不同种质茶籽脂质代谢特征分析
2021-06-19陈明杰杜正花秦健恒李若钰于朝夕郭丽
陈明杰,杜正花,秦健恒,李若钰,于朝夕,郭丽,3*
不同种质茶籽脂质代谢特征分析
陈明杰1,杜正花2,秦健恒1,李若钰1,于朝夕1,郭丽2,3*
1.信阳师范学院生命科学学院/河南省茶树生物学重点实验室,河南 信阳 464000;2.福建农林大学园艺学院,福建 福州 350002;3.中国农业科学院茶叶研究所,浙江 杭州 310008
叶籽两用茶树种质的研发利用对于茶产业结构调整、茶园综合效益提升具有特殊意义,因此深入研究现有茶树种质与茶籽相关性状特征,可为叶籽两用茶树的遗传育种提供基础信息。从福建、云南和湖南采集了31份同年结实茶籽,对种仁的含油率和脂肪酸组成特征进行测定。结果表明,不同茶树茶籽含油率存在2倍以上的差异;从茶籽油脂中检测到10种脂肪酸组分,包括棕榈酸、棕榈油酸、顺-11-十六碳烯酸、硬脂酸、油酸、顺-11-十八碳烯酸、亚油酸、亚麻酸、二十烷酸和二十烯酸。油酸是茶籽中的主要脂肪酸,占脂肪酸总量的42.1%~59.2%;其次为亚油酸,占脂肪酸总量的18.9%~32.8%;饱和脂肪酸含量占15.9%~20.8%,以棕榈酸为主。进一步分析各种质的脂肪酸组成特征,发现茶籽中的FAD2和FAD3活力在种质间存在较大差异。在此基础上,探讨了如何利用这些发现来选择育种,以进一步改善茶籽的油脂性状。
茶籽;含油率;脂肪酸;脂肪酸去饱和化酶
茶树()是传统的叶用经济作物,新梢通常被用来加工成各种茶制品。据2019年统计,我国茶园种植面积293.33万hm2,茶叶总产量260万t。随着全球茶叶市场供求关系的变化,国内茶产业供大于求,导致茶叶价格低迷,经济效益下滑,影响了茶产业链参与者的利益和积极性[1]。另一方面,随着经济发展和人民生活水平的提高,我国食用油消费量持续增加,每年需进口大量油菜籽、大豆等来满足食用油需求。据统计,我国大豆对外依存度已超过85%,并且进口渠道不多,主要依赖北美和南美的几个大豆生产国,国际油料市场供应波动对国内食用油市场影响较大。于此同时,我国茶籽资源利用率较低。虽然茶籽平均年产量可达2 175 kg·hm-2[2],但随着无性系茶园的推广,茶树有性繁殖被扦插繁殖大规模替代,导致茶籽需求进一步萎缩,茶籽除少量用于繁殖或被混入油茶籽提取油脂外,大部分被遗弃[3]。因此开发利用叶籽两用茶树种质资源,不仅可以促进我国茶产业提质增效,也有助于保障我国食用油的供应稳定。
目前叶籽两用茶的发展思路是立足于现有茶园,通过减少种植密度和调整茶园管理来实现[4-5]。目前推广的茶树品种基本上是基于叶的生产性状进行培育,较少考虑茶籽的农艺和经济性状。从长远看,为了最大程度发挥茶树种植的经济和社会效益,需要在育种环节有针对性地开发一系列在叶和茶籽两个方面均表现优异的新种质。种仁含油率和油脂脂肪酸组成是决定作物油料产量和质量的重要方面,也是叶籽两用的茶树育种中亲本选配的重要指标。由于复杂的人文、历史、消费习惯,以及茶树的环境适应性差异等多方面因素,我国主要茶区茶树品种繁多,既有国家级、省级优良无性系良种,也有地方品种和群体种。多样化的种植模式给当前的叶籽两用茶树转换带来挑战,同时也说明叶籽两用茶树的开发利用应因地制宜,充分了解和利用现有的茶树种质资源。
现有研究表明,以叶用为主的茶树茶籽与油茶籽的脂肪酸组成分布特征具有明显差异,主要表现在油酸、亚油酸、亚麻酸含量及占比上[1,6-7]。-6和-3脂肪酸去饱和化酶是亚油酸、亚麻酸形成的关键酶[8],但其在不同茶树种质中的差异尚不明晰。基于此,本研究采集同年结实的31份茶籽,分析其含油率和脂肪酸组成,研究茶籽含油率的种质间差异及脂肪酸的组成特征,查明脂肪酸去饱和化酶在不同种质间的差异,旨在为叶籽两用茶树的选育提供参考依据。
1 材料与方法
1.1 试验材料
31份茶籽采摘于2016年,福建、湖南和云南的种质分别有26、3份和2份(表1)。茶籽经干燥脱壳后取出果仁,将来自同一品种的3个茶籽合并碾碎,作为1个生物学重复。每个品种设置3个生物学重复。碾碎后的茶籽采用真空冷冻干燥机干燥。
1.2 试验方法
1.2.1 油脂衍生化处理
油脂的衍生化处理参考文献[9]并略作修改。称量10 mg茶籽粉末装入8 mL螺口玻璃管中,加入1 mL 5%(/)硫酸-甲醇溶液、25 μL 0.2%(/)丁基化羟基甲苯、300 μL甲苯和75 μg十七烷酸甲酯作为内标,在95℃金属浴加热90 min后冷却至室温,加入1 mL 0.9%(/)氯化钠溶液和3 mL正己烷,剧烈振荡30~60 s,1 000离心10 min。将上层有机相转移到干净玻璃管中,下层水相用3 mL正己烷重复提取1次,合并有机相,氮气吹干后加入1 mL正己烷溶解。
1.2.2 气相色谱-质谱和气相色谱-离子火焰检测器分析
气相色谱-质谱和气相色谱-离子火焰检测器分析采用Shimadzu的多维气相色谱质谱分析仪(MDGC/GCMS-2010)。毛细管柱采用RESTEK公司的RT-2560柱(0.25 mm×30 m×0.25 µm)。氦气作为载气,流速为1 mL·min-1,分流比为10∶1。进样口和检测器温度分别设定为240℃和220℃。柱温箱的起始温度为150℃,以2℃·min-1的速度升温到200℃,保持5 min。每份样品分别用气相色谱-质谱和气相色谱-离子火焰检测器检测,气相色谱-质谱数据用来做脂肪酸定性分析,气相色谱-离子火焰检测器数据用来做脂肪酸定量分析。
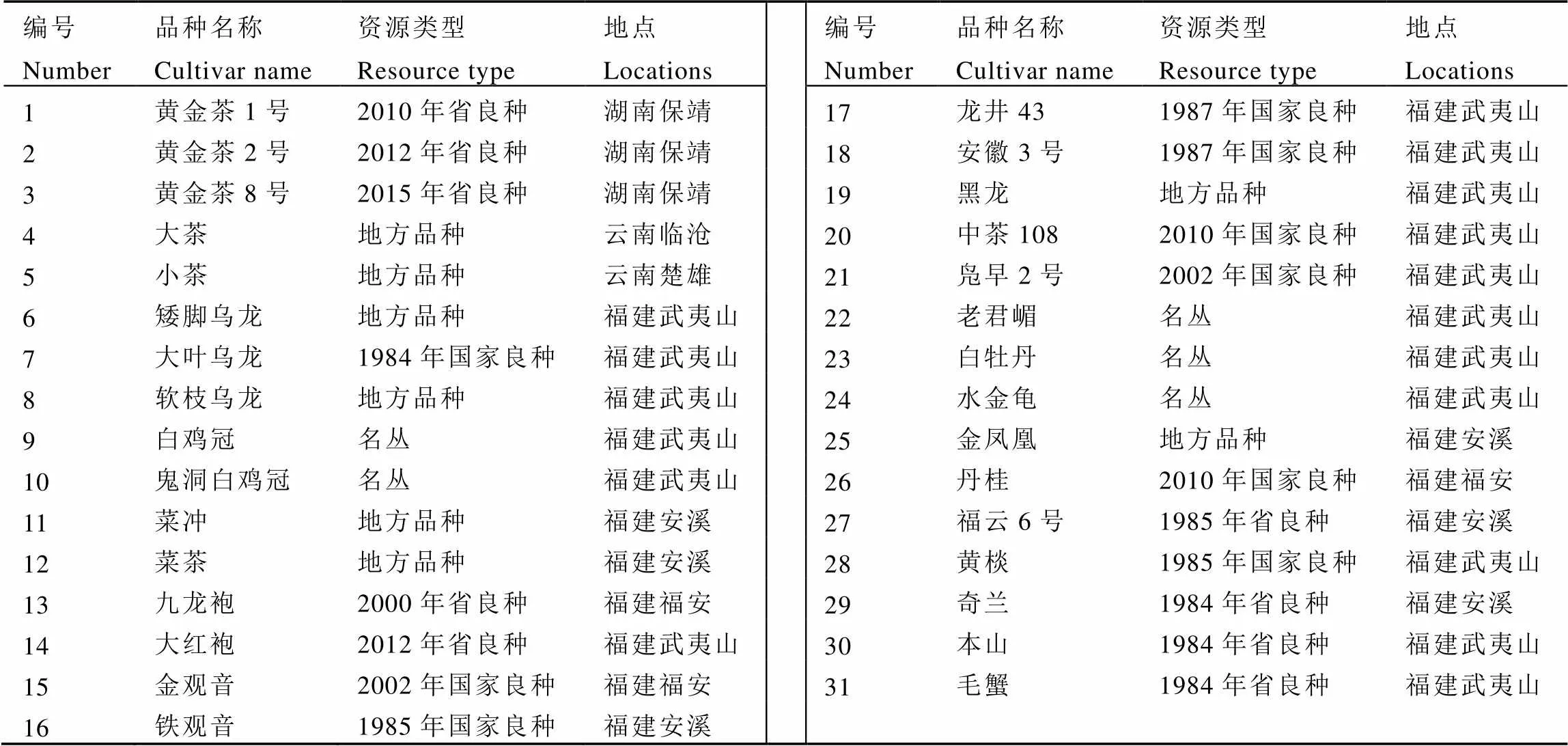
表1 31个茶树品种相关信息
1.2.3 茶籽含油率和脂肪酸组成分析
用离子火焰检测器数据进行脂肪酸的定量分析,通过与内标的峰面积进行比对计算出各个脂肪酸的含量,公式如下:
脂肪酸甲酯质量/µg=(脂肪酸甲酯峰面积÷内标峰面积)×75。
茶籽含油率计算公式:
茶籽含油率=(脂肪酸甲酯总质量÷种子干重)×100%。
油脂的脂肪酸组成按照摩尔百分数来进行计算。根据各个脂肪酸甲酯的质量及其分子量计算出各个脂肪酸的摩尔数,从而得到样品中脂肪酸甲酯的总摩尔数。各个脂肪酸的摩尔数占样品总摩尔数的百分比即为该脂肪酸的摩尔百分数。
1.2.4 茶籽中脂肪酸去饱和酶活力分析
为探讨脂肪酸去饱和酶对茶籽不饱和脂肪酸形成的影响,脂肪酸去饱和酶2(Fatty acid desaturase 2,FAD2)的活力用油酸与亚油酸的比值表示,脂肪酸去饱和酶3(Fatty acid desaturase 3,FAD3)的活力用亚油酸与亚麻酸的比值表示。
1.3 数据分析

2 结果与分析
2.1 茶籽油中脂肪酸总体特征
茶籽经硫酸-甲醇法直接衍生化,利用GC-MS分析得到茶籽油脂的典型质谱图(图1)。经与美国标准研究所的化合物谱图数据库进行比对鉴定出茶籽中的脂肪酸种类。排除了内标十七烷酸(C17∶0)后,从茶籽油脂中检测到10种脂肪酸,包括棕榈酸(C16∶0)、棕榈油酸(C16∶1Δ9)、顺-11-十六碳烯酸(C16∶1Δ11)、硬脂酸(C18∶0)、油酸(C18∶1Δ9)、顺-11-十八碳烯酸(C18∶1Δ11)、亚油酸(C18∶2)、亚麻酸(C18∶3)、二十烷酸(C20∶0)、二十烯酸(C20∶1)。根据谱图峰高可以看出,棕榈油酸、顺-11-十六碳烯酸、二十烷酸和二十烯酸含量较少,茶籽油中主要的脂肪酸是棕榈酸、硬脂酸、油酸、亚油酸和亚麻酸。在当前分析条件下,油酸和顺-11-十八碳烯酸流出峰部分重叠,以油酸为主。饱和脂肪酸包括棕榈酸、硬脂酸和二十烷酸,含量为棕榈酸>硬脂酸>二十烷酸;不饱和脂肪酸包括棕榈油酸、顺-11-十六碳烯酸、油酸、顺-11-十八碳烯酸、亚油酸、亚麻酸和二十烯酸。根据脂肪酸中不饱和键数量,不饱和脂肪酸可分为单不饱和脂肪酸和多不饱和脂肪酸。茶籽中的单不饱和脂肪酸包括棕榈油酸、顺-11-十六碳烯酸、油酸、顺-11-十八碳烯酸和二十烯酸,以油酸为主;多不饱和脂肪酸主要包括亚油酸和亚麻酸,亚油酸的含量高于亚麻酸。
2.2 不同茶树种质间茶籽含油率比较
茶籽含油率以单位茶籽种仁干重所含脂肪酸甲酯的质量表示。31个茶树种质茶籽平均含油率为(31.8±0.9)%,与大茶、小茶、菜茶、老君嵋、水金龟茶籽间的含油率存在显著差异(<0.05)。含油率前5位的种质分别为奇兰(44.7%)、白牡丹(37.9%)、矮脚乌龙(36.4%)、菜茶(36.3%)和凫早2号(35.5%);含油率靠后的5个种质分别为老君嵋(28.2%)、安徽3号(27.8%)、水金龟(25.9%)、大茶(20.2%)和小茶(19.1%)(图2-A)。饱和脂肪酸在这31个种质中的含量为3.7%~8.9%,其中奇兰茶籽的饱和脂肪酸含量最高,比最低的大茶高5.2%(图2-B)。不饱和脂肪酸含量在31个种质中含量为15.0%~35.8%,其中奇兰茶籽的不饱和脂肪酸含量最高,比含量最低的小茶高20.8%(图2-C)。单不饱和脂肪酸是不饱和脂肪酸的主体,在31个种质中含量为11.3%~23.7%;多不饱和脂肪酸含量为3.7%~12.1%,小茶的单不饱和脂肪酸含量最低,比含量最高的奇兰低12.4%,其多不饱和脂肪酸含量比奇兰低8.3%(图2-D、图2-E)。由此可见,小茶和奇兰茶籽含油率具有一定的特殊性。
2.3 不同茶树种质茶籽油脂的脂肪酸组成特征
2.3.1 脂肪酸主要成分相对含量及种质间差异
将每个茶树种质茶籽油脂中检测到的脂肪酸换算成摩尔百分数来表示其相对丰度,结果如图3所示。所有茶树种质茶籽中仅油酸、亚油酸和棕榈酸含量超过10%,平均值分别是51.0%、25.5%和18.3%。油酸是各茶树种质茶籽中的主要脂肪酸成分,占脂肪酸总量的42.1%~59.2%。和其他油料相比,茶籽中油酸含量低于山核桃油(59%~75%)、油菜籽油(61%)、橄榄油(80%)和油茶油(78%~80%)[10-11]。31个茶树中黄金茶2号茶籽的油酸含量最高,达59.2%;水金龟茶籽的油酸含量最低,为42%(图3-A)。
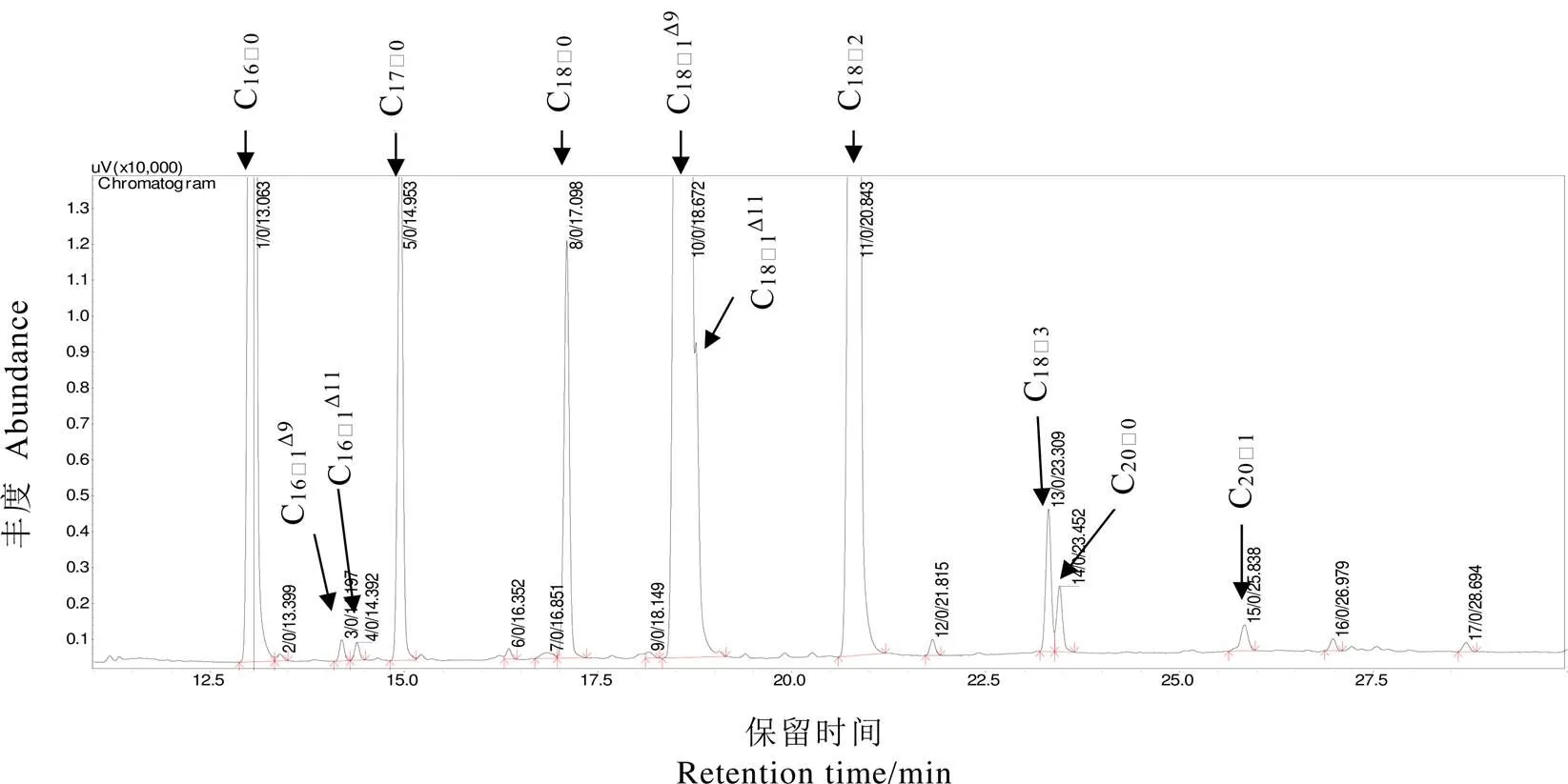
图1 茶籽油脂的总离子色谱图

注:*表示该种质茶籽含油率与均值存在显著差异(<0.05)
Note: * indicates the significant difference of the total oil content between tea plant germplasm and their means (<0.05)
图2 31个茶树种质茶籽含油率
Fig.2 The seed oil contents of the 31 tea germplasms
不同茶树种质茶籽中亚油酸含量仅次于油酸,占脂肪酸总量的18.9%~32.8%。黄棪的亚油酸含量最低,仅为18.9%;而水金龟茶籽的亚油酸含量最高,达32.8%。茶籽油脂中的亚油酸含量低于向日葵油(65%~75%)、玉米油(57%)和大豆油(52.2%),与花生油(31%)和油菜籽油(19%~26%)相近,但高于橄榄油(7.8%)和油茶油(8.3%)[10]。各个种质间棕榈酸含量变化幅度为15.9%~20.8%,小茶的棕榈酸含量最低,为15.9%,较含量最高的鬼洞白鸡冠低4.9%。不同茶树种质茶籽亚麻酸占脂肪酸总量的0.57%~1.0%,其中金凤凰茶籽的亚麻酸含量最低,为0.57%,比含量最高的菜冲茶籽低0.43%。
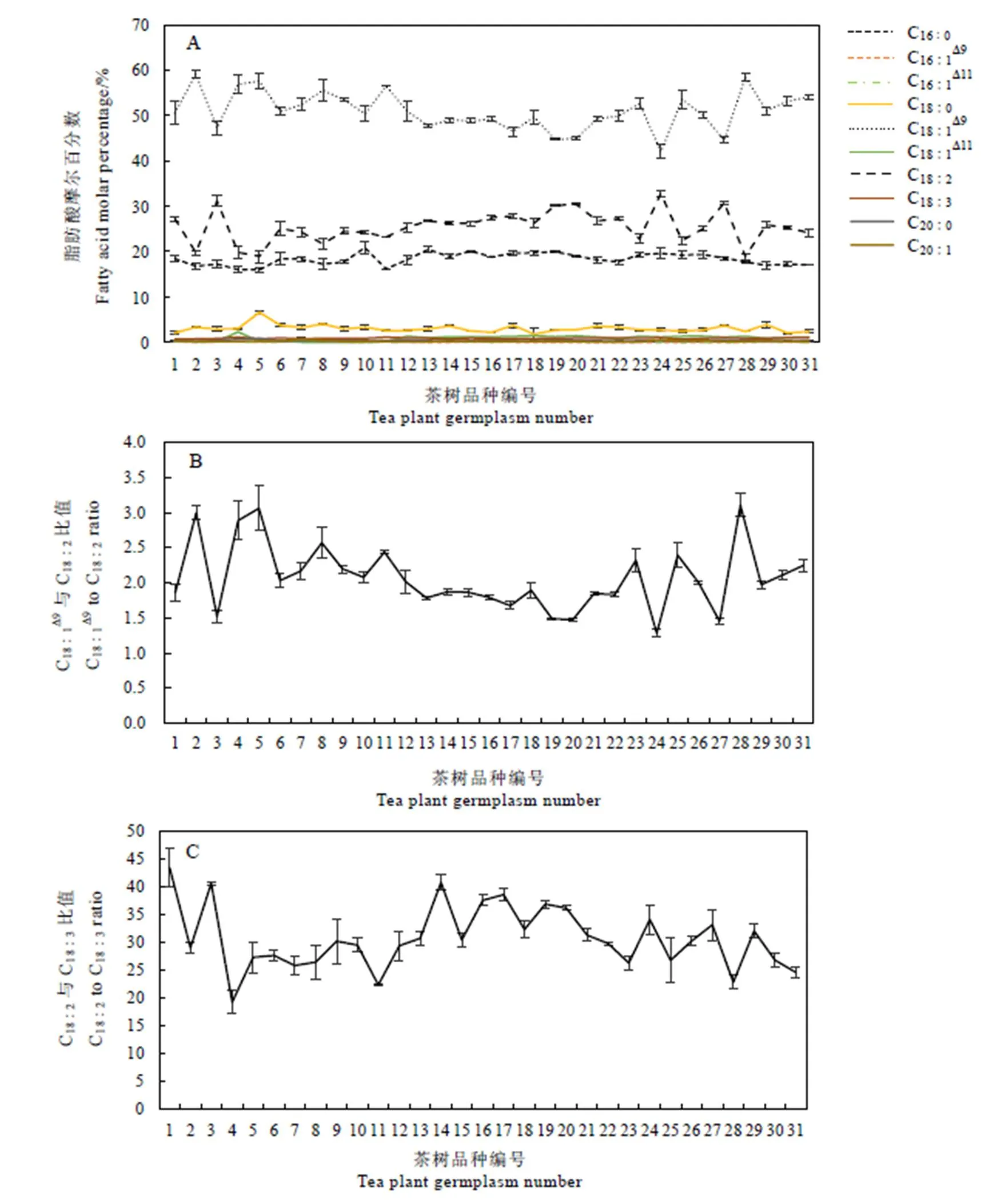
图3 31个茶树种质茶籽油脂脂肪酸组成
2.3.2 油酸、亚油酸和亚麻酸组成特征与FAD2和FAD3活力分析
油酸和亚油酸相对含量在各个种质内呈现出相反的变化趋势,种质中油酸含量高则亚油酸低,反之亦然(图3-A)。这种现象说明控制油酸向亚油酸的转化是决定二者在茶籽中含量的关键步骤。
31个种质茶籽中油酸与亚油酸相对含量的比值为1.28~3.10(图3-B),其中,水金龟最低,黄棪最高。油酸向亚油酸的转化是由微粒体的FAD2蛋白催化完成[12]。油酸与亚油酸的比值可以反映FAD2的相对酶活力,比值越大,FAD2活力越低。油酸与亚油酸的比值排在前5位的是黄棪(3.1)、小茶(3.0)、黄金茶2号(3.0)、大茶(2.9)和软枝乌龙(2.6),表明这5个种质中FAD2酶活力较低。
不同种质茶籽中亚油酸与亚麻酸含量的比值为19.2~43.2。在种子发育过程中亚油酸向亚麻酸的转化由微粒体的FAD3催化完成[13]。亚油酸与亚麻酸的比值高低反映了茶树种质中FAD3的相对酶活力,比值越小则FAD3酶活力越高。31个茶树种质中大茶的亚油酸与亚麻酸含量比值最低,为19.2;而黄金茶1号的比值最高,为43.2。表明大茶的FAD3酶活力最高,黄金茶1号的FAD3酶活力最低(图3-C)。
综上所述,黄棪茶籽的亚油酸相对含量低于其他种质,这与FAD2、FAD3的酶活力有关。
2.3.3 饱和脂肪酸与不饱和脂肪酸分析
虽然供试的31个茶树种质茶籽油脂绝对含量变化幅度较大(图2),但其中饱和脂肪酸占比仅在18.9%~24.4%。菜冲茶籽中饱和脂肪酸占比最低,为18.9%;鬼洞白鸡冠的饱和脂肪酸占比最高,达24.4%;二者的不饱和脂肪酸占比呈现相反的规律(图4-A),这导致不饱和脂肪酸与饱和脂肪酸的比值在3.1~4.3变动(图4-B)。31个茶树种质茶籽油脂中单不饱和脂肪酸的占比为43.6%~59.9%,其中水金龟的单不饱和脂肪酸占比最低,为43.6%;黄棪的单不饱和脂肪酸占比最高,达59.9%。31个茶树种质中多不饱和脂肪酸的占比为19.6%~33.8%,小茶的多不饱和脂肪酸占比最低,为19.6%;而水金龟的多不饱和脂肪酸占比最高,达33.8%(图5-A)。单不饱和脂肪酸与多不饱和脂肪酸的比值为1.3~3.0,水金龟的最低,为1.3;黄棪的最高,达3.0(图5-B)。
2.3.4 18C与16C脂肪酸比值分析
茶籽油脂中的16C脂肪酸包括棕榈酸(C16∶0)、棕榈油酸(C16∶1Δ9)、顺-11-十六碳烯酸(C16∶1Δ11);而18C脂肪酸包括硬脂酸(C18∶0)、油酸(C18∶1Δ9)、顺-11-十八碳烯酸(C18∶1Δ11)、亚油酸(C18∶2)、亚麻酸(C18∶3)。根据样品中各个脂肪酸的摩尔百分数可以计算出该样品中16C和18C脂肪酸的摩尔百分数,从而可以得出18C脂肪酸对16C脂肪酸的比值(图6)。该比值大小近似反映了16C脂肪酸向18C脂肪酸的转化能力,比值越大,说明更多16C脂肪酸被转化成18C脂肪酸。18C与16C脂肪酸比值排前5位的种质分别为小茶、菜冲、大茶、黄金茶2号和奇兰,这说明在地方小种中存在一些有价值的变异。
3 讨论
3.1 茶籽油脂的脂肪酸组成和含油率的种质间特征
常玉玺等[1]分析了福建42份茶树种质资源的茶籽油脂脂肪酸相对含量;常亚丽等[7]分析了豫南地区42份茶树种质茶籽的含油率和脂肪酸组成。本研究分析了26份福建种质以及3份湖南和2份云南的种质,定量方法与以上研究略有差异,但各个脂肪酸相对含量的均值与上述结果较接近。这说明,不同年份茶籽在省内范围和跨省地域间,其油脂均表现出比较稳定的脂肪酸组成。此外,本研究还测定了各个种质的含油率,发现不同种质茶籽含油率存在2倍以上差异(19.1%~44.7%),奇兰的含油率高达44.7%,接近印度种茶籽的含油率[14],高于豫南种质中含油率最高的中黄1号(38.39%)[7]。油脂含量高的前5位的种质依次为奇兰、白牡丹、矮脚乌龙、菜茶和凫早2号,这些种质在叶籽两用茶树育种中可以优先考虑作为杂交亲本,以进一步提高后代茶籽的含油率。目前,茶树种质间含油率存在较大差异的原因尚待进一步研究。一般认为小叶种茶树茶籽含油率高于大叶种[3],而本研究所用的31个种质的叶型分类尚不清楚,且叶的大小受环境条件影响较大,因此还无法证实是否存在此种相关性。另外,结实率对种子含油率的影响尚不清楚,一般乔木茶树种群的结实率较低。茶籽中主要成分为脂肪、淀粉和蛋白,其中淀粉含量仅次于脂肪,占20%左右;蛋白的含量占10%以上[15]。因此,茶树种质间茶籽含油率的差异很可能与茶籽发育过程中光合产物向这3种物质分配的能力有关。了解茶籽发育过程中主要贮藏物质的调控机理将有助于设计新的策略来降低光合产物向淀粉和蛋白的分配,从而提高茶籽的含油率。本研究发现的一系列高含油率种质和低含油率种质,将为进一步探索这一问题提供重要的遗传材料。
3.2 茶籽中不饱和脂肪酸含量的调控途径
随着人们生活水平的提高和健康意识的增强,对食用油的品质要求也越来越高。亚油酸和亚麻酸人体不能合成,必须从饮食中获取,是公认的必需脂肪酸。亚油酸是主要的-6多不饱和脂肪酸,而亚麻酸(-linolenic acid,ALA)是主要的-3多不饱和脂肪酸。此外,二十碳五烯酸(Eicosapentaenoic acid,EPA)和二十二碳六烯酸(Docosahexaenoic acid,DHA)也属于人体需要的-3脂肪酸。常玉玺等[1]在迎春、悦茗香等5个种质中检测DHA含量在0.1%左右,由于其含量低且检测困难,本研究忽略不计。此外,茶籽油中含有0.1%左右的花生酸,是人体合成前列腺素和白三烯的前体,人体可利用亚油酸合成花生酸,因此茶籽油中的微量花生酸不能提供足够的营养价值。虽然人体能够利用亚麻酸合成EPA和DHA,但合成能力非常有限,无法完全满足机体需求而必须从食物中摄取,EPA和DHA的推荐摄取量为0.65 g·d-1[16]。这些长链多不饱和脂肪酸参与大脑发育和功能维持、视觉、免疫和炎症反应,以及合成激素样分子等。-3脂肪酸有助于减少炎症,而大多数-6脂肪酸易促进炎症发生,过高的-6脂肪酸会抑制亚麻酸转化成DHA,因此-6脂肪酸与-3脂肪酸推荐摄取比例为2.3∶1[17]。
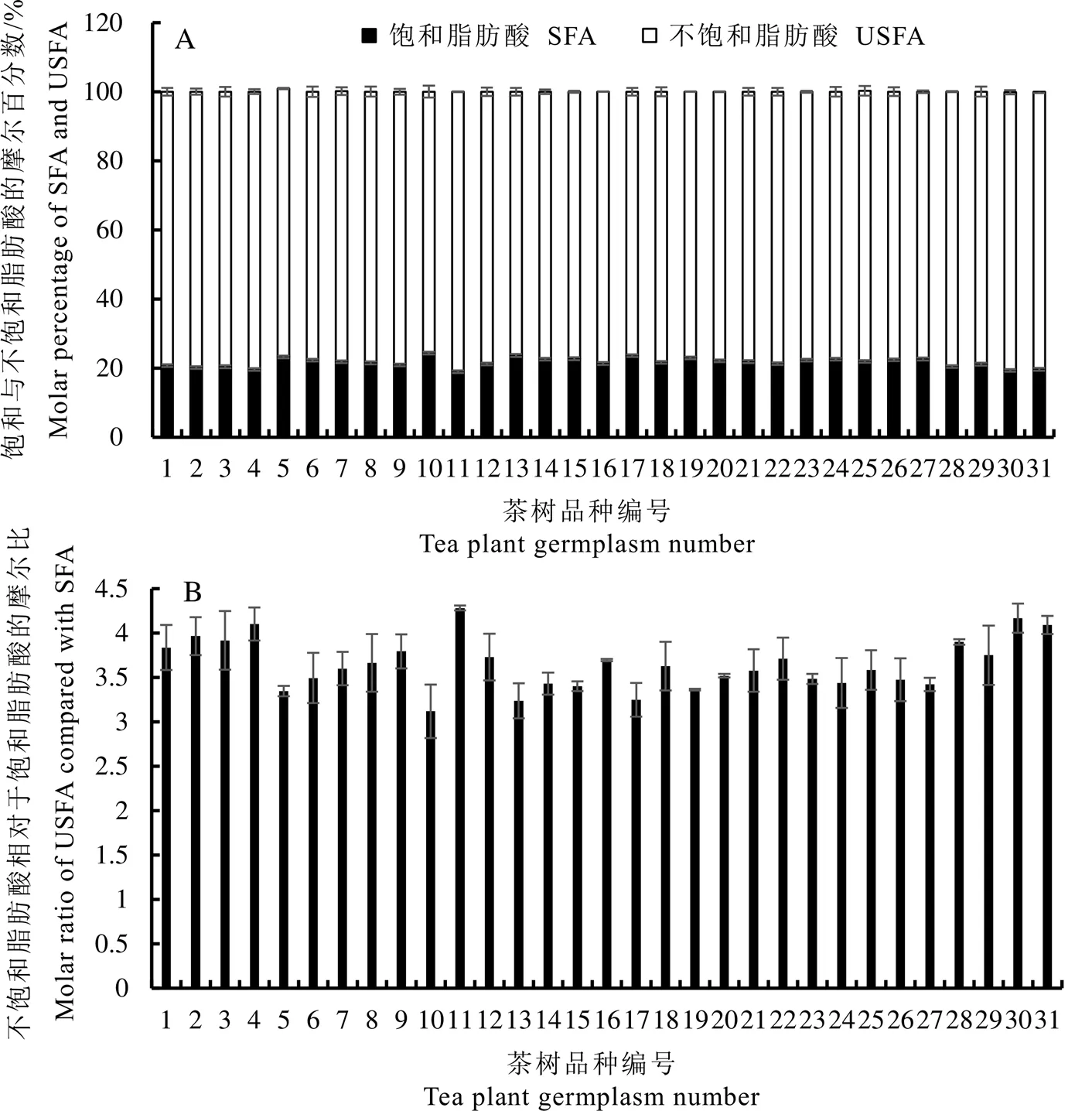
图4 31个茶树种质茶籽油脂饱和脂肪酸与不饱和脂肪酸的组成

图5 31个茶树种质茶籽油脂单不饱和脂肪酸与多不饱和脂肪酸的组成
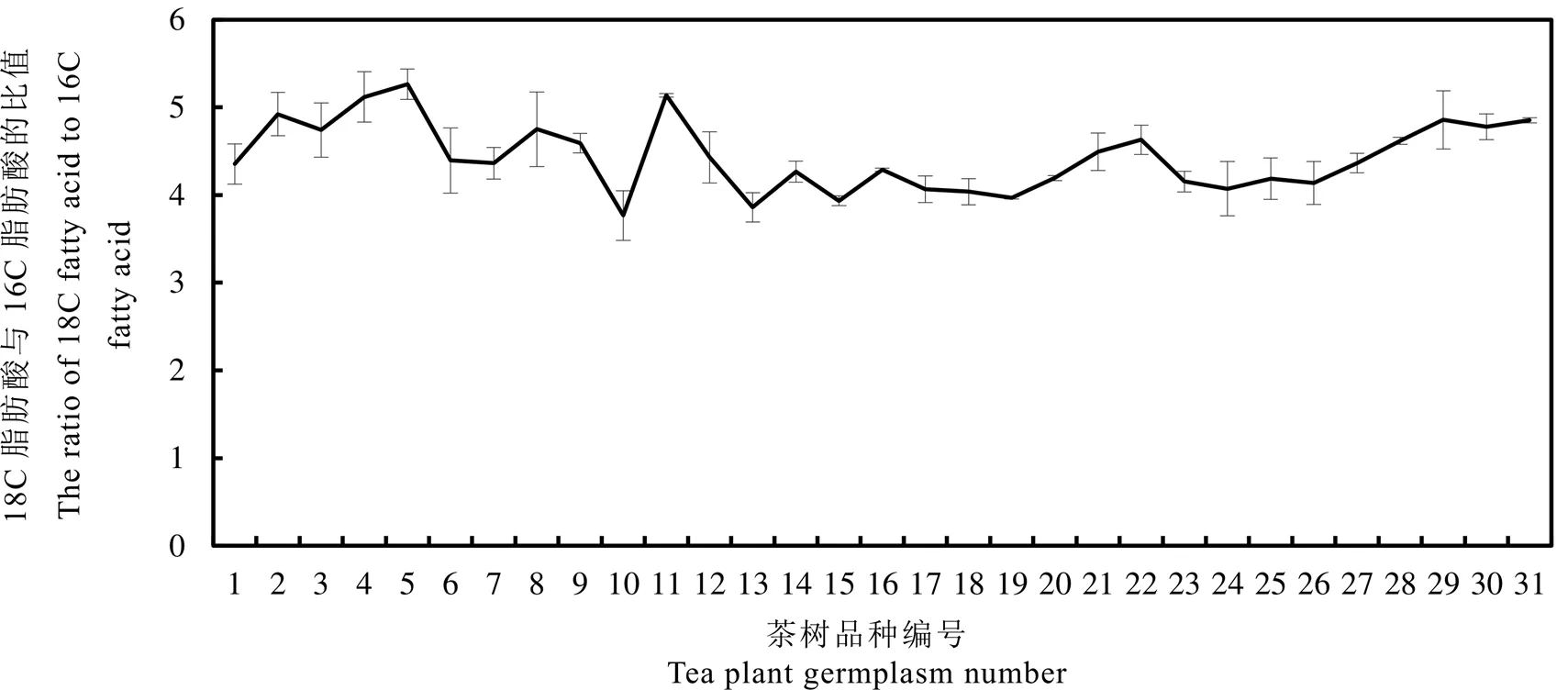
图6 31个茶树种质中茶籽18C脂肪酸与16C脂肪酸的比值
31个种质中亚油酸与亚麻酸的比值高于推荐值8~19倍,从提高茶籽油脂品质的角度看,在将来叶籽两用茶树育种中需要降低茶籽中亚油酸的含量。有两个可能的实现途径:第一,促进亚油酸向亚麻酸的转化。这在降低亚油酸含量的同时提高亚麻酸的含量,从而降低二者比例。因此,在叶籽两用茶树遗传育种中应该优先选用FAD3活力高的种质,FAD3活力排名前5位的分别为大茶、菜冲、黄棪、毛蟹和白牡丹。茶树中有3个FAD3同源基因,分别由TEA014072、TEA017122和TEA000441编码,应进一步确定这3个FAD3同源基因的组织表达特异性,特别是它们在茶籽发育过程中的时空表达模式,从而明确哪些FAD3直接参与茶籽亚油酸和亚麻酸含量调节,进一步开发成分子标记用于茶树育种。第二,降低油酸向亚油酸的转化。选用FAD2活性低的种质可以在降低茶籽亚油酸含量的同时增加油酸的含量。茶树中有2个FAD2同源基因,分别由TEA016089和TEA025470编码。31个种质中FAD2活性低的5个种质为黄棪、小茶、黄金茶2号、大茶和软枝乌龙,可优先考虑作为叶籽两用茶树育种的杂交亲本。
茶籽油脂中饱和脂肪酸含量较高,尤其是棕榈酸,过量食用会增加罹患心血管疾病的风险。降低茶籽油脂棕榈酸的一个可能途径是通过增强棕榈酸向硬脂酸的转化,一旦形成硬脂酸,硬脂酸会在脂去饱和化酶的作用下进一步转化成其他不饱和脂肪酸。因此,选用18C脂肪酸与16C脂肪酸比值高的种质作为杂交亲本更有可能培育出低棕榈酸的后代,候选种质包括小茶、菜冲、大茶、黄金茶2号和奇兰。
选择具有目标性状的种质作为杂交亲本能有效提高杂交育种的成功率,缩短育种周期。本研究对31个种质的油脂性状进行了详细的分析,为将来茶树杂交育种的亲本选择提供有价值的基础信息。单位面积茶园产油量是叶籽两用茶树的一个重要经济指标。除了种仁含油率之外,果实大小、坐果率和出籽率是决定产果量和产油量的关键指标,而这些指标在本研究中尚未涉及,需要进一步调查以最终确定其在叶籽两用茶树育种中的价值。从改善茶籽脂肪酸组成的角度看,本研究发现黄棪同时具有低FAD2和高FAD3活性,是理想的杂交亲本。也可通过多轮杂交的策略,分步将不同种质的优异性状基因整合到一起。
3.3 茶树新梢适制性影响叶果的生长和经济性状的平衡
叶籽两用茶树品种主要利用的器官是叶。每年6月份茶树花芽开始分化,9—12月份开花,完成受精作用后停止生长,翌年4月份果实中脂肪开始累积,6月份后积累速度加快,10月份种子成熟,因此叶和茶果之间对光合产物的竞争很大程度上取决于鲜叶采摘时间。名优绿茶鲜叶采摘大部分集中在4—5月份,此时茶果的营养积累刚刚开始,对叶的产量影响较小。而对于采摘成熟芽叶的茶类如黑茶和乌龙茶,特别是在秋季,叶和茶果之间会产生一定竞争,这也是叶籽两用茶树培育中需要考虑的因素。
3.4 茶籽的综合利用与其组成的关系
茶籽的主要组分有脂肪、淀粉、粗蛋白和茶皂素等[3,10]。种子的油脂含量和蛋白含量一般呈负相关,目前茶籽脂肪含量与茶皂素含量之间的关系还不清楚。本研究重点关注了茶籽的脂肪,下一步需要进一步明确这些组分在不同种质中的分布及其内在关系。在叶籽两用茶树育种中除了油脂外,还需要考虑茶籽的主要终端用途,从而培育出多样化的茶树种质来满足各种下游产业链的需求。
[1] 常玉玺, 郑德勇, 叶乃兴, 等.福建茶树种质资源的茶籽油脂脂肪酸组成分析[J].茶叶科学, 2012, 32(1): 22-28.
Chang Y X, Zheng D Y, Ye N X, et al.Analysis of fatty acid composition in the seed oil from Fujian tea germplasm [J].Journal of Tea Science, 2012, 32(1): 22-28.
[2] 郭元超, 陈荣冰, 肖丽平.茶树不同区系与种群间生殖特性与生殖力的初步探讨(六): 结实力与果实发育类型的观察研究[J].茶叶科学简报, 1983(4): 15-29.
Guo Y C, Chen R B, Xiao L P.Preliminary study on reproductive characteristics and fertility among different floras and populations of tea plants (Ⅵ): study on the relationship between fruit strength and fruit development type [J].Tea Science Bulletin, 1983(4): 15-29.
[3] 叶乃兴, 常玉玺, 郑德勇, 等.茶树果实的特性、功能成分与利用[J].茶叶科学技术, 2011(2): 1-6.
Ye N X, Chang Y X, Zheng D Y, et al.Characteristics, functional components and utilization of tea tree fruits [J].Tea Science and Technology, 2011(2): 1-6.
[4] 马跃青.籽叶两用茶园管理技术的研究[D].合肥: 安徽农业大学, 2012: 15-38.
Ma Y Q.Study on the management technology of tea garden with dual-purpose seed and leaf [D].Hefei: Anhui Agriculture University, 2012: 15-38.
[5] 罗雅慧.茶树良种浙农21叶籽两用栽培技术[J].浙江农业科学, 2014(6): 828-830.
Luo Y H.Cultivation techniques for leaf and seed dual-purpose tea plant Zhenong 21 [J].Zhejiang Agricultural Sciences, 2014(6): 828-830.
[6] Hu J B, Yang G L.Physiochemical characteristics, fatty acid profile and tocopherol composition of the oil fromAbel cultivated in Henan, China [J].Grasas y Aceites, 2018, 69(2): 255.doi: 10.3989/gya.1224172.
[7] 常亚丽, 黄晓兵, 蒋双丰, 等.豫南茶树种质资源籽实脂肪含量及脂肪酸组成分析[J].茶叶科学, 2020, 40(3): 352-362.
Chang Y L, Huang X B, Jiang S F, et al.Analysis of fat content and fatty acid composition of seeds of tea germplasm resources in southern Henan [J].Journal of Tea Science, 2020, 40(3): 352-362.
[8] 官玲亮, 侯凯, 陈郡雯, 等.ω-6和ω-3脂肪酸脱氢酶家族系统进化与功能分化[J].遗传, 2013, 35(5): 643-654.
Guan L L, Hou K, Chen J W, et al.Systematic evolution and functional differentiation of ω-6 and ω-3 fatty acid dehydrogenase families [J].Heredity, 2013, 35(5): 643-654.
[9] Guo L, Du Z H, Wang Z, et al.Location affects fatty acid composition incv Tieguanyin fresh leaves [J].Journal of Food Science and Technology, 2020, 57: 96-101.
[10] 郭华.高档茶籽油的提取及茶籽综合利用技术研究[D].长沙: 湖南农业大学, 2007.
Guo H.Study on extraction of high-grade tea seed oil and comprehensive utilization technology of tea seed [D].Changsha: Hunan Agriculture University, 2007.
[11] 刘莉, 黎锐, 李星儀, 等.四川雅安引进油茶品种含油率及茶油品质分析[J].中国粮油学报, 2019, 34(12): 53-58.
Liu L, Li R, Li X Y, et al.Oil content and tea oil quality analysis of four introducedvarieties in Yaan Sichuan [J].Journal of the Chinese Cereals and Oils Association, 2019, 34(12): 53-58.
[12] Okuley J, Lightner J, Feldmann K, et al.Arabidopsis2 gene encodes the enzyme that is essential for polyunsaturated lipid synthesis [J].The Plant Cell, 1994, 6: 147-158.
[13] Yadav N, Wierzbicki A, Aegerter M, et al.Cloning of higher plant ω-3 fatty acid desaturases [J].Plant Physiol, 1993, 103(2): 467-476.
[14] 夏春华, 朱全芬.茶籽(L)油研究的现状[J].中国茶叶, 1981, 3(4): 63-75.
Xia C H, Zhu Q F.Research status of tea seed (L) oil [J].Chinese Tea, 1981, 3(4): 63-75.
[15] 刘国艳, 王兴国, 金青哲, 等.不同地区茶叶籽油理化指标及脂肪酸组成的比较分析[J].中国油脂, 2013, 38(7): 85-88.Liu G Y, Wang X G, Jin Q Z, et al.Comparative analysis of physical and chemical indexes and fatty acid composition of tea seed oil in different regions [J].China Oil, 2013, 38(7): 85-88.
[16] Kris-Etherton P M, Taylor D S, Yu-Poth S, et al.Polyunsaturatedfatty acids in the food chain in the United States [J].The American Journal of Clinical Nutrition, 2000, 71(sl): 179-188.
[17] Masters C.ω-3 fatty acids and the peroxisome [J].Molecular and Cellular Biochemistry, 1996, 165: 83-93.
Analysis of Characteristics of Lipid Metabolism of Different Germplasm Tea Seeds
CHEN Mingjie1, DU Zhenghua2, QIN Jianheng1, LI Ruoyu1, YU Zhaoxi1, GUO Li2,3*
1.College of Life Sciences,Henan Key Laboratory of Tea Plant Biology, Xinyang Normal University, Xinyang 464000, China; 2.College of Horticulture, Fujian Agriculture and Forestry University, Fuzhou 350002, China; 3.Tea Research Institute, Chinese Academy of Agricultural Sciences, Hangzhou 310008, China
The development of dual-purpose tea resource is especially important for the structural adjustment and efficiency improvement of tea industry.Thus, it is essential to comprehensively characterize the seed-related traits from current available tea germplasms, which can be used to guide dual-purpose tea genetic improvement.In this study, tea seeds of the same year were collected from 31 germplasms grown in Fujian, Yunnan, and Hunan, seed oil contents and compositions were analyzed.We found that there were more than two-fold variations in seed oil content.Ten different fatty acids were detected from tea seed oil, including palmitic acid, palmitoleic acid, 11()-hexadecenoic acid, stearic acid, oleic acid, 11()-octadecenoic acid, linoleic acid, linolenic acid, arachidic acid, and eicosanoid acid.Oleic acid was the dominant component of tea seed oil, which accounted for 42.1%-59.2% of the total fatty acids, followed by linoleic acid (18.9%-32.8%).Saturated fatty acid accounted for 15.9%-20.8% of the total fatty acids, with palmitic acid as the dominant component.Feature analysis of fatty acid composition indicates that there were large variations of the FAD2 and FAD3 activities among these germplasms.Based on these findings, the methods to select parent lines for dual-purpose tea breeding were discussed.
tea seeds, oil content, fatty acid, fatty acid desaturase
S571;S326
A
1000-369X(2021)03-350-11
2021-01-18
2021-03-07
国家自然科学基金(31870803)
陈明杰,男,博士,主要从事植物脂类代谢和脂去饱和化酶研究,mjchen@xynu.edu.cn。*通信作者:fl0206@126.com
(责任编辑:黄晨)
