黄土高原油松和辽东栎林下植物β多样性差异及影响因素
2021-06-10王世雄夏婷婷王孝安
王世雄 夏婷婷 王孝安

摘 要:β多樣性是生态学研究的热点论题,相同的β多样性格局可能由不同的生态过程所决定。该文通过构建零假说模型和典范变异分解的方法,比较了黄土高原油松人工林(Form. Pinus tabulaeformis)和辽东栎天然林(Form. Quercus wutaishanica)林下植物群落β多样性,确定了环境过滤和扩散限制在β多样性形成过程中的相对重要性。结果表明:(1)油松人工林和辽东栎天然林的林下群落均具有相似的β多样性大小,均呈现明显的种内聚集过程。(2)环境和空间共同解释了两种林型林下物种组成的较大变化(草本层[E+S]为33%~45%;灌木层[E+S]为21%~35%),且主要以环境解释量为主(包括纯环境变量[E | S]和空间化的环境变量[E ∩ S])。(3)油松人工林和辽东栎天然林林下群落β多样性形成的不同过程主要体现在生境异质性差异以及特定的环境因子间,例如,海拔梯度、枯落物厚度以及土壤养分(速效氮和速效钾)是引起两种林型林下物种差异的显著环境因子。综上结果认为,黄土高原的油松人工林和辽东栎天然林的群落构建均由确定性的环境过滤为主导,但两种林型的环境过滤过程并不一样,主要表现为生境异质性的差异和不同的环境限制因子。对于黄土高原区域生态环境建设,首先应该保证不同群落的生境异质性,同时不能忽视扩散限制及其与生境异质性二者交互作用对β多样性的影响。
关键词: 生境异质性, 扩散限制, 人工林, 种内聚集, 典范变异分解
中图分类号:Q948.15
文献标识码:A
文章编号:1000-3142(2021)03-0362-10
收稿日期:2020-02-23
基金项目:贵州省自然科学基金(黔科合基础[2019]1060);贵州省教育厅青年科技人才成长项目(黔教合KY字[2018] 098);贵州大学引进人才科研项目(贵大人基合字[2017]38号);青海师范大学中青年科研基金(2019zr011)[Supported by Natural Science Foundation of Guizhou Province ([2019]1060); Youth Science and Technology Talent Growth Program of Guizhou Provincial Department of Education ([2018]098); Program of Introducing Talents of Discipline to Guizhou University ([2017]38); Youth Foundation of Qinghai Normal University (2019zr011)]。
作者简介: 王世雄(1985-),博士,讲师,主要从事物种多样性保护与恢复生态学研究, (E-mail) wangshix812@163.com。
通信作者
Understory β diversity differences and influential factors
between Pinus tabulaeformis plantation and natural
Quercus wutaishanica forest on the Loess Plateau
WANG Shixiong1,2,3*, XIA Tingting2, WANG Xiaoan3
( 1. College of Life Sciences, Academy of Plateau Science and Sustainability, Qinghai Normal University, Xining 810008, China; 2. College of Forestry, Guizhou University, Guiyang 550025, China; 3. College of Life Sciences, Shaanxi Normal University, Xian 710119, China )
Abstract:Beta diversity is a hot topic in ecology, and the same patterns of β diversity may be determined by different ecological processes. Null model and canonical variation partitioning methods were used to determine the relative importance between environmental filter and dispersal limitation on community assembly between Pinus tabulaeformis plantation and natural Quercus wutaishanica forest. The results were as follows: (1) The understory communities of Pinus tabulaeformis plantation and natural Quercus wutaishanica forest had similar β diversities and similar β deviations with strong intraspecific aggregations of most species. (2) Environmental and spatial variables jointly explained the great variation of species composition of understory plant community in the two forest types (Herb layer: [E + S] was 33%-45%; Shrub layer: [E + S] was 21%-35%), and dominated by the environmental explanations (including pure environmental variables [E | S] and spatial environmental variables [E ∩ S]). (3) Although Pinus tabulaeformis plantation and natural Quercus wutaishanica forest on the Loess Plateau were both dominated by environmental filter, β diversities in both forests reflected dissimilar mechanisms of community assembly. It was mainly reflected in the differences of habitat heterogeneity and specific environmental factors between these two forests. Such as, altitude, litter depth and soil nutrients (available nitrogen and available potassium) were significant environmental factors that caused species differentiation between the two forest types. Therefore, it suggests that even if the same environmental filter process, it is necessary to compare the differences of regional environmental heterogeneity and further analyze the differences of the specific environmental factors that cause environmental filter. To restore species diversity effectively on the Loess Plateau, habitat heterogeneity is an important guarantee. At the same time, dispersal limitation caused by geographical distance should not be neglected, and special attention should be paid to interaction between dispersal limitation and habitat heterogeneity on β diversity.
Key words: habitat heterogeneity, dispersal limitation, plantation forest, intraspecific aggregation, canonical variation partitioning
β多样性连结了物种多样性格局与群落构建过程,是生态学研究的热点论题(Anderson et al., 2011; 李新辉等, 2016)。β多样性是很多陆地生态系统物种多样性的重要成分,但其维持机制仍存在很大的争议(Soininen et al., 2007)。生态位理论认为,群落中的物种是确定的,每个物种都有各自的适应环境,不同的环境条件决定了适应特定环境的物种,群落内物种要共存就必须有生态位分化;表现在β多样性方面即为环境差异越大,物种组成的差异就越大,也就是说环境因子能较好地解释β多样性格局,即环境过滤。而扩散限制理论认为,由于群落内的物种具有相同的竞争能力、相同的出生率和死亡率,群落内的物种是随机的,扩散限制是决定群落β多样性格局的重要过程。然而,大量研究表明,群落的构建并不是由任何一个过程单独完成,而可能受环境过滤和扩散限制共同影响(赵鸣飞等, 2017),二者的相对贡献与群落类型和所研究的尺度有关。
从1960年开始,油松(Pinus tabulaeformis)作为主要造林树种在黄土高原大量种植,旨在促进该区植被恢复,加速生态系统重建(刘立品, 1998; 王世雄等, 2014)。Wang et al.(2014)研究表明,经过50多年的恢复演替,油松人工林(Form. Pinus tabulaeformis)已显著改善了其林下层的物种组成。油松人工林和该区的辽东栎天然林(Form. Quercus wutaishanica)的物种多样性具有相似的多尺度格局,都表现为物种丰富度随空间尺度的增加而增加,大尺度上的β多样性都是两种林型物种多样性的主要成分(王世雄, 2013; Wang et al., 2014)。过程决定格局,格局反映过程(Loreau, 2000),但相似的物种多样性格局并不一定表明油松人工林和辽东栎天然林具有相似的群落构建过程,而揭示群落构建过程是指导区域生态环境建设的重要理论基础(王世雄, 2013)。
王世雄等(2013)研究表明,环境过滤和扩散限制共同影响着黄土高原森林的群落构建过程。但是,针对特定的群落类型,扩散限制和环境过滤的相对贡献仍不得而知。尤其是对于具有相似多样性格局的群落,揭示其构建过程是理解多样性格局形成机制的重要途径。该研究旨在度量环境过滤和扩散限制两种重要的生态过程对物种多样性格局的相对贡献,揭示油松人工林和辽东栎天然林在群落构建过程中的差异,为黄土高原物种多样性的恢复与保护提供理论依据和实践指导。
1 研究区概况与研究方法
1.1 研究区概况
研究区域位于黄土高原中部的子午岭,属暖温带半湿润气候,最高海拔约为1 700 m,最低海拔约为1 200 m。该区光照充足,湿热同期,年平气温为7 ℃,≥0 ℃积温为3 134 ℃。降水季节分布不均,旱涝相间(刘立品, 1998)。林区土壤为棕色森林土,土层厚,结构疏松,林下枯枝落叶较厚,但腐化不好。
由于强烈的人类活动导致该区森林植被大面积减少(王世雄, 2013),现存的森林大部分是屡遭破坏后形成的天然栎类次生林,占林地总面积的74%,主要包括辽东栎林(Form. Q. wutaishanica),少量次生白桦林(Form. Betula platyphylla)、山杨林(Form. Populus davidiana)及二者与辽东栎的混交林等(王世雄等, 2014)。为了恢复该区植被,自20世纪六七十年代起,政府实施了诸如“天然林保护工程”等多项造林措施,造林树种依局域生境特征主要有油松、刺槐(Robinia pseudoacacia)、侧柏(Platycladus orientalis)、山杏(Armeniaca sibirica)和沙棘(Hippophae rhamnoides)等。其中,油松通过人工栽植或飞机播种等多种方式种植成为了黄土高原面积最大的人工林,截止到1993年底,油松人工林面积已达到4百万公顷,占黄土高原子午岭林地总面积的11%(刘立品, 1998)。
1.2 样地设置
依据实地勘察结果,选择典型的油松人工林和辽东栎天然林各设置3个样地,每个样地设置5个20 m × 20 m的样方,在样方的每个象限设置小样方进行林下物种调查。其中,灌木层的小样方为4 m × 4 m,草本层的小样方为1 m × 1 m。依据经典植物群落学方法对每个小样方进行群落调查,记录每个个体的物种名、高度和盖度等。同时,记录每个样方的环境因子以及空间位置。环境因子包括林冠郁闭度、海拔、坡度、坡向、坡位、枯落物和腐殖质厚度等。在每个小样方各取0~30 cm的土壤样品3个,带回实验室风干后测定土壤速效氮、速效磷、速效钾、有机质以及pH值。土壤pH用电位法测定;有机质用重铬酸钾容量法测定;速效氮用硝态氮和氨态氮的总和表示,用AA3连续流动化学分析仪测定;速效磷含量用钼锑抗比色法测定;速效钾采用火焰光度法测定(鲍士旦, 2000)。
1.3 数据分析
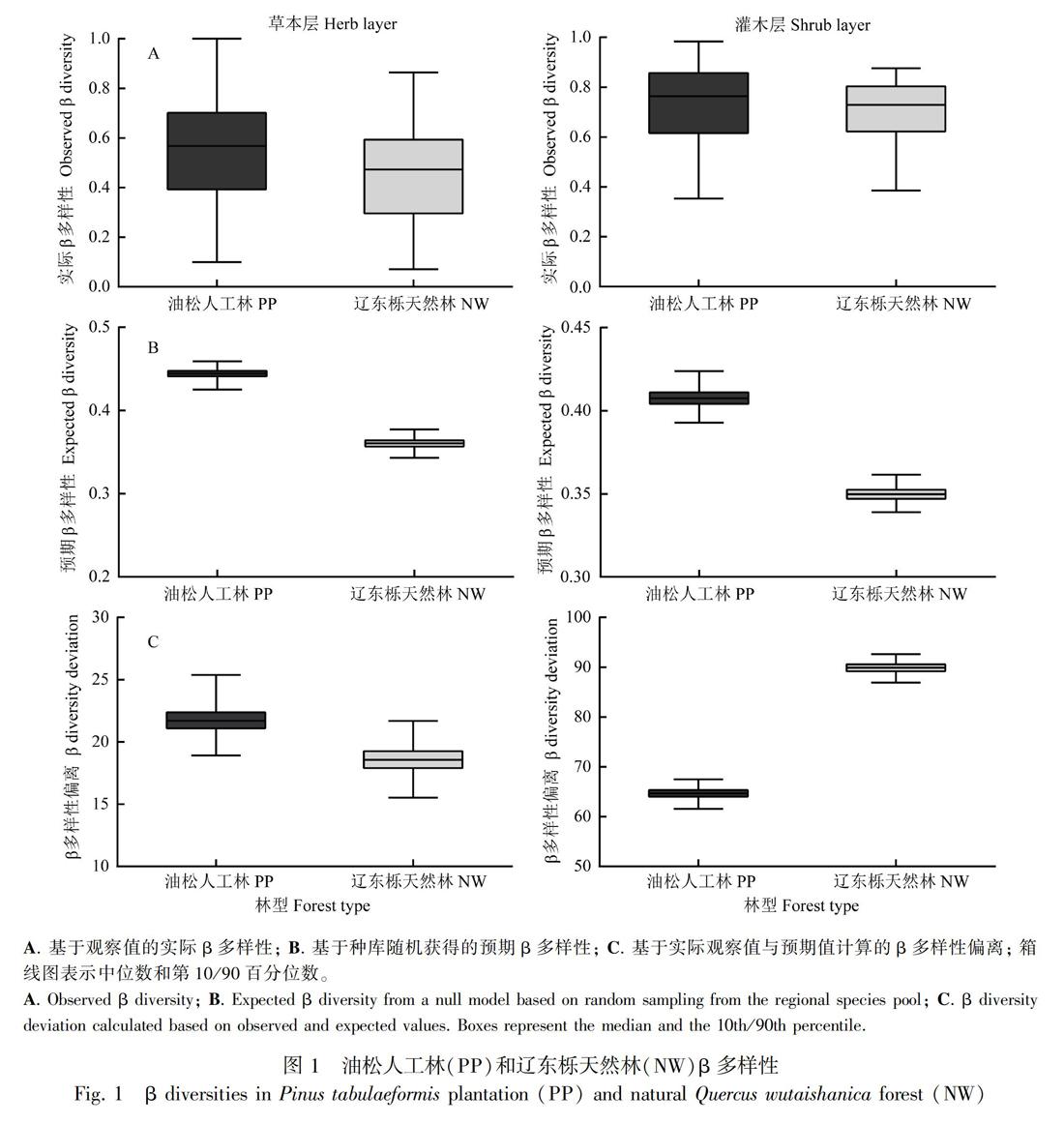
通过构建基于物种个体的完全随机零模型去除环境过滤和扩散限制的作用,比较实际观察的β多样性和随机群落β多样性间的差异来揭示群落构建过程(Myers et al., 2013)。零模型构建步骤如下:第一,将每个林型的所有物种定义为该群落的区域种库;第二,保持每个样方中的个体多度不变,从种库中随机抽取物种进行样方重建;第三,重复上面两个步骤499次,计算实际观察的β多样性和预期β多样性间的差异,记为β偏离,其大小等于观察值与预期值的差与预期值的标准差的商。其中,β多样性用基于物种多度数据的Bray-Curtis指数来计算。β偏离接近于0,意味着群落的构建过程是完全随机,不受环境过滤和扩散限制的作用;正的β偏離表示实际β多样性明显高于随机值,即实际群落的相似度会低于随机群落,表明实际群落中物种呈聚集分布,可能是环境过滤或者扩散限制亦或二者共同作用;而负的β偏离表示实际β多样性明显小于随机的β多样性,表明实际群落是均匀分布的(谭凌照等, 2017)。两个林型间的实际观察的β多样性、预期β多样性以及β偏离通过基于非参数模型的方差分析进行比较(Anderson et al., 2011)。
环境梯度以及空间梯度上的变异分解方法提供了研究群落构建过程的重要途径(Gilbert et al., 2004; Legendre et al., 2009)。如果群落中的物种组成变异(β多样性)与样地间的环境紧密相关,表明环境过滤起主要作用(Valencia et al., 2004);
而如果β多样性与空间因子的变化密切相关,表明扩散限制起主要作用(Hubbell, 2001)。环境过滤和扩散限制的相对大小通过对环境和空间因子的典范变异分解方法度量。其中,空间因子使用主轴邻距法(principal coordinates of neighboring matrices, PCNM)根据样方的坐标产生(Peres-Neto et al., 2006)。环境因子进行自然对数转化(Kristiansen et al., 2012),物种多度数据进行‘Hellinger转化(Legendre & Gallagher, 2001)。环境和空间因子通过前向选择的方法(Monte Carlo permutation, n=999)筛选出显著因子(P<0.1)后再进行典范变异分解(王世雄等, 2014),即通过冗余分析(redundancy analysis, RDA)及偏冗余分析(partial RDA, pRDA)实现(王世雄, 2013)。物种组成变化的解释量分解为如下3个部分:纯环境解释量[E | S]、纯空间解释量[S | E]以及空间化的环境解释量[E∩S](Laliberté et al., 2009; 王世雄, 2013)。该研究采用矫正的R2来度量每个解释量的贡献率(Peres-Neto et al., 2006)。所有分析通过R软件实现(R Core Team, 2019)。
2 结果与分析
油松人工林和辽东栎天然林两种林型共调查记录到草本层物种111种,灌木层物种86种。其中,油松人工林草本层物种70种,灌木层物种64种;辽东栎天然林草本层物种75种,灌木层物种66种。总体上,辽东栎天然林具有比油松人工林较高的α多样性(物种丰富度),即辽东栎天然林的样方具有比油松人工林较多的物种数。对于草本层,油松人工林的α多样性为11.80±3.12,天然辽东栎的为12.70±5.65;对于灌木层,油松人工林的α多样性为18.00±6.10,天然辽东栎的为22.53±5.24。
两种林型的物种呈现了明显的分布不均匀性。例如,对于草本层,油松人工林稀有种(频度≤5%小样方)达52种,常见种(频度≥25%小样方)4种;辽东栎林稀有种48种,常见种2种。其中,很多稀有种仅出现在单个样方中,且多度较小,而常见种的多度相对较大,如草本层的细叶薹草(Carex duriusata)在两种林型中均占有绝对的优势地位。同样的,灌木层的物种多度在稀有种和常见种间也呈现了明显的两极分化。
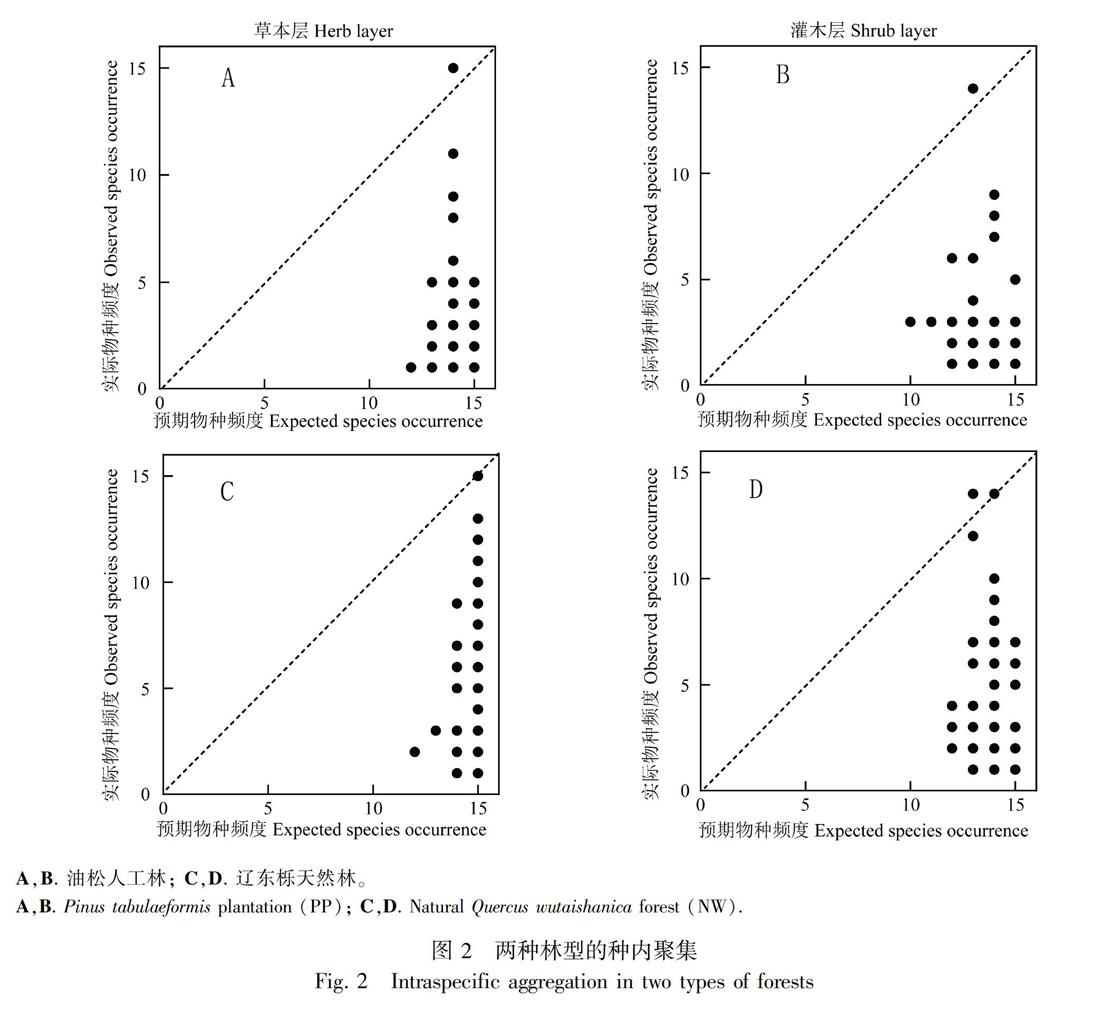
油松人工林和辽东栎天然林的林下群落均具有相似的β多样性大小,且没有显著性差异(图1;草本层:F=0.93,P=0.34;灌木层:F=0.41,P=0.52)。但是,两种林型的预期β多样性以及β偏离均具有显著性差异(P<0.05),油松人工林具有显著高于辽东栎天然林的预期β多样性。重要的是,两种林型的β偏离均呈现了明显的正相关关系,即实际β多样性明显大于预期β多样性,这反映了两种林型中的绝大多数物种呈现了很强的种内聚集过程(图2)。
为揭示种内聚集形成的原因,本研究利用典范变异分解方法进行了分析。结果表明,环境和空间因子共同解释了两种林型林下物种组成的较大变异(草本层:[E + S] = 33%~45%;灌木层:[E + S] = 21%~35%)。其中,环境变量(包括纯环境变量[E | S]和空间化的环境变量[E ∩ S])的解释率占总解释量的50%以上(图3)。空间因子、环境以及二者的交互作用对油松人工林林下群落物种组成变化的解释量高于辽东栎天然林,且以环境解释量[E]为主。同时,两种林型具有相似的(草本层16%~18%;灌木层4%~5%)纯空间解释量[S | E],这反映了两种林型相似的扩散限制过程。
RDA分析的结果表明,尽管两种林型均呈现了相似的环境解释率,但是,引起两种林型林下物种组成变异的环境因子并不相同(图4)。海拔(altitude)以及枯落物厚度(litter depth)是引起两种林型草本层物种差异的显著环境因子,而海拔、土壤速效氮(N)和速效钾(K)以及枯落物厚度是引起两种林型灌木层物种差异的显著环境因子(图4:A-B)。对于特定的林型,不同层次的林下群落也呈现了明显不同的环境依赖性。影响辽东栎林草本层的显著环境因子为海拔、土壤pH值以及林冠郁闭度(canopy openness),而土壤pH值和土壤湿度(soil moisture)是影响其灌木层的显著环境因子(图4:C-D);影响油松人工林草本层的显著环境因子为土壤腐殖质厚度(humus thickness)和坡度 (slope degree),而土壤速效氮(N)和枯落物厚度是影响其灌木层的主要环境因子(图4:E-F)。
3 讨论
β多样性作为群落物种多样性的重要成分,能够将多样性格局与群落构建过程联系起来。研究表明,油松人工林和辽东栎天然林两种林型具有相似的β多样性大小以及显著大于0的β偏离,并且呈现了明显的种内聚集过程,这与施晶晶等(2019)的研究结果相似,即环境过滤和扩散限制二者相互作用,共同影响各自β多样性的形成过程。本研究中,無论是黄土高原的油松人工林还是辽东栎天然林都属于典型的温带森林, 二者均存在一定的环境条件限制,环境变量(包括纯环境因子和空间化的环境因子的贡献)解释了两种林型林下群落物种组成的较大变化,且以空间化的环境因子([E ∩ S])为主,印证了环境过滤的确是温带森林群落构建的重要过程(Myers et al., 2013; 王世雄等, 2013)。
但是,相似的β多样性以及相似的环境解释量并不能说明相似的群落构建过程。研究表明,尽管温带森林和热带森林具有相似的β多样性格局(Kraft et al., 2011),但却显示了截然不同的群落构建过程(Myers et al., 2013)。群落β多样性主要受物种聚集程度的影响,生境过滤和扩散限制都能增加同种聚集程度。由于温带森林环境较热带森林严酷,其较高的种内聚集是由于环境过滤所致,主要表现为较低的物种多样性以及较高的环境过滤过程,而热带森林较高的种内聚集是由于扩散限制所致,主要表现为较高的空间依赖性,即较强的扩散限制(Myers et al., 2013)。本研究中,无论是油松人工林还是辽东栎天然林均呈现了较高的种内聚集过程,导致了实际β多样性明显大于预期β多样性,环境过滤是解释两种林型种内聚集的重要过程。
环境过滤对β多样性的影响,可能更依赖于生境的异质性(Myers et al., 2013)。从RDA排序图可以看出,辽东栎天然林的样方散布相对均匀,而油松人工林的部分样方间具有明显的聚集现象,这说明辽东栎天然林样方间的β多样性都比较高,而油松人工林较高的β多样性主要由部分样方间的生境异质性所致。同时,生境异质性可以通过种间相互作用增强物种聚集格局,进而影响β多样性格局。种间竞争使某些竞争能力弱的物种往往被优势种排挤,这对于林下物种更是如此,从而提高了优势种在优越环境中的聚集程度(Lancaster, 2006; 谭凌照等, 2017)。与热带森林相比,温带森林的物种少,且比较常见(Pitman et al., 2001; Gilbert et al., 2010)。该研究中,生境异质性对两种林型物种聚集的影响主要表现在常见种和稀有种种类数量间的较大差异以及特定样方中常见种和稀有种多度间的较大差异,这在两种林型的草本层更明显。
同时,环境过滤对β多样性的影响还表现为特定的环境因子方面。例如,海拔作为一个综合环境因子,反映了物种的扩散和定居的综合结果,是两种林型林下物种差异的重要环境因子。同时,油松人工林林下较厚的松针厚度及其较慢的分解速率都明显不利于其林下物种的定居(Barlow et al., 2007; 王世雄, 2013)。另外,林冠郁闭度、土壤养分等都是影响林下物种定居的重要环境因子,决定了林下的物种多样性大小。本研究结果表明,即使是相同的环境过滤过程,也应该进一步分析引起环境过滤的具体环境因子,特别是对于不同的群落类型,应针对不同的环境限制因子采取不同的恢复措施。
尽管环境过滤是主导油松人工林(王世雄等, 2014)和辽东栎天然林群落构建的重要过程,但两种林型显著的纯空间因子的解释量([S | E])说明,扩散限制也是两种林型群落构建过程中不可忽视的重要过程。前人研究表明,林下绝大多数物种的扩散距离十分有限(Bierzychudek, 1982; Whigham, 2004),常被限制在一个生境或者单个样地中,即稀有种(王世雄, 2013),而稀有种主要在较大尺度上影响群落的物种多样性格局(王世雄等, 2018)。扩散限制使得稀有种很难拓殖到区域内所有适宜的环境中,从而加剧了同种间的聚集程度,增强了扩散限制对群落β多样性的影响(谭凌照等, 2017)。同时,扩散限制也依赖于生境的连接程度(王世雄, 2013)。黄土高原地区沟壑纵横,加上人类活动的影响,使得黄土高原的生境破碎化十分明显。同时由于受到栽种条件限制,油松人工林镶嵌于不同天然次生林的周围而呈斑块状分布。生境的不连续造成隔离效应,改变了种群的迁移能力和迁入模式、种群遗传和变异等,影响了物种的繁殖和扩散,从而显著地降低局域上的物种多样性(Fralish et al., 1991;王世雄, 2013)。
无论是油松人工林还是辽东栎天然林,β多样性均是两种林型物种多样性的重要支撑成分(王世雄, 2013)。因此,保护好β多样性是保护黄土高原的物种多样性的首要任务。β多样性是物种种类对生境异质性的反映,要保护好β多样性,需要注意以下两个问题: 第一,生境异质性对黄土高原物种多样性恢复的重要性,特别要有针对性地创造每个林型限制性环境因子的异质性生境可能是维持两种林型β多样性的重要前提。例如,对于油松人工林物种多样性的保护,可以重点创造林下枯落物和腐殖质的异质性;而对于辽东栎天然林,可以考虑创造土壤养分以及林冠郁闭度的异质性等。第二,不能忽视由于地理距离引起的扩散限制及其与生境异质性二者交互作用对β多样性的影响。例如,在不同天然林周围植树造林是一个很好维持人工林群落异质性的重要措施,不同的天然林由于扩散限制而决定了不同的人工林种源,加之有效的生境異质性,这将会在很大程度上有助于维持人工林林下物种多样性。
4 结论
油松人工林和辽东栎天然林的林下群落均具有相似的β多样性大小,且两种林型均呈现了明显的种内聚集过程。与很多研究结果一致,黄土高原的油松人工林和辽东栎天然林的群落构建由确定性的环境过滤决定,且以空间化的环境为主导。但是,两种林型的环境过滤过程并不一样,主要表现为不同的生境异质性和不同的环境因子。因此,即使是相似的环境过滤过程,也应该比较区域环境异质性的差异,并进一步分析引起环境过滤的特定环境因子。所以,对于黄土高原区域生态环境建设,应该针对不同群落类型采取差异化的恢复措施:第一,生境异质性对黄土高原物种多样性恢复的重要性,特别要有针对性地创造每个林型限制性环境因子的异质性生境可能是维持两种林型β多样性的重要前提。第二,不能忽视由于地理距离引起的扩散限制及其与生境异质性二者交互作用对β多样性的影响。
参考文献:
ANDERSON MJ, CRIST TO, CHASE JM, et al., 2011. Navigating the multiple meanings of β diversity: A roadmap for the practicing ecologist [J]. Ecol Lett, 14(1): 19-28.
BAO SD, 2000. Soil and agricultural chemistry analysis [M]. 3rd ed. Beijing: China Agriculture Press. [鮑士旦, 2000. 土壤农化分析 [M]. 3版. 北京: 中国农业出版社.]
BARLOW J, GARDNER TA, FERREIRA LV, et al., 2007. Litter fall and decomposition in primary, secondary and plantation forests in the Brazilian Amazon [J]. For Ecol Manage, 247(1-3): 91-97.
BIERZYCHUDEK P, 1982. Life histories and demography of shade-tolerant temperate forest herbs: A review [J]. New Phytol, 90(4): 757-776.
CONDIT R, PITMAN N, LEIGH EG, et al., 2002. Beta-diversity in tropical forest trees [J]. Science, 295(5555):666-669.
FERNANDEZ-GOING BM, HARRISON SP, ANACKER BL, et al., 2013. Climate interacts with soil to produce beta diversity in Californian plant communities [J]. Ecology,94(9): 2007-2018.
FRALISH JS, CROOKS FB, CHAMBERS JL, et al., 1991. Comparison of presettlement, second-growth and old-growth forest on six site types in the Illinois Shawnee Hills [J]. Am Midland Nat, 125(2): 294-309.
FREESTONE AL, INOUYE BD, 2006. Dispersal limitation and environmental heterogeneity shape scale-dependent diversity patterns in plant communities [J]. Ecology,87(10): 2425-2432.
GILBERT B, LECHOWICZ MJ, LEVIN SA, 2004. Neutrality, niches, and dispersal in a temperate forest understory [J]. Proc Natl Acad Sci USA, 101(20): 7651-7656.
GILBERT GS, HOWARD E, AYALA-OROZCO B, et al., 2010. Beyond the tropics: Forest structure in a temperate forest mapped plot [J]. J Veg Sci, 21(2): 388-405.
HANNA T, KALLE R, MARKKU YH, 2003. Dispersal, environment, and floristic variation of western Amazonian forests [J]. Science, 299(5604): 241-244.
HUBBELL SP, 2001. The unified neutral theory of biodiversity and biogeography [M]. New Jersey: Princeton University Press.
KRAFT NJB, COMITA LS, CHASE JM, et al., 2011. Disentangling the drivers of β-diversity along latitudinal and elevational gradients [J]. Science, 333(6050): 1755-1758.
KRAFT NJB, VALENCIA R, ACKERLY DD, 2008. Functional traits and niche-based tree community assembly in an Amazonian forest [J]. Science, 322(5901): 580-582.
KRISTIANSEN T, SVENNING J-C, EISERHARDT WL, et al., 2012. Environment versus dispersal in the assembly of western Amazonian palm communities [J]. J Biogeogr,39(7): 1318-1332.
LALIBERT E, PAQUETTE A, LEGENDRE P, et al., 2009. Assessing the scale-specific importance of niches and other spatial processes on beta diversity: A case study from a temperate forest [J]. Oecologia, 159(2): 377-388.
LANCASTER J, 2006. Using neutral landscapes to identify patterns of aggregation across resource points [J]. Ecography, 29(3): 385-395.
LEGENDRE P, GALLAGHER ED, 2001. Ecologically meaningful transformations for ordination of species data [J]. Oecologia, 129(2): 271-280.
LEGENDRE P, MI X, REN H, et al., 2009. Partitioning beta diversity in a subtropical broad-leaved forest of China [J]. Ecology, 90(3): 663-674.
LI XH, LIU YH, LIU Y, et al., 2016. Impacts of geographical distances and environmental differences on the beta diversity of plant communities in the dry-hot valley of the Yuanjiang River [J]. Biodivers Sci, 24(4): 399-406. [李新輝, 刘延虹, 刘晔, 等, 2016. 地理距离及环境差异对云南元江干热河谷植物群落beta多样性的影响 [J]. 生物多样性,24(4): 399-406.]
LIU LP, 1998. Flora arborum et fruticum Ziwu Mountain [M]. Lanzhou: Lanzhou University Press. [刘立品, 1998. 子午岭木本植物志 [M]. 兰州: 兰州大学出版社.]
LOREAU M, 2000. Are communities saturated? On the relationship between α, β and γ diversity [J]. Ecol Lett,3(2): 73-76.
MYERS JA, CHASE JM, JIMNEZ I, et al., 2013. Beta-diversity in temperate and tropical forests reflects dissimilar mechanisms of community assembly [J]. Ecol Lett, 16(2): 151-157.
PERES-NETO PR, PIERRE L, STPHANE D, et al., 2006. Variation partitioning of species data matrices: estimation and comparison of fractions [J]. Ecology,87(10): 2614-2625.
PITMAN NCA, TERBORGH JW, SILMAN MR, et al., 2001. Dominance and distribution of tree species in upper amazonian terra firme forests [J]. Ecology, 82(8):2101-2117.
PLOTKIN JB, POTTS MD, LESLIE N, et al., 2000. Species-area curves, spatial aggregation, and habitat specialization in tropical forests [J]. J Theor Biol, 207(1): 81-99.
R CORE TEAM, 2019. R: A language and environment for statistical computing [DB/OL]Foundation for Statistical Computing, Vienna, Austria. https://www.R-Project.org.
RAO MD, FENG G, ZHANG JL, et al., 2013. Effects of environmental filtering and dispersal limitation on species and phylogenetic beta diversity in Gutianshan National Nature Reserve [J]. Chin Sci Bull, 58(13): 1204-1212. [饶米德, 冯刚, 张金龙, 等, 2013. 生境过滤和扩散限制作用对古田山森林物种和系统发育β多样性的影响 [J]. 科学通报, 58(13): 1204-1212.]
SHEN G, YU M, HU XS, et al., 2009. Species-area relationships explained by the joint effects of dispersal limitation and habitat heterogeneity [J]. Ecology, 90(11): 3033-3041.
SHI JJ, ZHAO MF, WANG YH, et al., 2019. Community assembly of herbaceous layer of the planted forests in the central Loess Plateau, China[J]. Chin J Plant Ecol, 43(9): 834-842. [施晶晶, 赵鸣飞, 王宇航, 等, 2019. 黄土高原腹地人工林下草本层群落构建机制[J]. 植物生态学报, 43(9): 834-842.]
SOININEN J, MCDONALD R, HILLEBRAND H, 2007. The distance decay of similarity in ecological communities [J]. Ecography, 30(1): 3-12.
TAN LZ, FAN CY, ZHANG CY, et al., 2017. Effects of local ecological processes on community similarity in the Jiaohe broadleaved Korean pine forests in Jilin Province [J]. Sci Silv Sin, 53(11): 12-19. [谭凌照, 范春雨, 张春雨, 等, 2017. 局域生态过程对吉林蛟河阔叶红松林群落相似度的影响 [J]. 林业科学, 53(11): 12-19.]
VALENCIA R, FOSTER RB, VILLA G, et al., 2004. Tree species distributions and local habitat variation in the amazon: large forest plot in eastern Ecuador [J]. J Ecol,92(2): 214-229.
WANG SX, 2013. Spatio-temporal patterns and processes of plant community species diversity in Ziwuling, Loess Plateau [D]. Xian: Shaanxi Normal University. [王世雄, 2013. 黄土高原子午岭植物群落物种多样性的时空格局与过程(博士学位论文) [D]. 西安: 陕西师范大学.]
WANG SX, GUO H, WANG XA, et al., 2013. Dispersal limitation versus environment filtering in the assembly of plant communities in the Ziwu mountains [J]. Sci Agric Sin, 46(22): 4733-4744. [王世雄, 郭华, 王孝安, 等, 2013. 扩散限制和环境筛选对子午岭森林群落构建的相对贡献 [J]. 中国农业科学, 46(22): 4733-4744.]
WANG SX, GUO H, WANG XA, et al., 2014. Multiple-scale assessment of understory herb species diversity in pine plantations after long-term restoration [J]. Nord J Bot,32(5): 680-688.
WANG SX, WANG XA, GUO H, et al., 2014. Multiple-scale patterns in understory species diversity of pine plantations in the Loess Plateau [J]. Arid Zone Res, 31(2): 294-301. [王世雄, 王孝安, 郭華, 等, 2014. 黄土高原油松林林下物种多样性的多尺度格局 [J]. 干旱区研究,31(2): 294-301.]
WANG SX, WANG XA, GUO H, et al., 2018. Relative contributions of rare and common species to the multiple-scale patterns of species diversity in Quercus wutaishanica communities on the Loess Plateau [J]. Acta Ecol Sin, 38(22): 8060-8069. [王世雄, 王孝安, 郭华, 等, 2018. 稀有种和常见种对黄土高原辽东栎群落物种多样性贡献的多尺度分析 [J]. 生态学报, 38(22): 8060-8069.]
WHIGHAM DF, 2004. Ecology of woodland herbs in temperate deciduous forests [J]. Ann Rev Ecol Evol Syst, 35:583-621.
ZHAO MF, WANG GY, XING KX, et al., 2017. Patterns and determinants of species similarity decay of forest communities in the western Qinling mountains [J]. Biodivers Sci, 25(1): 3-10. [赵鸣飞, 王国义, 邢开雄, 等, 2017. 秦岭西部森林群落相似性递减格局及其影响因素 [J]. 生物多样性, 25(1): 3-10.]
(责任编辑 李 莉)
