海南三亚滨海雨林、半落叶季雨林到落叶季雨林的植物群落多样性及种间联结研究
2021-06-10吕安琪李东海杨小波吴律欣
吕安琪 李东海 杨小波 吴律欣


摘 要:为探索三亚沿海地区(自东向西)从滨海雨林过渡到半落叶季雨林、落叶季雨林的物种多样性及种间关系的变化趋势。该文采用植物群落学调查方法对三种植被类型进行群落调查,进一步使用物种丰富度指数(dGL)、Shannon-Wiener指数(H)、Simpson指数(P)和Pielou均匀度指数(E)来表征各植被类型的物种多样性水平,并使用联结系数(AC)、共同出现百分率(PC)及Spearman秩相关系数检验三种方法來表征各植被类型的种间关系。结果表明:(1)从滨海雨林过渡到半落叶季雨林再到落叶季雨林的生态序列变化过程中,森林优势物种组成逐渐由喜湿树种过渡为耐旱性较强的树种。(2)生物多样性的变化与降雨量呈正相关,均呈现自东向西逐渐递减。(3)三种植被类型共有物种的种间关系分析显示,三种植被类型种间关系松散,滨海雨林的种间联结强度相对较高,半落叶季雨林与落叶季雨林相似。综上结果认为,从滨海雨林到落叶季雨林物种间的协同共存关系在减弱,排斥竞争关系在增强;无论是物种组成、物种多样性水平还是种间关系,半落叶季雨林与落叶季雨林更相似。
关键词: 三亚滨海丘陵, 物种多样性, 联结系数, 相关系数, 年降水量
中图分类号:Q948
文献标识码:A
文章编号:1000-3142(2021)03-0384-12
收稿日期:2019-10-08
基金项目:国家自然科学基金(31760170) [Supported by the National Natural Science Foundation of China(31760170)]。
作者简介: 吕安琪(1994-),硕士研究生,主要从事植物生态学研究,(E-mail)1250938850@qq.com。
通信作者:杨小波,博士,教授,研究方向为植物生态学与植物资源学,(E-mail)yanfengxb@163.com。
Plant community diversity and inter-specific relationship of coastal rain forest,semi-deciduous monsoon forest to deciduous monsoon forest in coastal hills of
Sanya City, Hainan Province
L Anqi1, LI Donghai2, YANG Xiaobo2*, WU Lüxin1
( 1. School of Life and Pharmaceutical Sciences, Hainan University, Haikou 570228, China; 2. School of Ecology and Environment, Hainan University, Haikou 570228, China )
Abstract:To explore the changing trend of species diversity and interspecific relationship of the forests in coastal hills of Sanya City (from east to west) from coastal rain forest to semi-deciduous monsoon forest and deciduous monsoon forest, the phytocoenological investigation methods were used to investigate those three types of vegetation communities in our field works. The species diversity in different types of vegetation were analyzed by the species richness index(dGL), Shannon-Wiener index(H), Simpson index(P) and Pielou evenness index(E). The interspecific relationships in different types of vegetations were analyzed by association coefficient(AC), co-occurrence percentage(PC) and Spearmans rank correlation coefficient test. The results were as follows: (1) Dominant species gradually changed from hygrophilous species to drought-tolerant species during the process of ecological sequence change from coastal rain forest to semi-deciduous monsoon forest and then to monsoon deciduous forest; (2) The change trend of species diversity was positively related to the annual precipitation, which decreased gradually from east to west; (3) Interspecific relationship of the three vegetation types was relatively loose; while the intensity of interspecific association in coastal rain forest was relatively high, and showed likeness between the semi-deciduous monsoon forest and deciduous monsoon forest. Those indicated that the co-existence of species was weakened from coastal rain forest to deciduous monsoon forest while the repelling competition was strengthened. In conclusion, the species composition, diversity and interspecific associations of semi-deciduous monsoon forest were more similar to those of deciduous monsoon forest.
Key words: coastal hills of Sanya City, plant species diversity, association coefficient, correlation coefficient, annual precipitation
植物生长发育所需的水分主要来自降水,降水量的不同会诱导植物改变对水分的利用方式,进一步影响到物种的分布格局,使得一定区域内的物种多样性水平及各物种间的关系发生变化(Mcguire & Treseder,2010;刘菊红等,2019)。物种多样性与种间联结是植物生态学研究的重要组成内容,也是植物群落重要的指示参数,其研究不仅有助于了解群落的物种组成、丰富度水平、物种变化程度及各物种间的相互作用,还有助于揭示群落组成结构特点、群落动态及演替趋势等(陈玉凯等,2011)。目前已有大量的研究表明影响物种多样性的因素很多,包括气候因子、地形因子、土壤因子和生物因子等(刘冠成等,2018)。武建双等(2012)探讨了藏北高寒草地物种多样性沿降水梯度的分布格局,结果表明该研究区的物种丰富度、多样性及均匀度均受降水条件的显著影响并与降水量呈指数增长,降水对群落结构和地上生物量还起到促进作用;Irl et al.(2015)通过探究气候、降水等因素对拉帕尔马地区物种丰富度的影响,结果表明年降水量显著影响该区域的物种丰富度。当然,群落的种间联结性也受多种因素的影响,如环境因素、生物自身因素等,而生物自身因素又包括了竞争、化感作用等(徐满厚等,2016)。刘菊红等(2019)研究了荒漠草原主要植物种间关系对降水年型变化的响应,其结果表明年降水量的下降使得种对间的关系由共存为主转变为竞争为主。因此,降水量的不同会影响群落的物种多样性及种间关系,研究不同降水条件下的植物群落的多样性及种间关系,对不同植被类型植物群落的管理及保护具有重要的生态意义与价值。
热带地区高温多雨,而海南三亚降水干湿季分明,滨海丘陵地区年降水量沿水平经度梯度自东向西呈现逐渐减少的自然变化趋势,植被类型也表现出由滨海雨林过渡到季雨林的变化特点,其季雨林自东向西又可分为半落叶季雨林与落叶季雨林。海南岛东侧滨海丘陵地区植物种类多样且结构相对稳定,目前对该区域内植物多样性的研究较少(陈红锋等,2005 ;车秀芬等,2006;黄运峰,2009),在多样性基础上进一步探索群落内种间关系的研究更少(杨琦等,2014;杨青青等,2017)。基于此,本研究在海南三亚滨海丘陵地区三种植被类型(滨海雨林、半落叶季雨林及落叶季雨林)分别设置了乔灌木群落样方,详细调查了群落内的物种组成与群落结构,以进一步阐释,在水分梯度变化条件下物种多样性水平及种间关系从滨海雨林过渡到半落叶季雨林及落叶季雨林的過程中的变化规律。
1 研究区域及样地概况
三亚市位于海南岛南端(18°09′34″—18°37′27″ N、108°56′30″—109°48′28″ E),陆地总面积为1 919.58 km2,东西长91.6 km,丘陵面积约占25.2%,地势由北向南逐渐倾斜。由于地处海南岛半干旱气候区,光照资源丰富且受海洋性气候,降水量分为明显干湿季。由于人类活动的影响,目前在三亚市的沿海地区仅在部分丘陵山包上,仍然保存有自然森林植被,即东部滨海雨林(简称为滨海雨林),主要在红霞岭和六道岭综合生态自然保护区有自然森林的分布,年降水量1 600~1 800 mm,年均温25.0 ℃;中部半落叶季雨林(简称为半落叶季雨林),主要在火岭猕猴自然保护区和鹿回头岭有自然森林的分布,年降水量1 200~1 371 mm,年均温25.4 ℃;以及西部落叶季雨林(简称为落叶季雨林),主要在南山岭有自然森林的分布,年降水量1 000~1 100 mm,年均温25.0 ℃(气候数据来自世界气候数据网站http://www.worldclim.org/)(Hijmans et al.,2005),三个地区年降水量具有明显差异。
2 研究方法
2.1 样地设置与调查
依照《植物群落学实验手册》中植物群落的分析方法,对植物群落数量特征采用标准的样方法进行。由于相对典型的热带雨林、季雨林或滨海森林的植物群落组成要简单一些,植物群落最小面积也较小,一般调查面积在2 800 m2左右。朱华等(1998)在讨论云南热带雨林物种多样性的取样面积时认为最适样方面积为2 500 m2;陈红锋等(2005)、车秀芬等(2006)研究海南岛滨海雨林植物群落结构时样方调查面积分别为2 600、2 800 m2;刘万德等(2009)、苏红华等(2018)研究海南岛季雨林植物群落时样方调查面积均近3 000 m2。结合前人对海南岛滨海雨林及季雨林的调查经验,再根据三亚滨海丘陵植被分布现状,样方设置情况如下:在东部红霞岭(109°38′55.00″—109°39′45.00″ E、18°14′30.40″—18°14′42.60″ N,海拔121~179 m)及六道岭(109°35′12.30″—109°36′15.10″ E、18°11′45.40″—18°13′57.00″ N, 海拔59~384 m)的滨海雨林共设置了30个10 m×10 m样方,累计调查面积3 000 m2;在中西部火岭(109°30′8.70″—109°30′27.50″ E、18°12′32.50″—18°13′8.00″ N,海拔21~230 m)、鹿回头岭(109°28′30.00″—109°29′40.00″ E、18°11′43.20″—18°11′48.20″ N,海拔72~244 m)及南山岭(109°10′30.40″—109°11′37.40″ E、18°17′30.30″—18°18′30.60″ N,海拔188~456 m)的季雨林共设置了36个10 m × 10 m样方,累计调查面积3 600 m2(在中部自然保护区内取样2 800 m2,在西部略受旅游活动影响的区域内补点800 m2)。每个样方进行植物群落学调查,对样方内每个株高≥1.5 m的树木进行每木调查,记录种名、胸径、树高、枝下高和冠幅大小等指标。由于植被分布在沿海低丘陵区,受强风影响大,森林中乔灌木整体偏矮。东部滨海雨林森林高度多在2~7 m,密度约为1.366 株·m-2,中部半落叶季雨林森林高度多在2~6 m,密度约为1.611株·m-2),西部落叶季雨林森林高度多在2~6 m,密度约为1.473 株·m-2。
根据各样方数据计算每个种群的重要值(方精云等,2009)。重要值(IV)=(相对密度+相对频度+相对显著度)/3。
采用Jaccard相似性系数(Cj)(方精云等,2009)表征各地区物种组成的相似程度。
Cj=ca+b-c。
式中:c为两地共有物种数;a、b分别为两样方的物种数。
2.2 物种多样性计算
植物物种鉴定通过野外采集标本,查阅《海南植物图志》及Flora of China等对样地内植物进行分类鉴定。采用的物种多样性指数(方精云等,2009)包括物种丰富度(dGL)、Shannon-Wiener指数(H)、Simpson指数(P)及Pielou均匀度指数(E)。
dGL=SlnA;
H=-∑Si=1PilnPi;
P=1-∑Si=1Pi2;
E=HlnS。
式中:S为样地内出现的物种数;A为样方面积;Pi为第i种的相对密度。
2.3 种间联结计算
采用联结系数(AC)和共同出现百分率(PC)(张倩媚等,2006)表征联结强度,将16个共有物种在样方内是否出现转化为二元数据矩阵,构建2×2列联表(Dice,1945),并计算a、b、c、d值。
若ad-bc≥0,则AC=ad-bc(a+d)(b+d);
若ad-bc<0且a≤d,则
AC=ad-bc(a+b)(a+c);
若ad-bc<0且a>d,则AC=ad-bc(c+d)(b+d);
PC=aa+b+c。
式中:a为2个物种均出现的样方数;b、c为有且仅有其中1个物种出现的样方数;d为2个物种均不出现的样方数。将AC划分为五个等级,分别为-1≤AC<-0.6;-0.6≤AC<-0.2;-0.2≤AC<0.2;0.2≤AC<0.6;0.6≤AC≤1。AC数值越趋近1,则种对正联结越强;AC数值越趋近-1,则种对负联结越强。将PC划分为四个等级,分别为0%≤PC<20%;20%≤PC<50%;50%≤PC<70%和70%≤PC≤100%。PC数值越趋近100%,则种对正联结越强。
进一步使用Spearman秩相关系数检验(徐满厚等,2014)表征各种群的种间相关性。
r(i,k)=1-6∑Nj=1(xij-xi)2(xkj-xk)2N3-N。
式中:N為样方总数;xij和xkj分别是种i和种k在样方j中的秩;r(i,k)值域是 [-1,1],正值表示正相关,0表示无相关,负值表示负相关。
3 结果与分析
3.1 三种植被类型物种组成比较
滨海雨林样方内株高≥1.5 m的植物有123种,隶属于41科97属;半落叶季雨林样方内株高≥1.5 m的植物有63种,隶属于30科53属;落叶季雨林样方内株高≥1.5 m植物有46种,隶属于21科39属。从科、属、种丰富度来看,滨海雨林最高,半落叶季雨林次之,落叶季雨林最低。三种植被类型群落物种组成的相似性结果为滨海雨林与半落叶季雨林、落叶季雨林植物群落相似性系数分别为
19.86%、21.57%,半落叶季雨林与落叶季雨林相似性系数为26.74%,即半落叶季雨林与落叶季雨林物种组成更相似。重要值分析可以体现出树种在群落中的优势度,各植被类型物种重要值排序见表1(仅列出重要值排名前十的物种),其中三种植被类型共有优势种包括厚皮树(Lannea coromandelica)、刺桑(Streblus ilicifolius)等。滨海雨林优势种还包括银柴(Aporosa dioica)、青梅(Vatica mangachapoi)等;半落叶季雨林优势种还包括海南榄仁(Terminalia hainanensis)、黄花三宝木(Trigonostemon lutescens);落叶季雨林优势种还包括黄花三宝木。可见三种植被类型物种组成既具有相似性又具有差异性。
3.2 三种植被类型物种多样性比较
群落多样性是生物群落的重要特征之一,反映了群落本身的特征及其与环境间的关系。由于样方内物种数受调查面积影响显著,因此以落叶季雨林样地面积为基准,进行物种多样性计算与分析比较。
三种植被类型统计结果见表2,其中滨海雨林丰富度为18.401;半落叶季雨林物种丰富度为9.425;落叶季雨林物种丰富度为6.881。滨海雨林丰富度明显高于半落叶季雨林,半落叶季雨林高于落叶季雨林,即滨海雨林物种丰富度最高。Shannon-Wiener指数(H)可以反映个体密度、群落类型及演替阶段等,并且群落物种分布越均匀、数量越多,则H值越大。Pielou均匀度指数(E)也能体现出群落内物种分布的均匀度程度,E值越高,均匀度越大。从Shannon-Wiener指数(H)和Pielou均匀度指数(E)来看,滨海雨林指数显著高于半落叶季雨林、落叶季雨林,且半落叶季雨林、落叶季雨林数值相近,即滨海雨林植物群落个体数量和树木分布均匀度相较于半落叶季雨林、 落叶季雨林更高。Simpson指数(P)又称为优势度指数,从结果来看,三种植被类型优势度指数都较高,群落优势种较显著。综上所述,滨海雨林植物多样性最高、半落叶季雨林次之,落叶季雨林最低,即物种多样性自东向西呈递减变化趋势。
3.3 三种植被类型植物群落的共有物种
根据植物群落的调查结果,选取三种植被类型共有的16个物种(表3)进行种间联结性与相关性计算,进一步探究群落种间关系的变化。
3.4 共有物种的种间联结比较
滨海雨林16个共有物种AC值见图1。其中,表现为正联结的种对有60对,占总种对数的50.00%;表现为负联结的种对有57对,占总种对数的47.50%;表现为无联结的种对有3对,占总种对数的2.50%,正负关联比为1.05。半落叶季雨林AC值见图2,其中表现为正联结的种对有36对,占总种对数的30.00%;表现为负联结的种对有40对,占总种对数的33.33%; 表现为无联结的种对有44对,占总种对数的36.67%,正负关联比为0.90。落叶季雨林AC值见图3,其中表现为正联结的种对有50对,占总种对数的41.67%;表现为负联结的种对有51对,占总种对数的42.50%;表现为无联结的种对有19对,占总种对数的15.83%,正负关联比为0.98。
随着生态序列的变化,种间联结倾向于正联结。从AC正负关联比来看,滨海雨林比值大于1,而半落叶季雨林、落叶季雨林比值均小于1且数值相近,故滨海雨林正联结种对数多于负联结种对数,种间联结相较更紧密,而半落叶季雨林、落叶季雨林正联结种对数少于负联结种对数,种间联结相较更松散,且半落叶季雨林、落叶季雨林联结强度相似。从联结强度上来看,滨海雨林种间联结达到强联结的种对有49对,占总种对数的40.83%;半落叶季雨林达到强联结的种对有32对,占总种对数的26.67%;落叶季雨林达到强联结的种对有35对,占总种对数的29.17%。这说明滨海雨林达到强联结程度的种对比例明显大于半落叶季雨林、落叶季雨林,同时半落叶季雨林、落叶季雨林强联结种对比例相近,故滨海雨林种间联结性更强,半落叶季雨林、 落叶季雨林种间联结较弱且强度相似。
三种植被类型共同出现百分率(PC)结果见图1、图2、图3。其中,滨海雨林PC>50%的种对有22对,占总种对数的18.33%;半落叶季雨林PC>50%的种对有20对,占总种对数的16.67%;落叶季雨林PC>50%的种对有19对,占总种对数的15.83%。虽然绝大多数种对PC值均较低,但是相比较而言,滨海雨林PC>50%的比例最大,而半落叶季雨林、落叶季雨林PC>50%的比例较小且数值相近,故滨海雨林种间联结性更强,半落叶季雨林、落叶季雨林种间联结较弱且强度相似。
综上所述,三种植被类型AC值与PC值结果具有一致性,即滨海雨林群落种间联结性更强,半落叶季雨林、落叶季雨林种间联结强度均相对较弱且联结水平相似。
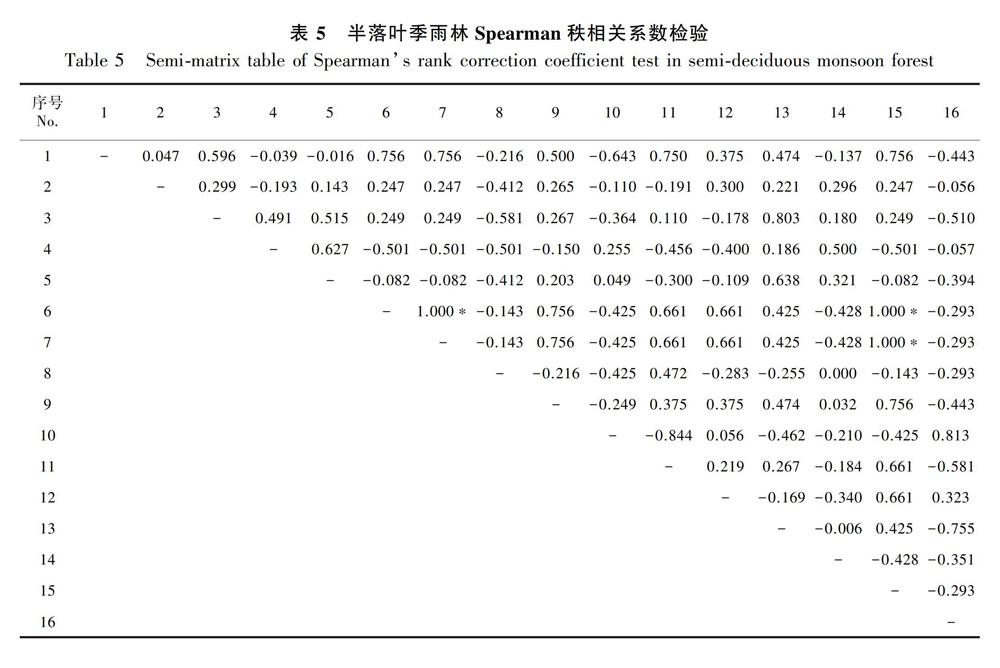
Spearman秩相关系数检验分析可以弥补和完善χ2检验,能更好地反映群落内物种间的关联,且其对物种的分布形式没有要求。滨海雨林Spearman秩相关系数检验结果见表4。其中,表现为正相关的种对有62对,占总种对数的51.67%;表现为无相关的种对有2对,占总种对数的1.66%;表现为负相关的种对有56对,占总种对数的46.67%。正负关联比为1.11。种间相关性达到极显著或显著水平的种对有4对,相关显著率为3.33%。其中,刺桑与青枣核果木、刺桑与崖柿(Diospyros chunii)、青枣核果木与崖柿3对种对为极显著正相关,占总种对数的2.50%;赤才与鹊肾树种对为显著正相关,占总种对数的0.83%,其他种对均未表现出显著的相关性,绝大部分种对相关性较弱。半落叶季雨林Spearman秩相关系数检验结果见表5。表现为正相关的种对有60对,占总种对数的50.00%;表现为无相关的种对有1对,占总种对数的0.83%;表现为负相关的种对有59对,占总种对数的49.17%,正负关联比为1.02。种间相关性达到极显著或显著水平的种对有3对,且龙眼(Dimocarpus longan)与毛柿(Diospyros strigosa)、龙眼与锈毛野桐(Mallotus anomalus)、毛柿与锈毛野桐这3对种对均表现为极显著正相关,相关显著率为2.50%,其他种对均未表现出显著的相关性,绝大部分种对相关性较弱。落叶季雨林Spearman秩相关系数检验结果见表6。从表6可以看出,表现为正相关的种对有61对,占总种对数的50.83%;表现为负相关的种对有59对,占总种对数的49.17%。
正负关联比为1.03。种间相关性达到极显著或显著水平的种对有2对,相关显著率为1.66%。其中,山石榴和细基丸种对为极显著正相关,占总种对数的0.83%;基及树与香合欢种对为显著正相关,占总种对数的0.83%,其他种对均未表现出显著的相关性,绝大部分种对相关性较弱。
从上述Spearman秩相关系数检验结果来看,滨海雨林种对正负关联比高于半落叶季雨林、落叶季雨林,且半落叶季雨林、落叶季雨林比值相近;从种对显著率来看,滨海雨林种对相关性显著率是半落叶季雨林显著率的1.33倍,是落叶季雨林显著率的2倍。因此,滨海雨林种间相关性更强,半落叶季雨林、落叶季雨林种间相关性相对较弱且相关性水平相似,这与三种植被类型的AC值、PC值的结果具有一致性。
4 讨论与结论
水分是植物生长发育重要的限制条件因素,且土壤水分是植物吸收水分的主要来源(贾海坤等,2005),而自然降水可直接影响土壤中的含水量,故常用年降水量来指示水分因子,年降水量显著影响植物多样性(Alvarez-Yepiz et al.,2014)。三种植被类型分别所在的东、中、西区域处于同一纬度带,并都属于滨海丘陵区,海拔波动范围小,温度差异小(王春乙,2014),根据研究地区气候因子数据可知,三个区域气候差异主要表现在年降水量的多少,且在研究区域内年降水量沿水平经度梯度自东向西呈现出逐渐递减的自然变化趋势,那么这种变化对自然植被的植物多样性会产生什么样的影响?
4.1 年降水量与物种组成
本研究区域内,东部年降水量丰富,植被类型为滨海雨林,样方内有植物123种,隶属于41科97属;中、西部年降雨量相对较少,植被类型过渡到半落叶季雨林、落叶季雨林,半落叶季雨林样方内有植物63种,隶属于30科53属;落叶季雨林样方内有植物46种,隶属于21科39属。从科、属、种丰富度来看,滨海雨林群落中科、属、种丰富度明顯高于半落叶季雨林、落叶季雨林。在热带地区,树木落叶最主要和直接的因素是水分匮乏(Condit et al.,2010)。海南岛热带季雨林植物群落适应旱季水分匮乏的其中一个重要特征为具刺植物种类及个体数量多(刘万德,2009)。从三种植被类型共有优势种如厚皮树(落叶树种)和刺桑(具刺)的种群密度来看,滨海雨林样方内厚皮树种群密度为0.005株·m-2,刺桑种群密度为0.053株·m-2,而半落叶季雨林样方内这两个种群密度分别为0.046株·m-2和0.173株·m-2,在落叶季雨林样方内这两个种群密度分别为0.073、0.233株·m-2,反映了物种对环境水分差异的适应性。从其他优势种来看,滨海雨林优势种包括银柴和青梅等,青梅是热带雨林典型代表树种之一,喜湿(胡玉佳,1986),且仅在滨海雨林发现有该种群的分布,体现了其喜湿的生物学特性及其对水分环境的喜好性。而半落叶季雨林、落叶季雨林森林主要由较耐旱的热带常绿和落叶阔叶树种组成,优势种包括海南榄仁、黄花三宝木、基及树等,均属于耐旱性强的树种(黄运峰,2009),也体现了其对水分匮乏环境的适应性。同时对比三种植被类型优势种组成,可发现半落叶季雨林与落叶季雨林较滨海雨林与落叶季雨林优势种相似度更高,同时物种相似性系数也表明半落叶季雨林与落叶季雨林群落物种相似性更高。这与刘万德等(2009)研究海南岛霸王岭两种典型热带季雨林群落特征结果具有一定的相似性。由此可见,三种植被类型物种组成上既具有相似性又具有差异性,在从滨海雨林过渡到半落叶季雨林再到落叶季雨林的生态序列变化过程中,森林优势物种组成逐渐由喜湿树种过渡为耐旱性较强的树种。
CONDIT R, WATTS K, BOHLMAN SA, 2010. Quantifying the deciduousness of tropical forest canopies under varying climates [J]. J Veget Sci, 11(5): 649-658.
DICE LR, 1945. Measure of the amount of ecological association between species [J]. Ecology, 26: 297-302.
FANG JY, WANG XP, SHEN ZH, et al., 2009. Methods and protocols for plant community inventory [J]. Biodivers Sci, 17(6): 533-548. [方精云, 王襄平, 沈泽昊, 等, 2009. 植物群落清查的主要内容、方法和技术规范 [J]. 生物多样性, 17(6):533-548.]
GAO Y, DU F, WANG YN, et al., 2016. Interspecific competition of six succession series species with different soil moisture treatments in loess hilly region [J]. Res Soil Water conserv, 23(6):192-197. [高艳, 杜峰, 王雁南, 等, 2002. 土壤水分对黄土丘陵区演替序列种种间竞争的影响 [J]. 水土保持研究, 23(6):192-197.]
HIJMANS RJ, CAMERON SE, PARRA JL, et al., 2005. Very high resolution interpolated climate surfaces for global land areas [J]. Int J Climatol, 25: 1965-1978.
HU YJ, 1986. Study on the population of Vatica hainanensis in Hainan Island [J]. Chin Bull Bot, (Z1):95-97. [胡玉佳, 1986. 海南岛青梅种群生物学研究简报 [J]. 植物学通报, (Z1):95-97.]
HUANG SN, LI YD, LUO TS, et al., 2000. Dynamic of associations between tree species in a secondary tropical montane rain forest at Jianfengling on Hainan Island [J], Chin J Plant Ecol, (5): 569-574. [黄世能, 李意德, 骆土寿, 等, 2000. 海南岛尖峰岭次生热带山地雨林树种间的联结动态 [J]. 植物生态学报, (5):569-574.]
HUANG YF, 2009. Stydy on forest vegetation types and plant diversity in coastal hilly areas of Sanya City, Hainan Province [D]. Haikou: Hainan University: 26-41. [黄运峰, 2009. 三亚市沿海低丘陵森林植被类型和植物多样性研究 [D]. 海口:海南大学:26-41.]
IRL SDH, HARTER DEV, STEINBAUER MJ, et al., 2015. Climate vs. topography-spatial patterns of plant species diversity and endemism on a high-elevation island [J]. J Ecol, 103: 1621-1633.
JIA HK, LIU YH, XU X, et al., 2005. Simulation of soil water dynamics in a caragana intermedia woodland in Huangfuchuan watershed: relationships among slope, aspect, plant density and soil water content [J]. Chin J Plant Ecol, (6):44-51. [贾海坤, 刘颖慧, 徐霞, 等, 2005. 皇甫川流域柠条林地水分动态模拟——坡度、坡向、植被密度与土壤水分的关系 [J]. 植物生态学报, (6):44-51.]
LIU GC, HUANG YX, WANG QG, et al., 2018. Effects of environmental factors on plant species diversity: research progress [J]. Chin Agric Sci Bull, 34(13): 83-89. [刘冠成, 黄雅曦, 王庆贵, 等, 2018. 环境因子对植物物种多样性的影响研究进展 [J]. 中国农学通报, 34(13):83-89.]
LIU JH, ZHANG J, L SJ, et al., 2019. Response of interspecific relationships among main plant species to the change of precipitation years in desert steppe [J]. Acta Bot Boreal-Occident Sin, 39(7): 1289-1297. [刘菊红, 张军, 吕世杰, 等, 2019. 荒漠草原主要植物種间关系对降水年型变化的响应 [J]. 西北植物学报, 39(7):1289-1297.]
LIU LL, SHENG JD, CHENG JH, et al., 2016. Relationship between plant species characteristics and climate factors in different grassland types of Xinjiang [J]. Acta Pratac Sin, 25(5): 1-12. [刘利利,盛建东, 程军回, 等, 2016. 新疆不同草地类型植物物种特征与水热因子的关系研究 [J]. 草业学报, 25(5):1-12.]
LIU WD, 2009. Community ecology of tropical monsoon forest in Hainan Island, China [D]. Beijing: Chinese Academy of Forestry: 30-37. [刘万德, 2009. 海南岛热带季雨林群落生态学研究 [D]. 北京:中国林业科学研究院:30-37.]
LIU WD, ZANG RG, DING Y, 2009. Community features of two types of typical tropical monsoon forests in Bawangling Nature Reserve, Hainan Island [J]. Acta Ecol Sin, 29(7): 3465-3476. [刘万德, 臧润国, 丁易, 2009. 海南岛霸王岭两种典型热带季雨林群落特征 [J]. 生态学报, 29(7):3465-3476.]
MCGUIRE KL, TRESEDER KK, 2010. Microbial communities and their relevance for ecosystem models: Decomposition as a case study [J]. Soil Biol Biochem, 42(4): 529-535.
SU HH, WANG H, CHEN ZJ, et al., 2018. The characteristics of species composition, structure and diversity of tropical semi-deciduous forest in Jianfengling,Hainan Island [J]. For Environ Sci, 34(6): 21-27. [蘇红华, 王红, 陈宗杰, 等, 2018. 海南尖峰岭热带半落叶季雨林物种组成、结构及其多样性特征 [J]. 林业与环境科学, 34(6):21-27.]
WANG CY, 2014. Hainan climate [M]. Beijing: China Mereorological Press: 18. [王春乙, 2014. 海南气候 [M]. 北京:气象出版社:18.]
WANG P, GOU ZH, NONG SQ, et al., 2018. Species diversity and floristic components of tropical secondary forests in hilly areas of central Hainan [J]. Chin J Trop Crops, 39(4):802-808. [王牌, 苟志辉, 农寿千, 等, 2018. 海南中部丘陵区热带次生林物种多样性及区系分析 [J]. 热带作物学报, 39(4):802-808.]
WU JS, LI XJ, SHEN ZX, et al., 2012. Distribution pattern of species diversity along precipitation gradient in alpine grassland transect of northern Tibet [J]. Acta Pratac Sin, 21(3): 17-25. [武建双, 李晓佳, 沈振西, 等, 2012. 藏北高寒草地样带物种多样性沿降水梯度的分布格局 [J]. 草业学报, 21(3):17-25.]
XU MH, LIU M, ZHAI DT,et al., 2016. A review of contents and methods used to analyze various aspects of plant interspecific associations [J]. Acta Pratac Sin, (24):8224-8233. [徐满厚, 刘敏, 翟大彤, 等, 2016. 植物种间联结研究内容与方法评述 [J]. 生态学报, (24):8224-8233.]
YANG Q, ZHOU J, TAO C, et al., 2014. Comparative study on interspecific association in tropical evergreen monsoon elfin forest at two successional stages in Tongguling of Hainan Island [J]. Chin Agric Sci Bull, 30(22): 8-15. [杨琦, 周婧, 陶楚, 等, 2014. 海南岛铜鼓岭热带常绿季雨矮林2个演替阶段的种间联结性对比研究 [J]. 中国农学通报,30(22):8-15.]
YANG QQ, YANG ZY, CHEN XH, et al., 2017. Interspecific associations of dominant plant populations in secondary forest of Syzygium odoratum in tropical coast [J]. Sli Silv Sin, 53(9): 105-113. [杨青青, 杨众养, 陈小花, 等, 2017. 热带海岸香蒲桃天然次生林群落优势种群种间联结性 [J]. 林业科学, 53(9):105-113.]
YANG XB, LIN Y, LIANG SQ, 1994. Forest vegetation of Wuzhishan of Hainan Island Ⅱ. Plant population analysis and forest structure analysis of wuzhishan forest vegetation [J]. Nat Sci J Hainan Univ, (4): 311-323. [楊小波, 林英, 梁淑群, 1994. 海南岛五指山的森林植被Ⅱ. 五指山森林植被的植物种群分析与森林结构分析 [J]. 海南大学学报(自然科学版), (4):311-323.]
ZHANG QM, CHEN BG, ZHOU GY, 2006. Interspecific association of the dominant species in two typical communities in Dinghushan, South China [J]. J S Chin Agric Univ, 27(1): 79-83. [张倩媚, 陈北光, 周国逸, 2006. 鼎湖山主要林型优势树种种间联结性的计算方法研究 [J]. 华南农业大学学报, 27(1):79-83.]
ZHANG XY,DU F, PANG TH, et al., 2019. Effects of soil moisture variation on competition among co-existing species in old field communities of the Loess Hilly Region [J]. Acta Ecol Sin, 39(3): 204-215. [张馨月, 杜峰, 潘天辉, 等, 2019. 土壤水分变化对黄土丘陵区撂荒群落共存种竞争的影响 [J]. 生态学报, 39(3):204-215.]
ZHU H, WANG H, LI BG, et al., 1998. Species diversity of primary tropical rain forest of South Yunnan of China with species reference to sampling area [J]. Biodivers Sci, (4): 1-7. [朱华, 王洪, 李保贵, 等, 1998. 滇南热带雨林物种多样性的取样面积探讨 [J]. 生物多样性, (4):1-7.]
(责任编辑 何永艳)
