基于液质联用技术对早期高脂血症金黄地鼠肝脏鞘脂的靶向代谢组学研究*
2021-06-07孙明谦杨会珍刘建勋
孙明谦 苗 兰 张 颖 彭 勍 杨会珍 刘建勋 林 力
(中国中医科学院西苑医院基础医学研究所,中药药理北京市重点实验室,北京 100091)
动脉粥样硬化是一种由多种因素引起的慢性炎症性疾病,其内皮损伤与脂质过氧化有关[1-2]。引发和加速动脉粥样硬化的一个主要原因是高脂血症导致氧化应激和炎症的增加,高脂血症是目前诱发心脑血管疾病的重要因素之一[3]。肝脏通过从血清中清除胆固醇和甘油三酯,在脂质代谢中发挥重要作用。因此,对肝脏代谢变化的描述对于高脂血症的分子机制研究具有重要意义[4]。
鞘脂(sphingolipids)一类含有鞘氨醇骨架的两性脂,是生物膜中的一类主要脂类,也是细胞信号传递中的一种生物活性脂类介质[5]。它能调节细胞的生存、增殖、凋亡、分化、迁移和免疫反应等重要生理过程。越来越多的研究表明,鞘脂在动脉粥样硬化斑块的形成和发展中起着重要作用,并导致心血管疾病[6-7]。 在高脂血症的情况下,肝脏经常遭受严重的脂肪堆积、炎症和氧化应激的损害。这些损伤与鞘脂中神经鞘磷脂(sphingomyelin,SM)-神经酰胺(ceremide,Cer)-鞘糖脂(glycosphingolipids)之间的代谢密切相关,这一代谢途径在鞘脂类成分的代谢中占有重要地位,它们之间的相互代谢也与多种疾病的发生发展密切相关[8]。神经酰胺占据了这一途径的中心枢纽,它们是一个紧密的系统,彼此密切相关和相互作用。因此,研究这一途径在动脉粥样硬化早期发病机制中的变化具有重要意义[9]。本研究主要针对鞘脂中的神经鞘磷脂、神经酰胺及葡萄糖神经酰胺(glucosyl-ceramide,Glc-cer)建立靶向代谢组学方法,围绕这一代谢途径进行系统研究,以期为动脉粥样硬化的治疗找到新的治疗靶点。
1 材料和方法
1.1 仪器与试剂
UltiMateTM3000超快速液相色谱、Q ExactiveTM四极杆轨道离子阱质谱、Mikro 220R真空离心浓缩仪(ThermoFisher Scientific);Mix-3000震荡混匀器(杭州米欧仪器有限公司); LAbOSPEC T003日立全自动生化仪、TG、TC、LDL-C 及 HDL-C测定试剂盒(HITACHI);台式高速冷冻离心机(Hettich); BX51显微镜(Olympus)。
高脂饲料(10 kg/箱),A级,购自北京科澳协力饲料有限公司;乙腈、异丙醇、甲醇、水和甲酸为高效液相色谱级,购自Fisher Scientific;醋酸铵购自Sigma-Aldrich。
1.2 动物实验
18只SPF级叙利亚金黄地鼠,雄性,7周龄,购自北京维通利华实验动物技术有限公司,实验动物生产许可证号:SCXK(京)2016-0011,合格证编号11400700209462。金黄地鼠适应性饲养一周后分组,分为正常对照组(n=9)和高脂血症模型组(n=9)。高脂血症模型组饲喂高脂饲料6周,正常对照组饲喂常规饲料。实验结束时,所有金黄地鼠均隔夜禁食不禁水。采血后将肝脏用生理盐水冲洗,一部分肝脏取出后立即注入液氮,储存在-80 ℃;另一部分肝脏进行固定、HE染色后观察组织学变化。血液样本经4 ℃、12 000 r/min,离心10 min,按血液生化指标试剂盒的操作说明进行相关生化指标检测。
1.3 样品处理与液质条件
1.3.1样本处理:称取适量组织肝脏,研磨后,加1 mL甲醇混匀。混匀震荡30 min,然后在4 ℃ 12 000 r/min离心10 min,取上层溶液100 μL,置于200 μL内衬管中,待测。
1.3.2液相条件:流动相为A(乙腈∶水=6∶4,0.1%甲酸,10 mmol/L乙酸铵)和B(异丙醇∶乙腈=9∶1,0.1%甲酸,10 mmol/L乙酸铵);洗脱程序: 20%B;1 min,20%B;11 min,100%B;13 min,100%B;16 min,20%B;停止色谱柱: Waters UPLC HSS T3(1.8 μm,2.1 mm×100 mm);流速:0.3 mL/min;进样量:1.0 μL;柱温:45 ℃。
1.3.3质谱条件:正离子离子源电压分别为3.7 kV;毛细管加热温度320 ℃;翘气压力30 psi,辅助气压力10 psi;容积加热蒸发温度300 ℃;翘气和辅助气均为氮气;碰撞气为氮气,压力为1.5 m Torr;一级全扫描参数为:70 000 dpi,自动增益控制目标为1×106,最大隔离时间50 ms,质荷比扫描范围50~1 500;液质系统由Xcalibur 2.2 SP1.48软件控制,数据采集和靶向代谢物定量处理均由该软件操作。
1.4 数据采集与分析
根据已建立方法,将原始LC-MS数据导入Skyline软件分析39种靶向鞘脂。在建立的极性脂质数据库中,根据保留时间和准确质量对鞘脂种类进行相对定量。使用SIMCA-P 12.0软件进行多变量分析[10]。
2 结果
2.1 血清参数比较
结果见表1所列。高脂血症模型组TG、TC、LDL-C,HDL-C水平明显高于正常对照组,均具有统计学差异(P<0.01)。

表1 金黄地鼠血清中TG、TC、LDL-C 及 HDL-C 水平Table 1 Level of serum TG,TC,LDL-C and HDL-C in hamsters
2.2 组织病理学
如图1所示,正常对照组肝组织结构正常,肝细胞索围绕中央静脉呈放射状分布,排列整齐有序,肝窦充盈、可见,肝细胞未见肿胀变性,汇管区未见炎细胞浸润及纤维组织增生。
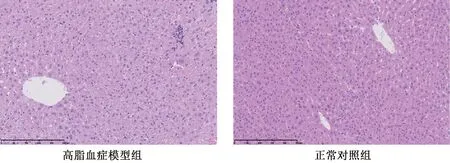
图1 HE 染色的病理图(×200)Fig.1 Micrographs of hyperlipidemia hamster(×200)
高脂血症模型组肝组织结构紊乱,肝细胞大片气球样变性,灶性小泡性脂肪变,肝细胞肥大致肝窦不明显,枯否细胞增生,肝小叶内可见炎细胞聚集灶,汇管区轻度炎细胞浸润及纤维组织增生。
2.3 多元数据分析
图2显示了肝匀浆液靶向代谢组研究的PCA和PLS-DA评分图。结果表明,与正常对照组比较,高脂血症模型组肝脏鞘脂代谢变化更为显著。数据模型显示区分效果良好,相关参数:对于PLS-DA模式(R2Y=0.993,Q2=0.939),PCA(R2X=0.866,Q2=0.729)。VIP值反映了各变量(代谢物)对分类的影响,经PLS-DA处理后生成。从t检验中筛选出VIP>1、fold变化>1.5、P<0.05的潜在生物标志物进行进一步研究。共鉴定出21种潜在的鞘脂生物标志物,包括8种SM、4种Glu-cer和9种Cer。潜在生物标志物变化趋势见图3。

图2 基于LC-MS的PCA和PLS-DA评分图Fig.2 PCA and PLS-DA score plots based on data from LC-MS

图3 肝脏鞘脂热图Fig.3 Heatmap of the sphingolipids in liver
3 讨论
越来越多的证据表明,鞘脂代谢途径的主要成分在心血管疾病、肥胖症、糖尿病、胰岛素抵抗和非酒精性脂肪肝疾病中都起着重要作用[11-13]。 鞘脂中各类成分之间的相互代谢都发挥着重要的生物学功能,如鞘磷脂与神经酰胺之间的代谢被激活会发挥促炎症、促氧化和细胞死亡诱导活性,导致动脉粥样硬化,衰老和心血管事件。 此外,近年来也发现鞘磷脂-神经酰胺-鞘糖脂通路中的成分作为潜在的次级信使分子直接或间接影响细胞和亚细胞器,在胰岛素抵抗和脂肪肝的病理状态中发挥着重要作用,可能是新的治疗靶点[14]。本研究发现高脂血症患者肝脏中4种Glc-cer和5种鞘磷脂呈下降趋势,另外3种鞘磷脂和9种神经酰胺均呈上升趋势,提示鞘脂代谢的整体异常。
神经酰胺在各种鞘脂的合成和分解代谢中起着关键作用。在鞘磷脂-神经酰胺-鞘糖脂途径中,神经酰胺的主要是通过鞘磷脂酶水解神经鞘磷脂而生成[15],而葡萄糖神经酰胺和半乳糖神经酰胺通过葡萄糖基转移酶的分解生成神经酰胺[16]。神经酰胺被认为是胰岛素抵抗和不同器官炎症的重要脂质介质。许多研究表明神经酰胺与胰岛素敏感性呈负相关,与炎症呈正相关,肥胖的啮齿动物和人类的血清和肝脏中神经酰胺的含量都会升高[17-18],血脂异常和炎症是非酒精性脂肪肝和动脉粥样硬化发病的共同关键因素。肝代谢异常不仅导致脂肪肝脂肪变性的发生,而且增加了动脉粥样硬化的风险。此外,升高的肝神经酰胺可能导致循环神经酰胺水平升高,从而通过直接损伤血管内皮促进动脉粥样硬化[19]。
鞘磷脂是脂蛋白中含量最丰富的鞘脂,约占血浆总鞘脂的87%,占血浆总磷脂的20%。鞘磷脂由神经酰胺通过鞘磷脂合酶产生[20]。和神经酰胺一样,鞘磷脂也是心血管疾病的危险因素,据报道,主动脉中高浓度的鞘磷脂可能有害,致动脉粥样硬化的脂蛋白,如VLDL和LDL都是富含鞘磷脂的[21]。高脂血症金黄地鼠肝脏中鞘磷脂水平呈现不同的变化趋势,大部分鞘磷脂呈显著的减少趋势,这也可能表明鞘脂代谢的被激活的方向是产生神经酰胺,而不是生成鞘磷脂。但由于SM的组成复杂,含量高,其代谢异常的机制有待进一步研究。葡糖神经酰胺是由葡糖神经酰胺合酶(GCS)引起的神经酰胺糖基化反应形成的,GCS将葡萄糖与神经酰胺的1-羟基连接起来。葡糖神经酰胺与炎症、糖尿病和动脉粥样硬化密切相关[22]。本研究中Glc-cer的降低可能表明神经酰胺的糖基化作用降低,导致神经酰胺的升高[19]。
从文献中可以看出,这三种成分对肝脏和心血管系统都有不良影响。然而,在高脂血症的早期,鞘脂的变化并不是都升高,其代谢途径主要还是朝着激活生成神经酰胺的方向变化[8]。神经酰胺在鞘脂的代谢过程和生物功能方面,都发挥着中心作用[23]。根据我们的研究,在高脂血症的早期,神经酰胺和鞘磷脂或葡萄糖神经酰胺之间的肝脏平衡被打破,可能导致神经酰胺升高的原因之一,脂质代谢的异常,最先导致神经酰胺的升高,其可能引起肝功能异常,从而导致包括鞘脂在内的脂质代谢的进一步紊乱。
本研究采用液质联用技术,对高脂血症金黄地鼠肝组织中的鞘脂类成分进行了靶向代谢组学研究。21种与动脉粥样硬化高度相关的鞘磷脂、神经酰胺和葡糖基神经酰胺在高脂血症仓鼠肝脏出现异常改变。其中,只有神经酰胺呈现较为明显上升趋势,表明鞘脂的代谢途径中被激活的生成是神经酰胺的途径,提示神经酰胺的增加可能是仓鼠动脉粥样硬化早期肝脏病变的主要特征。这种增加可能进一步导致鞘脂代谢功能障碍,甚至导致脂质损伤肝脏或内皮细胞。研究也表明,基于LC-MS的靶向代谢组学是进一步了解动脉粥样硬化早期脂质功能障碍机制的有效工具。
