的入侵区域,人工培养并释放莲草直胸跳甲,确保该专食性生防昆虫具有较高的野外种群密度。
(3)应加强对水生入侵生态系统的取样调查,防止由于昆虫分布与水环境的弱关联性导致空心莲子草“天敌逃逸”加剧而进一步入侵蔓延。此外,还应重视对空心莲子草群落中东亚飞蝗、中国长足摇蚊等有害昆虫的监测,防止其借助空心莲子草的“生态避难所”效应对农业生产和人类健康造成严重危害。
