10种鲤形目鱼类ZP2基因的克隆及特征分析
2021-05-28陈沙刘焕章曹文宣俞丹
陈沙,刘焕章,曹文宣,俞丹*
(1.大连海洋大学,辽宁大连116023;2.中国科学院水生生物研究所,水生生物多样性与保护重点实验室,武汉430072)
透明带(zona pellucida,ZP)是包围在卵母细胞或早期胚胎外的一层糖蛋白,通常被称为透明带蛋白(ZP蛋白),通常由3、4种糖蛋白组成,受ZP基因家族编码表达(Goudetetal.,2008)。ZP基因中除ZP结构域外,通常还包含N端信号肽序列、弗林蛋白酶切割位点、跨膜结构域、三叶草结构域或EGF结构域(Wassarman &Litscher,2004)。硬骨鱼类基因组经历过特有的基因组复制,因此基因组上存在着大量的ZP蛋白编码基因,被复制的基因在鱼类中可能演化出新的功能,比如南极鱼Patagonotothen ramsayi的ZP蛋白被证实是抗冻蛋白(马战领,2015;Caoetal.,2016)。除了抗冻功能,ZP蛋白在鱼类中是否还有其他功能尚不清楚。
ZP2基因是ZP基因家族的重要成员,除了含有保守的三叶草结构域和ZP结构域外,还有1个富含脯氨酸(P)和谷氨酰胺(Q)残基的N端重复结构域(Wang &Gong,1999)。Lyons等(1993)克隆获得了美洲拟鲽Pseudopleuronectes americanus的ZP2基因序列,发现其重复结构域的重复单元为(PQQPQQPQQPQQPKY)7。随后,发现鲤Cyprinus carpio的重复单元为(QQTSQQFQPQKPA/V)13(Changetal.,1997);稀有鲫Gobiocypris rarus的重复单元为(VQQQASQQFPQR/Q)(12~19)(Wuetal.,2012);鲫Carassius auratus的重复单元为(QQPKQFPF/LQKPVP)7(史建伍,2014)。Wang和Gong(1999)首次在斑马鱼Daniorerio上克隆到了ZP2基因序列,但此基因没有含P和Q的重复结构域。同样,在青鳉Oryzias latipes、日本鳗鲡Anguilla japonica中也没有发现重复结构域(Kanamorietal.,2003;Sanoetal.,2010)。已有研究表明,ZP2基因重复结构域的重复序列在受精后的谷氨酰胺的γ-羰基和赖氨酸的ε-氨基之间会形成分子间交联,从而参与鱼卵卵膜的硬化(Oppen-Berntsenetal.,1990;Giaccoetal.,2000)。但这些串联重复单元是否具有其他功能尚不清楚,例如是否与鱼类的产卵习性相关。
鲤形目Cypriniformes鱼类约4 200种,是世界上最大的淡水鱼类类群(Nelsonetal.,2016),广泛分布在除南极洲、南美洲和澳大利亚以外的世界各大洲。鲤形目鱼类起源于193 Ma的侏罗纪(Taoetal.,2019),并演化出了丰富多样的产卵习性。例如,为适应东亚季风气候的特点,鲤形目鱼类演化出了产漂流性卵习性的类群,并成为了河流、湖泊的主体和特色类群,该类群在水位连续上涨、水流刺激时产卵,卵随水漂流,孵化发育;相反,产粘性卵习性的类群繁殖喜欢稳定的环境,受精卵粘附于植物等基质上固着发育,水位上涨是产卵的刺激因素,多在近岸浅水处产卵;而产沉性卵习性类群的受精卵通常沉于水底发育,多在江河上游水质清澈、砂砾地质处产卵(解涵等,2010)。这种繁殖产卵方式的分化,对于鱼类的物种多样性的形成具有重要作用,但其相关的分子机制尚缺乏研究。Mansour等(2009)研究发现,糖蛋白是鲤卵表面具有粘性的主要原因,而编码这类糖蛋白的正是ZP基因家族。因此,本研究将以鲤形目不同产卵习性的代表性鱼类为研究对象,利用RT-PCR技术克隆ZP2基因,探讨ZP2基因重复结构域的变异与鲤形目鱼类产卵习性的相关性。
1 材料与方法
1.1 实验材料
本研究的克隆对象包含10种鲤形目鱼类,其中,产漂流性卵7种、产粘性或沉性卵3种(表1)。此外,在GenBank上下载其他4种鲤形目鱼类、美洲拟鲽和青鳉的ZP2基因作为外类群。
表1 鲤形目鱼类的产卵习性及ZP2基因序列来源Table 1 Spawning types of Cypriniformes and GenBank accession numbers of ZP2 gene
1.2 总RNA提取和第一链cDNA合成
按照动物组织RNA提取试剂盒操作流程提取卵巢和肝脏的总RNA。提取的总RNA直接进行逆转录反应。反应体积为20 μL:总RNA为2 μL,Oligo(dt)18 Primer试剂1 μL和Nuclease-free水 9 μL,65 ℃ 5 min,然后向反应产物加5×Reaction Buffer 4 μL、Ribolok RNase Inhibitor 1 μL、10 mmol·L-1dNTP Mix 2 μL和RevertAid Reverse Transcriptase 1 μL。最后在PCR仪中进行42 ℃ 60 min,25 ℃ 5 min,70 ℃ 5 min。
1.3 ZP2基因克隆
根据GenBank中已发表的其他鱼类的ZP2基因序列设计了6对同源PCR引物(表2),克隆10种鲤形目鱼类的ZP2基因的cDNA序列。反应体积为50 μL:16 μL Nuclease-free水,25 μL Buffer,1 μL dNTP,正、反引物各2 μL,1 μL Super Fidelity DNA Polymerase和3 μL cDNA。扩增程序:95 ℃ 3 min;95 ℃ 15 s,60 ℃ 15 s,72 ℃ 1 min,共35个循环;72 ℃ 5 min。产物经1%琼脂糖凝胶检测后,目的片段送生工生物工程(上海)股份有限公司克隆测序。

表2 本研究相关的引物Table 2 Primers used in this study
1.4 ZP2基因生物信息学分析
将克隆获得的cDNA序列在NCBI作BLAST比对分析,通过在线软件ExPASy推测不同ZP2基因的开放阅读框,使用在线软件EMBOSS explorer预测氨基酸序列。利用ExPASy分析ZP2基因的相对分子质量、等电点等特征,使用在线软件SMART分析氨基酸序列结构,并用DNAMAN对得到的序列进行同源性分析。采用在线工具SOPMA分析ZP2蛋白的二级结构及各成分的占比。通过Phyre2同源建模预测ZP2的三维结构模型。利用MEGA 7.0采用邻接法(Neighbor-Joining)构建鱼类ZP2基因的系统发育树。最后,通过EasyCodeML检测ZP2基因的受选择压力情况,用非同义替代率(nonsynonymous substitution rate,dN)与同义替代率(synonymous substitution rate,dS)的比值ω(ω=dN/dS)估算选择压力(Gaoetal.,2019)。
2 结果与分析
2.1 ZP2基因序列分析
根据RT-PCR方法,克隆扩增获得了10种鲤形目鱼类的ZP2基因的cDNA全长序列。同时获得了完整的开放阅读框,序列全长1 254~1 734 bp,编码了417~577个氨基酸,相对分子量为45.556~63.729 kDa,理论等电点为4.78~5.86(表3)。

表3 ZP2基因的序列信息Table 3 Sequence information of ZP2 gene
2.2 ZP2基因结构分析
10种鲤形目鱼类的ZP2基因均含有N端信号肽序列、三叶草结构域、ZP结构域和弗林蛋白酶切割位点,但都不含跨膜结构域(图1)。产粘性卵鱼类的重复结构域中含有重复次数较多的重复单元(7~19次),如团头鲂(15次)、稀有鲫(12~19次),每个物种的重复单元序列存在差异(图1;表4)。产漂流性卵和产沉性卵鱼类不含重复结构域或重复单元重复次数较少(2~5次),如:翘嘴鲌、银、光唇蛇、鳊、银鲴和宽鳍重复单元重复次数较少;犁头鳅、似鳊和蒙古鲌不含重复结构域。

表4 ZP2基因重复结构域的重复单元及重复次数Table 4 Repeat unit and its number in the repeat domain of ZP2 gene
2.3 ZP2氨基酸序列同源性分析及系统发育关系分析
根据克隆获得的ZP2基因氨基酸序列,分析鲤形目鱼类的ZP2基因的同源性(图2)。所有物种的ZP2氨基酸序列包含了18个保守的半胱氨酸残基(6个位于三叶草结构域中、12个位于ZP结构域中)。ZP2氨基酸保守区域集中在N端信号肽序列、三叶草结构域和ZP结构域(图2)。团头鲂与鳊的相似性最高(99.7%),斑马鱼和鲫的相似性最低(71.2%)。以美洲拟鲽和青鳉作为外类群,在系统发育树上,所有鲤形目鱼类的ZP2氨基酸序列构成一个单系(图3),其中斑马鱼单独为一支,位于系统树的基部位置,与其他鲤形目鱼类形成姐妹群。
2.4 ZP2蛋白二级结构分析
通过SPOMA对不同鲤形目鱼类的ZP2蛋白进行二级结构预测,结果表明:二级结构主要由α-螺旋(10.58%~27.38%)、β-转角(2.05%~3.60%)、延伸链(18.37%~26.79%)和无规则卷曲(49.61%~63.49%)组成,不同物种间的各成分所占比例各不相同(表5),不同物种的ZP2蛋白二级结构具有一定的差异(图4)。
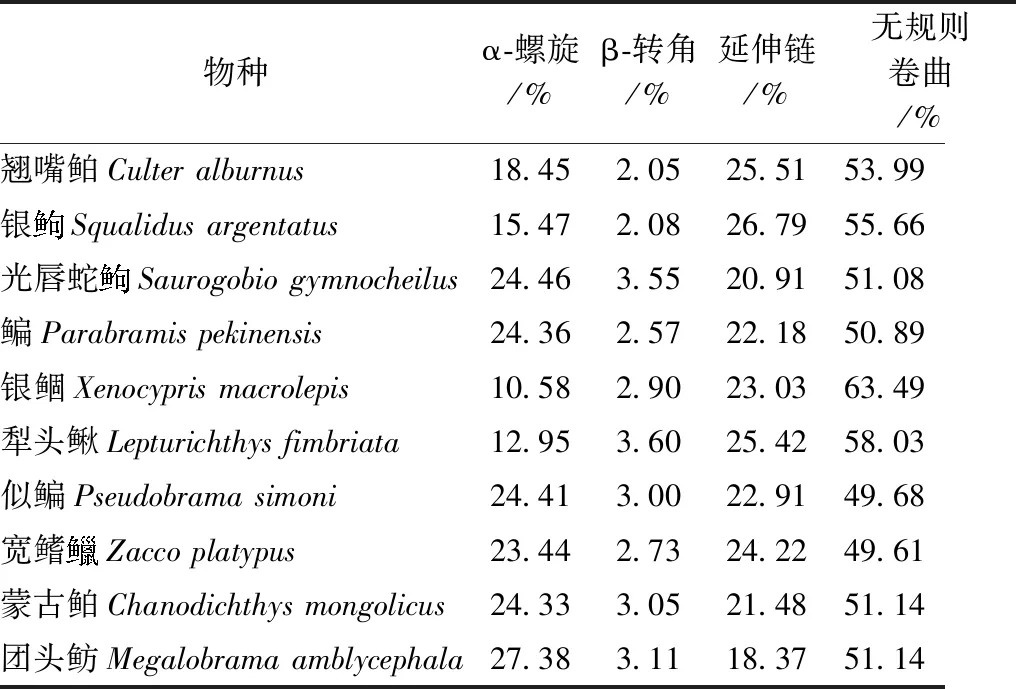
表5 10种鲤形目鱼类的ZP2蛋白二级结构相关参数Table 5 Secondary structure parameters of ZP2 protein of 10 Cypriniformes species
2.5 ZP2蛋白三维结构预测分析
通过Phyre2同源建模得到了鲤形目鱼类ZP2蛋白的三维结构模型,不同物种的置信度均为100%,均显示最大程度与数据库中的小鼠MusmusculusZP2晶体结构模板匹配,预测模型与模板匹配相似度为38%~41%。构建三维模型时,不同物种的入模氨基酸范围和数目存在差异,且入模的氨基酸也存在变异位点(表6),进而导致不同物种的三维结构存在细微的差异(图5)。

表6 10种鲤形目鱼类的ZP2蛋白三维结构建模相关参数Table 6 Three-dimensional structural parameters of ZP2 protein in 10 Cypriniformes species
2.6 ZP2基因选择压力分析
利用位点模型的M1a和M2a、M7和M8,分支模型的two-ration和one-ration,分支-位点模型的model A和model A-null对鱼类的ZP2基因进行正向压力选择分析(表7)。结果表明,M0模型中,所有位点的平均ω为0.21,鱼类的ZP2基因整体受到纯化选择的影响;分支模型(two-ration)和M0模型中,似然比检验差异显著(P<0.05),说明ZP2基因的氨基酸位点经历了不同的选择压力;M1a和M2a似然比检验差异不显著(P>0.05),认为有79%的位点受到纯化选择(ω=0.11),而其他位点处于中性进化状态;M7和M8似然比检验差异显著(P<0.05),表明大部分位点受到正向选择(ω=1.91),位点的概率经过BEB检验,总共检验出 9个后验概率大于0.5的正选择氨基酸位点(8 V 0.864、19 L 0.648、43 P 0.835、97 R 0.813、132 G 0.553、258 R 0.612、320 A 0.600、344 T 0.787、350 V 0.950),其中第350位的氨基酸位点(350 V)在5%的置信水平显著,后验概率为95%;在分支-位点模型的检测中,将鲤形目鱼类指定为前景分支,似然比检验在备择假设和零假设中没有显著差异(P>0.05),没有检验到正向选择位点。

表7 EasyCodeML计算的各个模型的似然值及参数Table 7 Likelihood values and parameters of each model calculated by EasyCodeML
3 讨论
3.1 鲤形目代表性鱼类的ZP2基因结构特性
本研究发现10种鲤形目鱼类的ZP2基因结构均符合ZP基因家族的特征,含有保守的N端信号肽序列、三叶草结构域、ZP结构域和弗林蛋白酶切割位点。ZP2基因是ZP基因家族的重要成员之一,它含有许多参与ZP蛋白分泌和组装的保守功能域。其中,N端信号肽序列可以使正在翻译的核糖体附着到内质网膜上合成并分泌蛋白(Blobel,1975);三叶草结构域的功能在于抗蛋白的水解消化(Bork,1993);ZP结构域是ZP蛋白的主要功能结构域,由大约260个氨基酸残基组成,其中ZP2基因的ZP结构域有10个保守的半胱氨酸残基,半胱氨酸残基之间的二硫键对ZP结构域的立体折叠和蛋白之间的相互作用有着决定性的作用,起着聚合作用,可以参与ZP蛋白聚合成高阶结构,如纤维和基质,此外,ZP结构域通常包含着ZP-N和ZP-C结构合成纤维,ZP-C总是与ZP-N相关联,并且可能具有其他重要的功能,如调节不同亚基之间特异性的相互作用;弗林蛋白酶切割位点主要参与ZP蛋白的分泌,在哺乳动物中,它可以启动蛋白质的聚合和组装(Bork &Sander,1992;Jovineetal.,2002,2004,2005,2006;Sasanamietal.,2006;Kanaietal.,2010)。本研究克隆获得的10种鲤形目鱼类的ZP2基因均含有上述4种结构域。此外,在本研究中发现10种鲤形目鱼类均不含跨膜结构域,有研究认为鱼类的ZP蛋白最初是具有跨膜结构域的,但在进化过程中丢失了,但所有哺乳动物的ZP蛋白均包含1个跨膜结构域,该结构域的作用是锚定ZP蛋白到细胞膜上进行组装(Wassarman &Litscher,2004;Sanoetal.,2010)。ZP2基因结构在哺乳动物和鱼类中的差异暗示着ZP蛋白在哺乳动物和鱼类中的功能存在不同。
3.2 鲤形目代表性鱼类ZP2基因的重复结构域
分析目前所有已获得的鲤形目鱼类ZP2基因序列,发现不同物种间存在较大的序列长度差异(1 254~1 734 bp),该差异主要来源于重复结构域的有无及重复单元和重复次数的不同,即是否存在串联重复序列或串联重复序列的长度是否存在差异,由此导致鲤形目鱼类的ZP2蛋白二级结构组分及其空间结构存在一定的差异。基因的重复结构域研究受到越来越多的关注。Charlesworth等(1994)已经证明重复序列可直接参与重组。当重组发生在重复结构域的不同位置时,姐妹染色单体之间或同源染色体之间会发生不均匀的交换,从而导致重复结构域的延长和缩短(Changetal.,1997)。但不是所有物种都有重复结构域,缺少重复结构域可能是该结构域的二次缺失,相反,重复结构域也可能发生平行或趋同进化,明显不一致的重复结构域的存在表明它们的快速进化,重复单元可以通过DNA的复制或连续不均匀交叉而形成或消除,但是重复结构域的作用与功能需要进一步研究(Lewin,2000;Kanamorietal.,2003)。重复序列能参与遗传物质结构的改变、基因调控以及细胞分化等过程,有的还可以直接编码蛋白质(王剑波,蒋左庶,1992)。一般来说,具有重复序列的基因容易发生缺失,缺失是基因组进化差异的主要驱动力之一,因为重复序列的缺失可能会导致相应蛋白质的三维结构发生变异,Guo等(2012)通过对HOMOLENS数据库中斑马鱼、三刺鱼Gasterosteus aculeatus、青鳉、红鳍东方鲀Takifugu rubripes、绿河鲀Tetraodon nigroviridis的分析,发现重复序列的缺失对蛋白质的三维结构有着很大的影响;也可能通过改变翻译阅读框来诱导蛋白质的功能发生变化(Raes &Peer,2005)。鱼卵粘性的产生是一个缓慢、持续时间最少为5 min的生理过程,皮层小泡释放的糖蛋白(主要由ZP基因家族编码)参与了卵粘性的生成(Chang &Huang,2002;Gallianoetal.,2003;Mansouretal.,2009)。此外,卵子在受精后,卵膜随后会发生硬化,有研究认为卵膜的硬化可分为2步:首先金属蛋白酶水解ZP2的N端脯氨酸和谷氨酰胺(Pro-Gln)区域,其次谷氨酰胺转移酶会催化水解ZP2和ZP3发生分子交联(Sugiyama &Iuchi,2000;Shibataetal.,2012)。Wu等(2012)也认为重复单元的重复次数可以影响硬骨鱼卵膜的硬度,重复单元的重复次数越多,卵膜就越坚硬。本研究通过比较鲤形目代表性鱼类重复结构域中重复单元的重复次数,发现其重复结构域与其产卵习性存在一定程度的相关性,例如产漂流性卵(7种)和产沉性卵鱼类(3种)不存在重复结构域或重复单元的重复次数较少(2~5次),而产粘性卵鱼类(4种)的重复单元的重复次数较多(7~19次),推测产漂流性卵和产沉性卵鱼类中ZP2基因重复结构域的缺失影响了鱼卵粘性的形成及卵膜硬化。鲤形目中,产漂流性卵鱼类有50余种,本研究获得了7种典型的代表性鱼类,包括翘嘴鲌(鲌亚科Culterinae)、鳊(鲌亚科)、银鲴(鲴亚科Xenocyprininae)、似鳊(鲴亚科)、银(亚科Gobioninae)、光唇蛇(亚科)和犁头鳅(平鳍鳅科Balitoridae),约占产漂流性卵类群的14%,但产沉性卵和粘性卵的物种数较少,因此不同物种重复结构域的差异是否引起了ZP2蛋白空间结构的差异并导致了产卵习性的不同还需要进一步功能实验的证实。
3.3 鲤形目鱼类ZP2基因的选择压力
突变、遗传变异、选择和基因流是自然生物进化的主要动力(沈银柱,黄占景,2012)。在选择压力检验中,非同义突变与同义突变的比值(ω=dN/dS)被用于衡量基因或者其中个别位点是否受到选择压力(Yang &Nielsen,1998)。不导致氨基酸改变的核苷酸变异被称为同义突变,反之为非同义突变。一般认为,同义突变不受自然选择,而非同义突变则受到自然选择作用。在长期进化过程中,选择压力不可能均匀地作用于1个基因,根据其功能作用,它更有可能在某些特定位点发生(Kamath &Getz,2011)。在硬骨鱼中,有关卵膜和精子相互作用的证据很少,ZP蛋白在精卵结合中可能没有起到作用,因为精子只能通过珠孔进入卵子,卵膜中任何与精子结合的分子都会聚集在珠孔周围,但是没有迹象表明ZP蛋白在鱼卵的珠孔周围,鱼类的ZP蛋白可能只起到了结构性作用,如保护功能(Conner &Huang,2003)。ZP蛋白家族通常包含大量的ZP基因(Xuetal.,2012)。ZP基因家族在硬骨鱼类中产生了不同程度的扩张,如斑马鱼中可分离出21个ZP基因,有研究认为,斑马鱼ZP基因的大量扩张可能与其需要大量产卵有关(Conner &Hughes,2003);雅罗鱼Leuciscus waleckii基因组可鉴定出 30个ZP基因,ZP基因的扩张增强了雅罗鱼在碱性环境中胚胎发育的保护能力,这些扩张均被正向选择驱动(Xuetal.,2017;吴天利,2018)。此外,南极鱼的ZP基因在寒冷条件下大量扩张,有助于丰富ZP蛋白的转录本和增加卵膜的厚度,从而被认为是防止冰渗透的关键物理屏障,使南极鱼可以适应极端的低温环境(Chenetal.,2008;马战领,2015)。但是,胎生月光鱼Xiphophorus maculatus的ZP基因家族在正选择下相对减少,因为这些ZP蛋白对胚胎的保护不再是必要因素,而是进化出了气体和物质交换的机制(Schartletal.,2013)。
本研究基于位点模型分析,发现鲤形目鱼类的ZP2基因中存在正选择的氨基酸位点(ω>1),表明ZP2基因在鲤形目鱼类演化中同样经历了正向选择,但ZP基因在鲤形目鱼类中是否经历扩张还需要对该类群进行更深层次的研究。
致谢:感谢中国科学院水生生物研究所张富铁老师、陈林、秦强在野外采集的样本,感谢张智和张晓宇在实验上提供的帮助。
