风沙采煤沉陷区地表生物结皮土壤碳排放对水热因子变化的响应
2021-06-18党晓宏蒙仲举魏亚娟刘禹辛
党晓宏,刘 阳,蒙仲举,高 永,魏亚娟,翟 波,刘禹辛
(1.内蒙古农业大学 沙漠治理学院,内蒙古 呼和浩特 010018; 2.内蒙古杭锦荒漠生态系统国家定位观测研究站,内蒙古 鄂尔多斯 017400; 3.内蒙古自治区水利科学研究院,内蒙古 呼和浩特 010020; 4.中央与地方共建高校特色优势学科“风沙物理”重点实验室,内蒙古 呼和浩特 010018)
我国煤炭资源开采具有空间上的不均衡性,主要集中在西北干旱区、半干旱区和西南山区[1]。据统计,截止到2005年,我国采煤沉陷区面积已达700 km2,且以每年约200 km2的速度增长。预计未来,我国采煤沉陷区面积将达到6.0×104km2[2]。由于我国煤炭资源主要采用井工开采工艺[3],导致原有的生态平衡被打破,形成大面积的采煤沉陷区,其地质、水分和土壤环境均发生了极大程度改变。例如:土地沙化、土壤结构破坏、生物多样性降低。这些变化改变了原有土体结构的水肥运移规律,从而改变了采煤沉陷区生态系统碳循环过程[4]。
生物结皮是由细菌、真菌、地衣、藻类和苔藓等隐花植物分泌的多糖物质与表层土壤颗粒相互作用形成具有生命活性的复合体[5],广泛分布于干旱、半干旱区[6-7],成为荒漠生态系统的重要组成部分。研究表明,生物结皮能提高土壤结构稳定性,增强土壤抗风蚀能力,具有改善土壤水文环境,增强土壤碳氮储蓄能力[8]。CASTILLO-MONROY等[9]研究发现,地衣结皮覆盖区是利比亚半岛土壤碳的主要释放源;王爱国等[10]发现藻类结皮和苔藓结皮土壤CO2通量较去除生物结皮后呈现下降趋势,同时土壤CO2通量的降低程度与生物结皮的组成和生物量呈正相关。齐玉春等[11]研究表明,古尔班通古特沙漠混生结皮在降水后,土壤碳排放速率显著高于裸地。管超等[12]研究发现,增温能抑制生物结皮碳排放。可见,生物结皮土壤碳排放强度与土壤水分和温度密切相关。然而由于荒漠生态系统缺少水分和养分,导致生物结皮在碳源(汇)方面存在较多不确定性。
近年来,关于生物结皮对土壤碳循环的研究较多,而对风沙采煤沉陷区生物结皮土壤碳排放的研究鲜有报道。因此,为了准确掌握采煤沉陷区生物结皮类型与土壤碳排放间的关系,对采煤沉陷区不同结皮类型土壤碳排放进行实地测定。李家塔煤矿沉陷区位于黄土高原与毛乌素沙地接壤的晋陕蒙交界区,生态环境异常脆弱。经过多年的矿区生态综合治理,矿区生态环境得到极大地改善,林分内分布大面积的生物结皮,其种类相对丰富,具有一定代表性[13]。基于此,笔者以毛乌素沙地采煤沉陷区为研究区域,以该地区典型生态修复树种小叶杨林和沙柳林内的生物结皮为研究对象,对各林分类型下生物结皮的土壤碳排放日动态、土壤水热条件进行实地原位动态监测,通过建立回归方程明确沙质土壤水热条件对不同林分类型下生物结皮土壤碳排放特征的影响,为采煤沉陷区生态修复中区域碳汇/源的评价提供借鉴。
1 研究区域与研究方法
1.1 研究区概况
研究区位于毛乌素沙地东北缘,内蒙古自治区鄂尔多斯市伊金霍洛旗上湾镇李家塔矿区(110.0°E~110.4°E,39.2°N~39.5°N)。气候类型为中温带干旱、半干旱大陆性季风气候,具有冬季漫长严寒,多风沙,夏季炎热,蒸发强烈,昼夜温差大,季节性温差悬殊,无霜期短等气候特点。年均降水量350 mm左右。地貌类型主要为波状沙地和黄土丘陵沟壑地貌。地质条件复杂,在基岩上覆有黄土、沙土、风积沙等,且地表层面土壤多与粉煤灰结合形成夹层现象。研究区地表常年分布有大面积生物结皮,由于水分条件、生物种类和发育阶段的不同,生物结皮颜色各异,主要呈绿色、黑色、黑褐色和黄绿色等。研究区生物结皮主要利用春季融雪、降雨、凝结水进行生长,并在干旱缺水季节和气温低于0 ℃时进入休眠状态。矿区优势种为小叶杨(Populussimonii)、沙柳(Salixpsammophila)、零星分布杨柴(Hedysarummongolicum)、柠条锦鸡儿(Caraganakorshinskii)、油蒿(Artemisiaordosica)、虫实(Corispermumhyssopifolium)、克氏针茅(Stipakrylovii)、羊草(Leymuschinensis)、百里香(ThymusmongolicusRonn)等[14-15]。
1.2 研究方法
1.2.1样地选择和布设
本试验于2019年4月中旬(春季)进行,选择小叶杨林地、沙柳林地和恢复裸地为试验样地。试验样地选择地势相对平坦、微地形相对一致的区域。在各样地分别设置3个20 m×20 m的样方,3个样地间相距不足20 m,利用5点法在每个大样方内设置5个2.0 m×2.0 m的小样方作为监测点。调查每个样方内的结皮类型(图1)、结皮厚度、结皮盖度等。具体情况见表1。
1.2.2土壤碳排放测定
选择晴朗无风或微风天气进行测定。利用ACE土壤碳通量自动监测系统(品牌:英国ADC;型号:ACE)对不同生物结皮类型土壤呼吸速率进行同步测定,每次测量时长为30 min。开始测定前12 h,用枝剪去除圈内地上草本植物并清除枯落物,利用取样器(高8.0 cm,直径34.5 cm的钢圈)垂直压入土层5.0 cm,并保证生物结皮的完整性。每种结皮覆盖样地制作5个取样点即为5次重复。样方设置在植物冠幅边缘。将土壤碳通量自动监测仪自带的水分和温度探头插入5.0 cm的土壤层中,同步测定土壤的温度和含水量。为了保证研究结果的代表性,观测时间7:00~18:00,观测频次为1次/h。连续观测7 d,分别记录生物结皮土壤碳排放速率、地表5.0 cm深度处土壤温度、地表温度和土壤含水量。
土壤CO2排放速率与土壤温度、土壤含水量关系分别采用线性模型、指数模型、对数模型、多项式函数模型进行拟合,最后通过赤池系统(Akaike information criterion,AIC)准则和决定系数R2筛选出最优拟合方程,最后通过极大似然值判定拟合方程的优劣。
Rs=a+bT+cW
(1)
Rs=a+bT+cW+dTW
(2)
Rs=aTbWc
(3)
AIC=2lnL+2P
(4)
式中,Rs为土壤碳排放速率;a,b,c和d为拟合参数;T为土壤温度;W为土壤含水量;L为回归方程的极大近似然函数;P为回归方程的独立参数个数;AIC值越小说明拟合方程越优。
1.2.3数据处理
用Excel 2007对监测土壤碳排放数据进行整理,剔除异常数据。采用SPSS 22.0软件进行统计分析,采用单因素方差分析(one-way ANOVA)对3类土壤碳排放CO2排放进行差异显著性分析,同时采用Eviews 10软件对荒漠地区土壤碳排放速率与土壤温度和含水量进行相关分析。利用Origin Pro 2018作图。
2 结果与分析
2.1 生物结皮土壤碳排放速率及环境因子日动态变化规律
由图2可知,3类结皮碳排放和环境因子日动态变化存在一定的差异性。3类结皮土壤碳排放速率日均值由大到小依次为藻类结皮(0.47 μmol/(m2·s))>藓类结皮(0.45 μmol/(m2·s))>地衣结皮(0.44 μmol/(m2·s))。土壤碳排放速率的日变化均呈现出“单峰”曲线特征,其中藻类结皮的土壤碳排放速率“峰值”出现在12:00,藓类结皮的土壤碳排放速率“峰值”出现在12:30,地衣结皮的土壤碳排放速率“峰值”出现在13:00。土壤碳排放日变幅由大到小依次为藓类结皮地、藻类结皮和地衣结皮,其值分别0.10~0.83,0.05~0.77和0.12~0.77 μmol/(m2·s)。3类结皮下5.0 cm深度土壤温度和地表温度呈现出相同趋势,整体表现为藻类结皮>地衣结皮>藓类结皮。3类结皮的土壤含水量出现最低值时间各异,藓类结皮在13:00达到土壤含水量的“谷值”,藻类结皮在14:00达到土壤含水量的“谷值”,地衣结皮在17:30达到土壤含水量的“谷值”,其时间最迟。
2.2 土壤碳排放速率与主要环境因子的关系
2.2.1土壤碳排放速率与土壤温度的关系
由图3可知,土壤碳排放速率呈先升高后降低的变化趋势。3类结皮土壤碳排放速率最大值时间相差约30 min。藻类结皮12:00达到土壤碳排放峰值0.77 μmol/(m2·s),其次为藓类结皮在12:30达到峰值0.83 μmol/(m2·s),最后为地衣结皮在13:00出现峰值0.77 μmol/(m2·s)。通过回归拟合发现3类结皮土壤碳排放速率与土壤温度的拟合关系均为二次函数关系;土壤碳排放速率与0~5 cm表层土壤温度呈顺时针环状分布,其中以藻类结皮最为明显。对3类结皮土壤碳排放速率和0~5 cm土壤温度进行分段拟合发现,相同土壤温度情况下,上升阶段显著大于下降阶段(p<0.01),导致这种现象的原因是土壤碳排放速率都呈现先升后降的趋势导致的时间滞后效应。

图3 采煤沉陷区3种类型结皮土壤碳排放速率与土壤温度回归拟合Fig.3 Regression fitting of soil carbon emission rate and soil temperature three typical biologically-crusted soils in coal mining subsidence area
由表2可知,由于生物结皮土壤碳排放速率对温度的响应不同,3类结皮的土壤碳排放速率与0~5 cm土壤温度均呈现二次函数关系,其决定系数分别为0.59,0.46和0.64(p<0.01)。

表2 土壤碳排放速率与土壤温度的回归方程Table 2 Regression equation of soil carbon emission rate and soil temperature
2.2.2土壤碳排放速率与表层土壤含水量的关系
由图4可知,3类结皮表层土壤含水量变幅较小。其中,藓类结皮为0.17~0.30 μmol/(m2·s)、藻类结皮为0.17~0.27 μmol/(m2·s)、地衣结皮为0.19~0.32 μmol/(m2·s)。
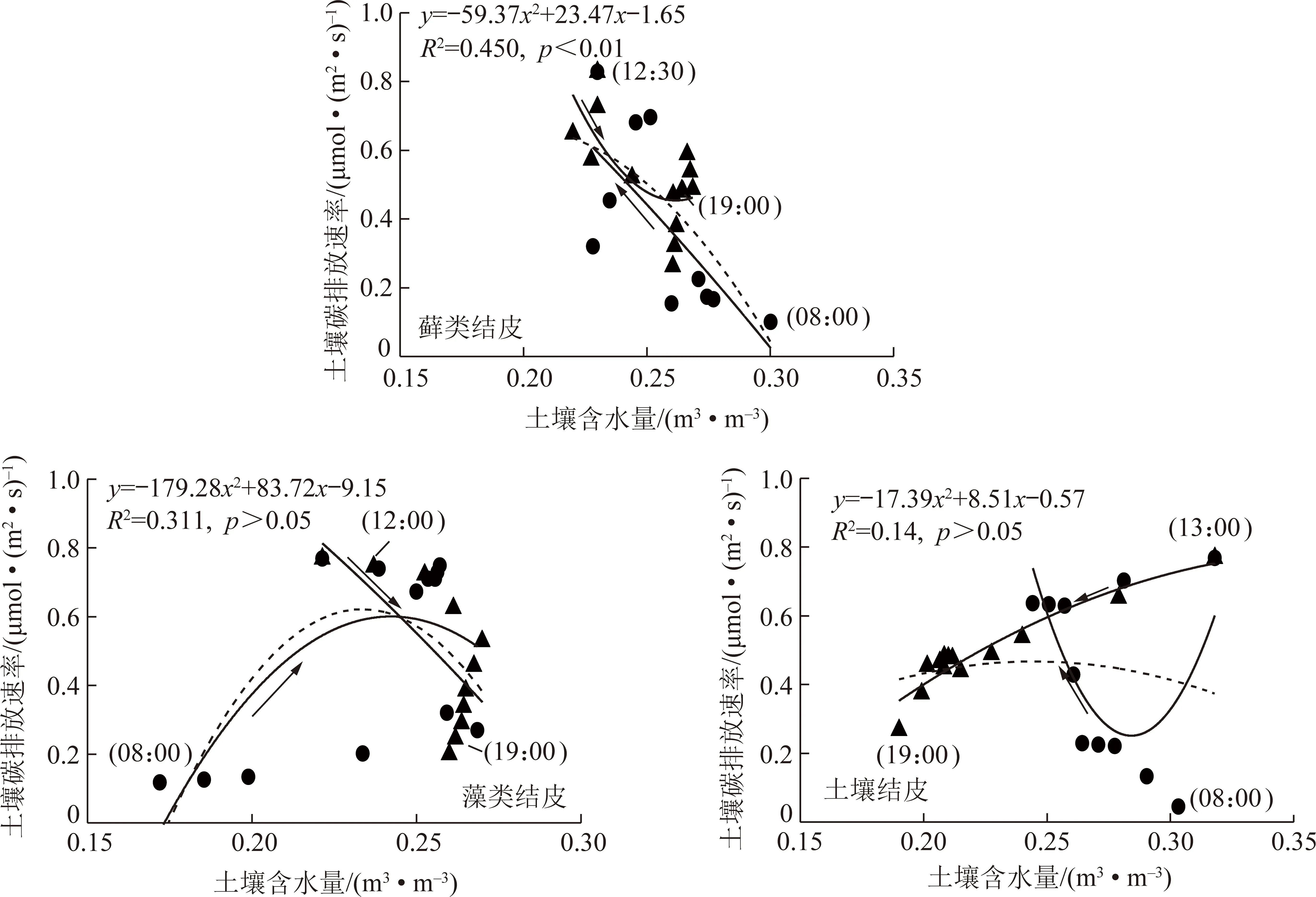
图4 采煤沉陷区3种类型生物结皮土壤碳排放速率与土壤含水量回归拟合Fig.4 Regression fitting of soil carbon emission rate and soil moisture in three forest stands in coal mining subsidence area
通过拟合回归发现,3类结皮土壤碳排放速率与土壤含水量最优关系均为二次函数。其中藓类结皮土壤含水量决定系数R2为0.45,且达到极显著水平(p<0.01)。藻类结皮和地衣结皮土壤碳排放速率与土壤含水量决定系数R2分别为0.14和0.13,但均未达到显著水平(p>0.05)。
由表3可知,对土壤碳排放速率日峰值为界限将土壤碳排放速率与土壤含水量进行分段拟合发现,3类结皮土壤碳排放速率与土壤含水量最优拟合关系均呈二次函数。其中藓类结皮土壤碳排放速率与表层土壤含水量呈逆时针环形分布,而藻类结皮和地衣结皮土壤碳排放速率与表层土壤含水量呈顺时针分布。
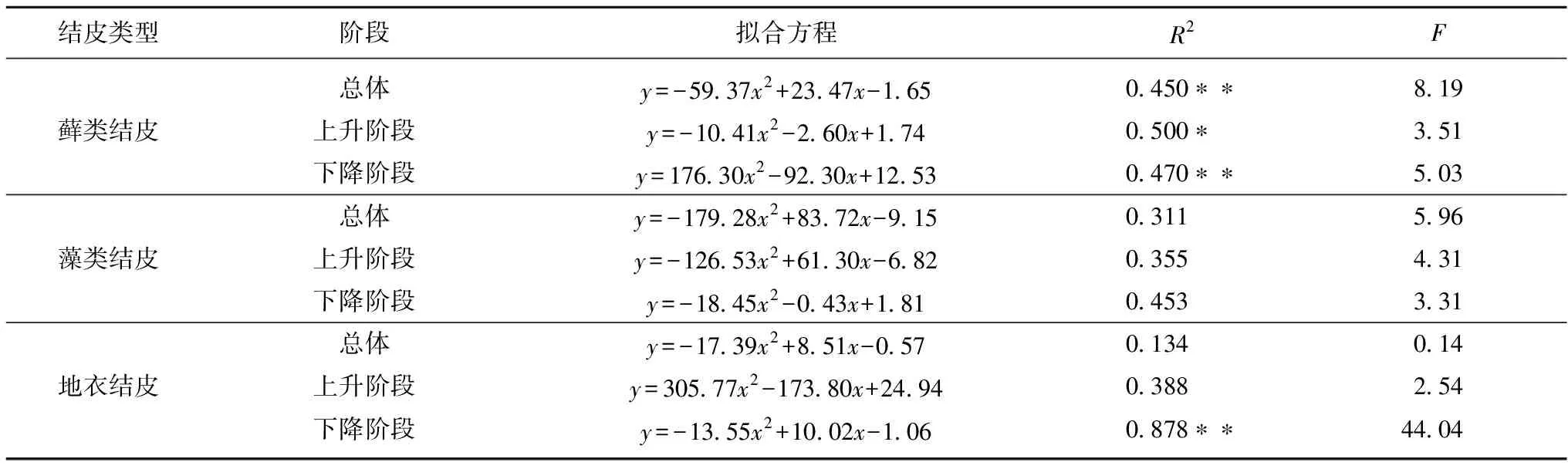
表3 土壤碳排放速率与土壤含水量的回归方程Table 3 Regression equation of soil carbon emission rate and soil moisture
2.2.3土壤碳排放速率与土壤表层温度、含水量的关系
由表4可知,采煤沉陷区3种类型结皮土壤碳排放速率与表层土壤温度和土壤含水量的协同关系均达到极显著水平(p<0.01)。3类结皮土壤碳排放速率与表层土壤温度和土壤含水量拟合方程均表现为方程(1)变异解释率最低,方程(2),(3)拟合效果较好,其中方程(3)的拟合效果最好,方程(1)在土壤碳排放速率对土壤温度和湿度协同响应研究中适用性最差。采煤沉陷区3种类型土壤含水量和土壤温度可以解释其土壤碳排放速率的57.8%~82.5%。通过分析AIC发现,藓类结皮在土壤碳排放速率对土壤温度和湿度协同响应研究中适用性强于方程(2),(3),可以解释其58.3%的土壤碳排放情况,但藻类结皮和地衣结皮,拟合方程(2),(3)可以较好的解释土壤碳排放速率,解释系数在57.8%以上。

表4 土壤碳排放速率与表层土壤温度和含水量的回归拟合关系Table 4 Regression fitting relationship between soil carbon emission rate and surface soil temperature and water content
3 讨 论
土壤碳排放速率是一个复杂的生化过程,其排放强度受生物因素和非生物因素共同影响[16]。本研究显示3类生物结皮土壤碳排放日动态变化各异。土壤碳排放速率日均值由大到小依次为藻类结皮(0.47 μmol/(m2·s))>藓类结皮(0.45 μmol/(m2·s))>地衣结皮(0.44 μmol/(m2·s)),说明结皮类型是导致碳排放强度的主要因素[17]。该研究结果与胡宜强等[18]对沙坡头生物结皮的研究结果一致。表明随着生物结皮的演替,生物结皮的土壤呼吸速率呈递增趋势且均高于裸地。生物结皮的形成增强了土壤碳排放速率[18],这可能是由土壤温度导致的。藻类结皮温度(13.27 ℃)高于藓类结皮(12.50 ℃)和地衣结皮(12.97 ℃)(表5),导致土壤酶活性升高,加速了土壤有机质的分解,进而导致土壤微生物呼吸加速[19],从而增强了土壤碳排放速率[20]。但是,与毛乌素沙地非采煤沉陷区相比(土壤碳排放速率为0.63 μmol/(m2·s))[21],采煤沉陷区各类生物结皮碳排放速率较小。因为采煤沉陷区土壤有机质含量较低,导致碳排放的减弱。加之春季气温较低,土壤干燥,导致生物结皮生理活动几乎处于休眠状态,因此导致采煤沉陷区生物结皮碳排放低于裸地碳释放[22]。说明生物结皮的覆盖,降低土壤碳呼吸速率。
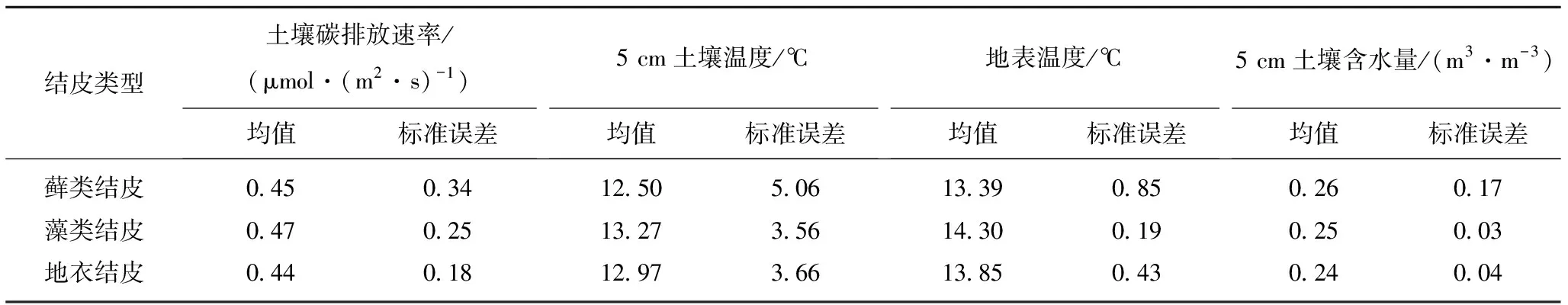
表5 生物结皮土壤碳排放速率及温湿度比较Table 5 Comparison of soil carbon emission rate,temperature and humidity of biological crust
本研究显示藓类结皮、藻类结皮和地衣结皮土壤碳排放日变化速率分别为0.17~0.30,0.17~0.27和0.19~0.32 μmol/(m2·s)。可见,藻类生物结皮土壤碳排放日变幅小于藓类结皮和地衣结皮。该研究结果显著低于管超等[19]对腾格里沙漠生物结皮的研究。这主要与土壤温湿度有关。研究表明,土壤呼吸速率与土壤表层温湿度呈显著正相关[23]。本研究中,监测时间为春季,土壤温度和含水量分别在3.87~18.46 ℃,0.17~0.32 m3/m3,远远小于其他季节土壤温湿度。土壤温湿度较低,抑制土壤酶的活性,降低了土壤有机质的分解,从而抑制土壤微生物呼吸。本研究显示土壤碳排放速率日动态均成单峰曲线特征,其中藓类结皮和地衣结皮的土壤碳排放速率“峰值”出现在12:30,藻类结皮的土壤碳排放速率“峰值”出现在14:00。与赵东阳[24]对黄土高原土壤藓结皮的研究结论一致。该研究结果表明,砂土和砂质壤土藓类结皮碳呼吸峰值出现在14:00。3类结皮土壤含水量均表现为先降低后升高的趋势,即为清晨和傍晚含水量较高,土壤含水量随着温度的升高而逐渐降低。土壤碳排放是酶促作用的结果,随着一天当中昼夜的变化,土壤温度和土壤水分等环境因子也会发生改变,这使得酶促反应中的各类酶活性不同,进而导致酶促反应的异质性,3类生物结皮土壤碳排放产生日动态间的差异[25]。研究中还发现,5 cm土壤温湿度日变化曲线峰值出现时间滞后于3类土壤碳排放速率,且土壤碳排放速率与土壤温度和水分均呈显著正相关关系。但是导致土壤温湿度滞后性的原因有待于进一步研究。
温度对土壤碳排放速率的影响局限在土壤表层[26]。土壤温度和土壤含水量对于碳排放有较大影响,并呈显著线性关系。土壤碳排放是一个复杂的生物地球化学循环过程,是环境、植物、土壤共同作用下的过程[27-29]。也有多数研究发现,温度是影响土壤CO2排放的重要因素[30-31]。研究中发现,土壤碳排放速率日动态变化与0~5 cm土壤温度变化趋势基本吻合,均在早晨8:00以后,随着土壤温度升高土壤碳排放速率骤然升高,午后土壤温度降低,土壤碳排放速率随之降低,但生物结皮土壤碳排放速率“峰值”都早于土壤温度“峰值”,2者时间上存在着滞后现象[32-35]。相关研究发现,造成土壤碳排放与土壤温度时间上的分离的主要原因是温度的混合效应。同时,生物因素也是影响土壤碳排放和土壤温度间滞后关系的重要环节,包括植物的光合作用和根系生长、枯落物以及微生物的动态变化的共同影响[36]。土壤温度影响了土壤的酶活性。在低温环境下,土壤酶的活性受到限制,随着温度的增加活性增强,当超过最适温度后,酶活性急剧下降,甚至降解。由于根系呼吸和土壤微生物呼吸都需要酶的参与,因此土壤温度会影响土壤碳排放速率[37]。整体而言,土壤碳排放速率时间变化对表层土壤温度的响应均达到了极显著水平,这与赵东阳[24],辜晨[38]研究结果一致。本研究中发现,李家塔采煤沉陷区3类结皮土壤含水量较低且变化幅度较小。水分对土壤碳排放作用相对较小,可能是结皮覆被区的微生物,能够利用少量的水分刺激其生理活性[39]。只有藓类结皮的土壤含水量和土壤碳排放速率之间存在一致的动态变化,2者存在显著的负相关关系。土壤含水量对于土壤碳排放的影响主要集中在对植物和微生物的能量供应和对其体内的二次分配作用,与此同时,对土壤通透性和气体扩散进行调控[40]。
4 结 论
(1)毛乌素沙地北缘采煤沉陷区3类结皮碳排放速率的日变化特征曲线基本一致。总体上呈现“不对称钟形”的“单峰”曲线特征,土壤碳排放速率峰值出现在12:00~13:00。土壤碳排放速率日均值由大到小依次为藻类结皮(0.47 μmol/(m2·s))>藓类结皮(0.45 μmol/(m2·s))>地衣结皮(0.44 μmol/(m2·s)),说明随着生物结皮演替,其土壤碳排放速率逐渐增强。
(2)3类结皮土壤碳排放速率与表层土壤温度和土壤含水量均呈二次函数关系。土壤含水量与土壤温度可以较好的解释土壤碳排放速率,拟合方程中2者对于土壤碳排放速率的解释系数在57.8%以上,说明土壤温度和土壤含水量能显著影响土壤碳排放速率。
(3)毛乌素沙地采煤沉陷区生物结皮覆盖可有效抑制土壤碳排放。在风沙采煤沉陷区生态修复过程中建议多建植小叶杨和沙柳等乡土树种。
