芥菜胞质雄性不育系结荚性状的筛选和细胞学鉴定
2021-06-08刘志新徐玉颖姚培杰许可净韩一鸣傅强万正杰
刘志新,徐玉颖,姚培杰,许可净,韩一鸣,傅强,万正杰
华中农业大学园艺林学学院/园艺植物生物学教育部重点实验室,武汉 430070
芥菜(Brassicajuncea)是一种原产于我国的十字花科芸薹属蔬菜[1]。研究表明,西北地区为我国芥菜的初生起源地,四川盆地为我国芥菜的次生起源地[2]。目前,芥菜在全国各地均有栽培,其中,长江中下游地区和西南地区是主产区[3-5]。由于芥菜具有营养物质丰富、栽培容易、多样化的加工与食用方式等特点,深受消费者欢迎[6]。芥菜作为一种常异花授粉植物,新品种主要来自于系统育种和优势杂交育种[7],芥菜杂交种的营养体在生长势、抗逆性、丰产性等方面较常规种具有明显优势[8],杂交种生产过程中,人工授粉存在种子生产成本过高、生产效率低下的问题,因此,细胞质雄性不育系在芥菜杂交种选育和制种中得到了大规模的应用[9]。
长期以来,雄性不育作为植物杂种优势利用和生殖发育研究的热点内容之一,已取得大量研究成果[10]。余凤群等[11]在对甘蓝型油菜花药败育的研究中发现,花药败育主要分为小孢子发育异常、花粉母细胞发育异常、花粉囊发育异常3类。在微观水平上,胡永敏等[12]进行了5种甘蓝型油菜细胞质雄性不育系的细胞学观察实验,发现PolCMS材料的败育大多发生在孢原细胞时期;许代香等[13]通过对甘蓝型油菜显性核不育系D3A的花药败育研究发现,不育系的花药败育主要是花粉囊的异常发育所导致。而ogu细胞质不育系材料则存在多种败育方式,黄邦全等[14]通过种间杂交方式,将ogu萝卜雄性不育胞质转入紫菜薹中,经过多代回交,获得了发育正常的紫菜薹ogu胞质不育系,其花药败育发生在孢原细胞分化期;田福发[15]通过种间杂交获得了Pol红菜薹不育系、ogu红菜薹不育系材料,并且进行细胞学切片观察,发现ogu红菜薹不育系花药在四分体时期,绒毡层细胞发育异常,挤压四分体,最终导致绒毡层和四分体同时解体,引起花药败育。我们在实践过程中发现,芥菜杂交育种过程中,以雄性不育系材料为母本创制杂交种,部分不育系材料会表现出不同程度的结荚畸形现象。为了揭示芥菜雄性不育系结荚畸形性状的形成机制,笔者对59份芥菜材料进行了结荚情况的鉴定以及细胞学研究,以期为芥菜胞质雄性不育系的品种改良提供理论依据。
1 材料与方法
1.1 试验材料
试验材料为华中农业大学十字花科蔬菜遗传育种课题组所收集来自全国各地的59份芥菜材料,经多年提纯复壮,如表1所示,包括29份芥菜保持系、30份芥菜细胞质雄性不育系、hauCMS原始材料(FanY-9A)及其保持系(FanY-9B)。
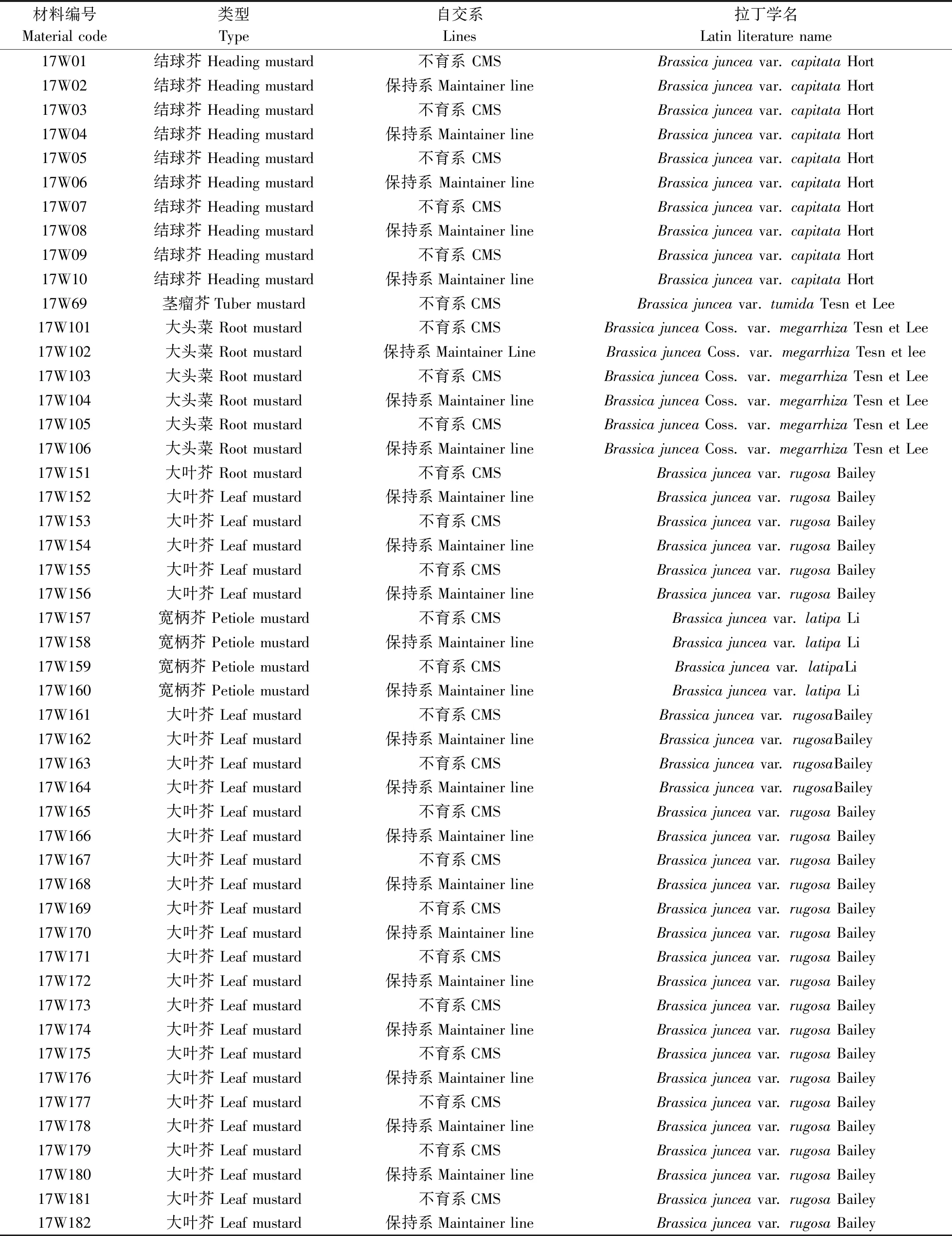
表1 供试材料Table 1 The list of experimental materials
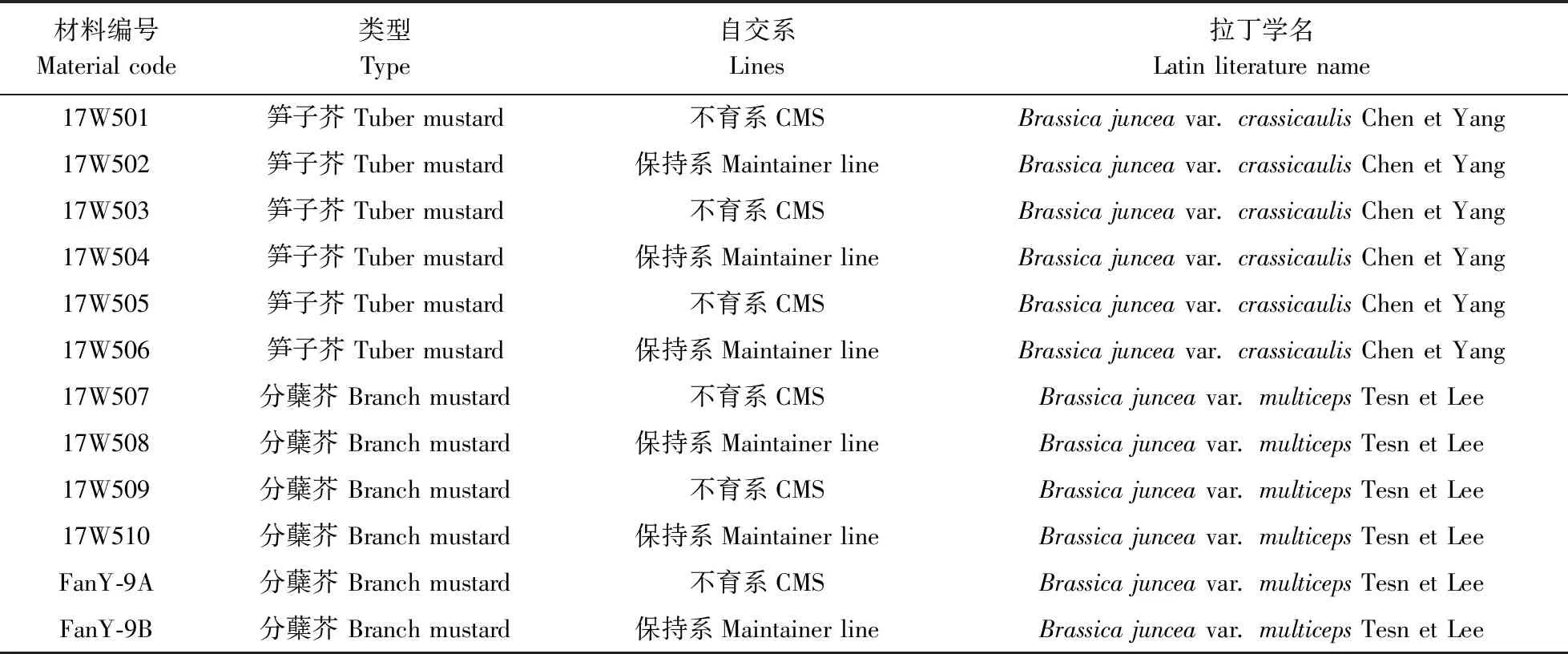
续表1 ContinuedTable 1
1.2 芥菜胞质雄性不育系材料不育类型鉴定
利用CTAB法提取30份芥菜细胞质雄性不育基因组总DNA,获得基因orf288和orf138的保守序列[16],利用NCBI在线网页设计引物,orf138正向引物:GAAACGGGAAGTGACAATAC,反向引物:GCATTATTTTCTCGGTCCAT;orf288正向引物CAATGGCTATTACCCCTGCT;反向引物: ATGCCGGGCATTTACCTGATGG。
1.3 芥菜材料的结荚情况调查
将59份材料于2017年9月同期播种育苗,之后统一水肥管理,杂交授粉完成后,待种荚成熟时,取典型角果,拍照记录。
1.4 花蕾的半薄切片技术
材料盛花期,选取生长良好的花序参照文献[17]制样,于显微镜下观察。
2 结果与分析
2.1 芥菜不育系材料不育胞质类型鉴定
利用CTAB法提取30份芥菜细胞质雄性不育系材料,利用设计的2对引物进行PCR扩增、琼脂糖凝胶电泳检测,结果(图1)显示:以17W05、17W155、17W179为代表的23份材料属于hau不育胞质类型,以17W501和17W173为代表的7份材料属于ogu不育胞质类型。
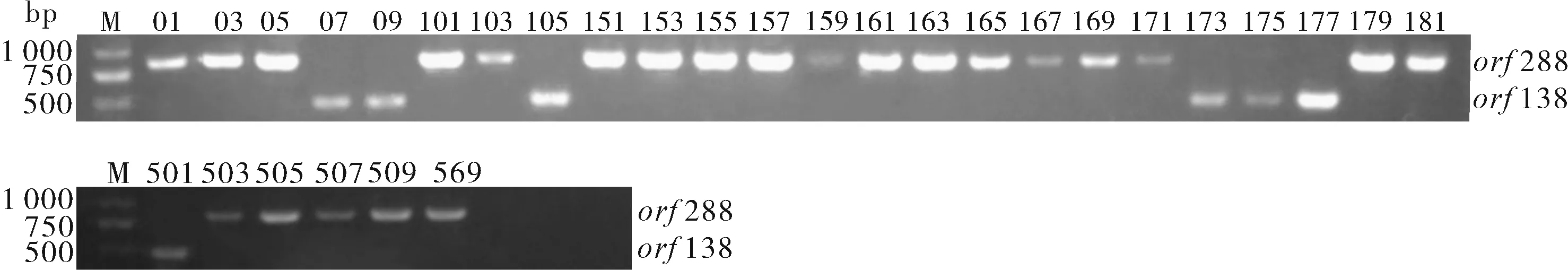
图1 芥菜雄性不育系材料不育类型鉴定Fig.1 Identification of CMS types of Brassica juncea
2.2 芥菜细胞质雄性不育系的结荚形态
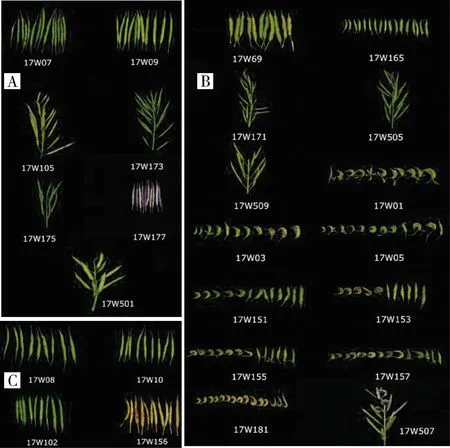
对59份芥菜材料结荚形态的调查结果(图2)显示,7份芥菜ogu雄性不育系没有畸形情况发生,荚果饱满直立,形态具有一致性(图2A);芥菜hau雄性不育系材料在畸形方面表现较大的差异(图2B),其中,17W165、17W171、17W505、17W509和17W69表现出良好的结荚特性,果荚直立饱满,其他材料则表现出不同程度的畸形;所有的芥菜保持系材料荚果均呈现饱满、直立的状态(图2C)。
2.3 芥菜细胞质不育系花药败育及花器官发育的细胞学研究
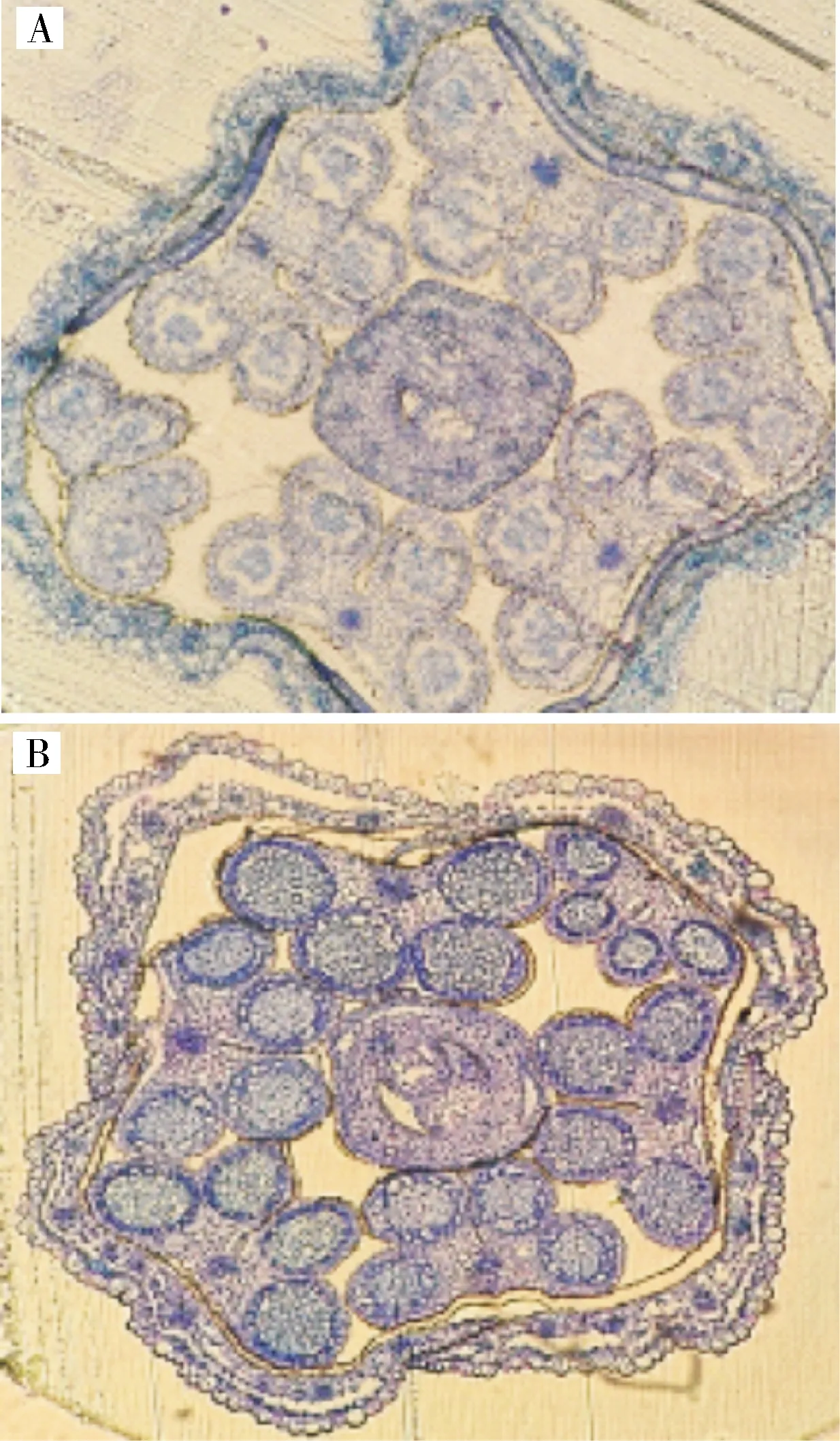
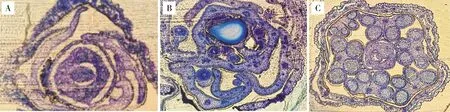
1)芥菜oguCMS材料花药败育及花器官发育观察。对芥菜ogu细胞质雄性不育系17W173及其对应保持系材料17W174进行半薄切片的细胞学观察(图3),可以发现,从花器官的整体发育来看,不育系绒毡层出现降解,小孢子集结成团不能分散,保持系则较为正常。进一步观察其花药的败育情况(图4),可以发现,四分体时期,绒毡层细胞发育异常,细胞出现空泡化现象;单核靠边期,绒毡层发生降解,药室中央的小孢子聚集成团;双核期,绒毡层已经基本降解;三核期时,小孢子出现空泡化迹象,花药败育。
A.ogu细胞质雄性不育系;B. hau细胞质雄性不育系;C.保持系。 A.ogu CMS;B.hau CMS; C.Maintainer lines.
A.ogu细胞质雄性不育系; B.保持系。A.ogu CMS; B.Maintainer line.
A.四分体时期; B.单核期; C.单核靠边期; D.双核期; E.三核期; F.释放花粉。A.Tetrad; B.Mononuclear; C.Mononuclear marginal; D.Dikaryotic; E.Trinuclear; F.Pollen release.
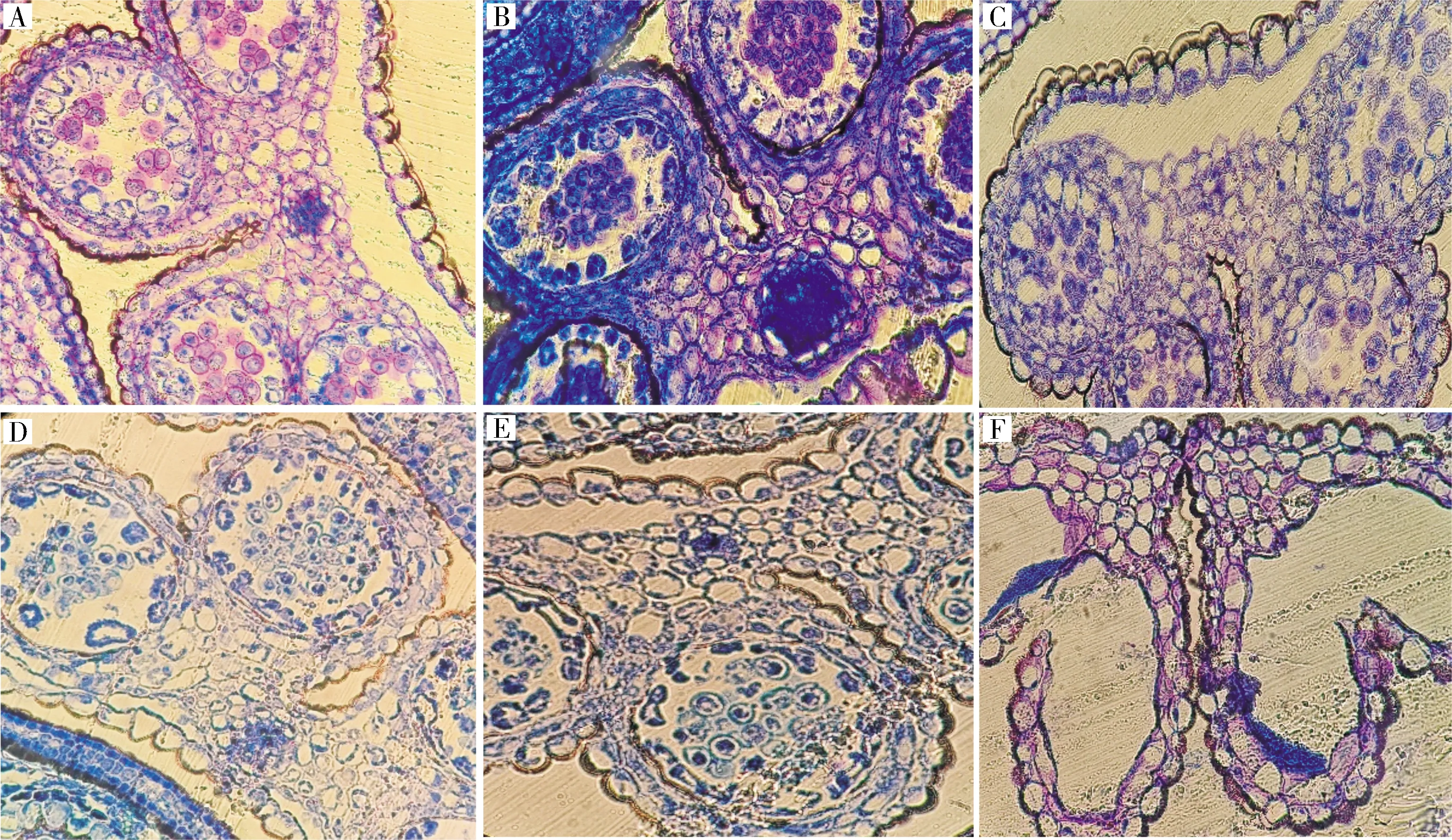
2)芥菜hau细胞质雄性不育系材料花器官发育及花药败育观察。将果荚正常和果荚畸形的芥菜hau细胞质雄性不育系和保持系的花器官进行对比(图5),可以发现,2类材料的雄蕊败育基本都由绒毡层的异常发育所引起;而结荚畸形材料,则直接形成了雌蕊-雄蕊嵌合体,嵌合体内生长着多于正常数量的胚珠。对结荚正常材料雄蕊进行败育观察(图6),可以看到,孢原细胞时期,产生心皮组织;花粉母细胞时期,绒毡层细胞异常膨大;在四分体时期,绒毡层发育异常,小孢子因养分不足随之降解,花药败育。对结荚畸形材料的败育过程进一步观察(图7),可以看到,雌蕊-雄蕊嵌合体在花药发育初期就开始形成,此时产生心皮组织而非正常的雄蕊原基,之后,心皮组织不断生长发育,形态各异,整个过程几乎观察不到正常的花器官。
A.正常荚果不育系; B.畸形荚果不育系; C.保持系。A.Normal pods with hau CMS; B.Abnormal pods with hau CMS; C.Maintainer line.
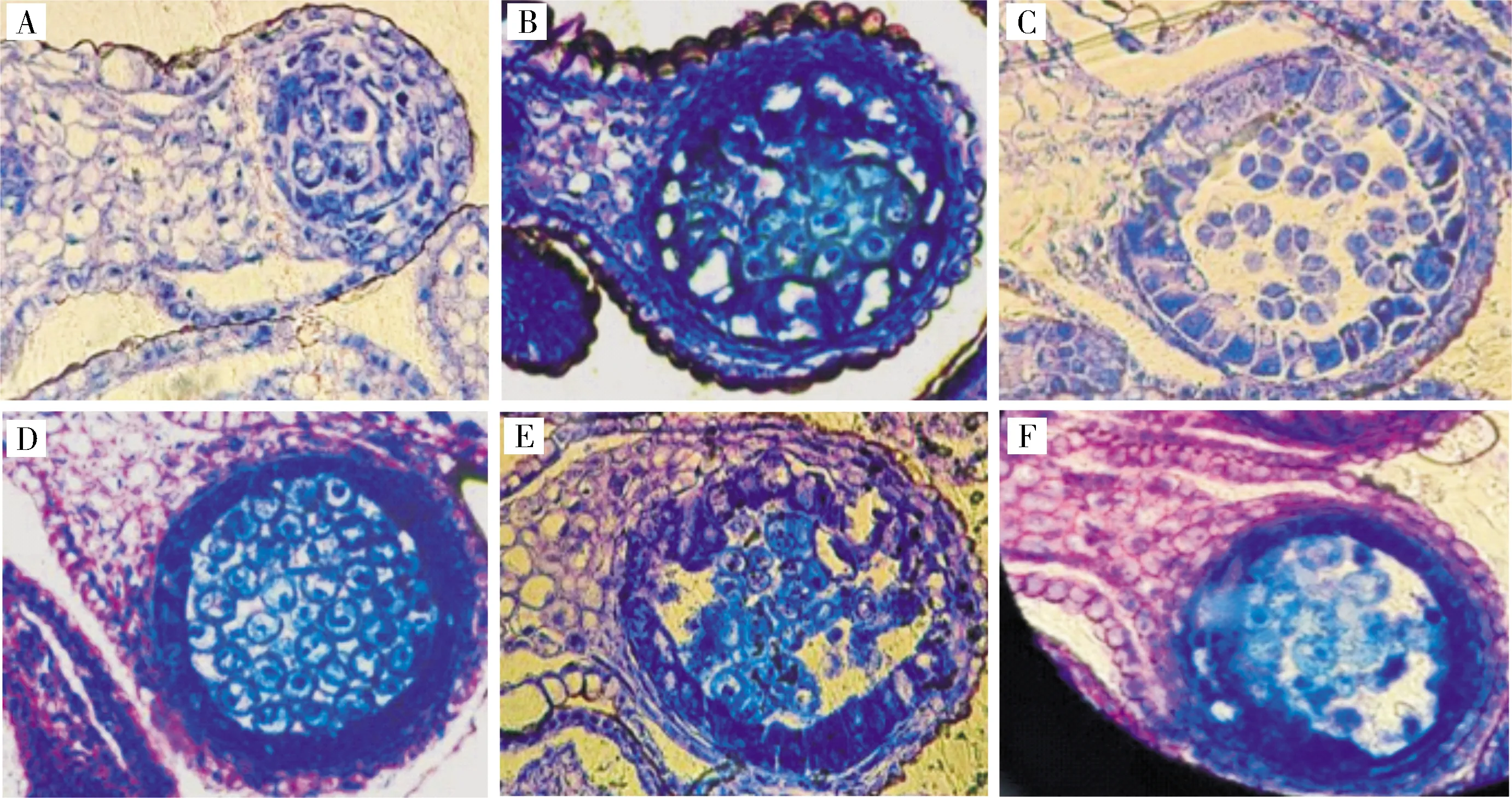
A.孢原细胞时期; B.花粉母细胞时期; C.四分体时期; D-F.配子体发育时期。A.Sporogenous cell stage; B.Pollen mother cell stage; C.Tetrad stage; D-F.Gametophyte development stage.
3 讨 论
在植物新品种的选育过程中,为避免长期使用单一不育胞质所带来的潜在危害,需要引进不同类型的胞质并且对其快速鉴定,而分子标记则具有准确、省时、高效的特点[18];本研究利用2对特异性引物(orf138和orf288)对芥菜CMS材料的不育胞质类型进行快速鉴定,发现orf138标记可鉴定芥菜oguCMS,而orf288标记则可将芥菜hauCMS鉴定出来,相较于观察花器官的方式简单且快速。从59份材料结荚性状的田间统计结果中可以发现,所有的保持系和oguCMS结荚表现均非常优异,无畸形情况发生,这与邹瑞昌[19]的研究结果相似。芥菜hauCMS是华中农业大学1999年所发现的天然芥菜型油菜雄性不育系,具有不育度和不育株率100%的特点,长期以来,在十字花科植物杂种优势利用中得到了大量的应用[3]。施展等[20]对芥菜hauCMS 进行了综合遗传分类后,将芥菜hauCMS的不育细胞质转育到大白菜中,创制了大白菜hauCMS种质资源6w-9605A。张艳等[21]以甘蓝型油菜hauCMS为母本,菜心为父本,进行种间杂交和连续回交,创制了新型菜心hauCMS不育材料。随着对芥菜hauCMS不育资源利用和研究的不断深入,笔者课题组也发现其不足之处,将芥菜hauCMS不育源转入芥菜后,部分材料存在果荚畸形的情况,实际种子生产中,果荚畸形会严重影响种子品质甚至制种产量,部分芥菜hauCMS材料结荚畸形可能会影响其后续在十字花科作物中的推广利用,因此,对芥菜hauCMS不育胞质进行遗传改良显得尤为重要。
本研究进一步对芥菜hauCMS和oguCMS材料进行花药败育的细胞学观察,可以发现,芥菜oguCMS和果荚正常的hauCMS材料,雄蕊的败育均由绒毡层的异常发育所引起,其雌蕊进行正常的授粉受精后,果荚仍能正常发育,这与单奇伟等[22]的研究结果类似;果荚畸形的芥菜hauCMS材料在其花器官发育过程中,除绒毡层发育异常外,还伴随着雌蕊-雄蕊嵌合体的发生,最终导致果荚畸形,可见,雌蕊-雄蕊嵌合体的形成是造成果荚畸形的原因,这与文献[23]的研究结果不一致,极有可能是核质互作效应引起的[24]。本研究通过细胞学水平的研究证实结荚畸形由雌蕊-雄蕊嵌合体的生成引起,但更深层次的机制尚待探究,根据花发育模型假说,B基因组是调控雄蕊发育、引起雄蕊心皮化形成嵌合体的关键[25],因此,后续可以从B基因组着手,对芥菜hauCMS进行深入研究,探究其败育的分子机制。
