土壤水分对冬烟旺长期光合作用的影响
2021-05-25陈小龙尹光庭王松岭段卫东王建伟
陈小龙,卫 剑,尹光庭,王松岭,段卫东,王建伟,张 展*
(1. 河南中烟工业有限责任公司,河南 郑州 450000;2. 中国烟草总公司郑州烟草研究院,河南 郑州 450001)
【研究意义】环境条件对烤烟生长发育具有重要的作用,优质烤烟生产需要良好的生态环境以及适宜的栽培措施[1-2]。我国烤烟种植地域分布广阔,气候条件和生态环境丰富多样,不同生态烟区烤烟营养代谢和栽培措施具有较大差异[3-4]。云南德宏作为我国典型的冬烟种植区域,明确冬烟在不同土壤水分条件下的光合作用变化规律,对充分利用当地光照、水分资源条件,发挥冬烟优质栽培潜力具有重要的意义。【前人研究进展】按照烤烟在季节中的分布可以分为春烟、夏烟、秋烟和冬烟四种类型,我国大部分地区主要种植春烟,春烟的种植面积占我国烤烟总面积的90 %以上[5]。但是,我国部分地区因其独特的地理环境、气候条件和农事生产特点,烤烟的移栽时间会集中在冬至以前,采用这种移栽节令的烤烟称作冬烟[5-6]。冬烟能够充分利用冬闲耕地和劳动力资源,缓解粮烟“争地”矛盾,提高复种指数,在20世纪80年代发展较快。但由于冬烟受制于产量和品质的局限,近年来种的植面积不断减小[7],所以有关冬烟配套栽培技术和资源利用分析的研究报道较少。我国冬烟的种植区域主要分布在云南南部、广东南部、广西南部和福建东南部,这些烟区冬季气候温暖,热量条件丰富,年无霜期在300 d以上,烤烟的移栽期在11-12月,翌年4-5月收烤[5]。光合作用是烤烟生长发育十分重要的生理过程,烤烟的光合作用受外部环境影响强烈,特别是环境中的水分,直接影响烤烟的光合效率以及同化产物分配方向[8],研究烤烟的光合作用变化规律有利于采取合适的栽培措施提高烤烟品质。国内外许多学者对烤烟的光合作用进行了大量研究,烤烟光合作用变化规律以及作用机制已有较多阐述[9-12],但研究类型主要是针对春烟,有关冬烟的相关论述较少。【本研究切入点】由于冬烟生态区域的气候条件与我国大多数烟区差异较大,所以栽培措施也存在明显不同,关于冬烟同化作用鲜见报道,尤其是不同水分环境下冬烟的光合作用特征未见系统阐述,因此有必要针对冬烟的水分利用效率以及光合作用特征开展深入的研究。【拟解决的关键问题】本文以我国典型冬烟产区云南德宏为代表,探索不同土壤水分条件下冬烟在旺长期光合作用日变化特征,旨在为丰富烟草光合作用科学资料以及冬烟优质栽培技术提供依据和支撑。
1 材料与方法
1.1 试验地自然概况
试验地点设在云南德宏州盈江烤烟育苗基地,该地区种植的烤烟主要为冬烟。试验地点位于北纬24°32′、东经97°36′,海拔957.6 m,年平均气温18~20 ℃,年均极端最低气温2.2 ℃、极端最高气温33.8 ℃,全年>10 ℃的活动积温为7283 ℃,年平均日照时数2364 h,年平均降水量为1464 mm,主要集中在6-8月。
1.2 试验材料
烤烟供试品种为云烟87,该品种为云南德宏烤烟主栽品种,种子由云南省烟草农业科学研究院提供。采用避雨栽培盆栽试验,以称重法控制土壤含水量。试验用土壤为红壤土,田间最大持水量为23.6 %,土壤pH 6.7,碱解氮69.8 mg/kg,速效钾68.3 mg/kg,速效磷9.6 mg/kg,有机质16.2 g/kg。将土壤风干后装入一次性塑料盆,每盆装风干土20 kg,土壤中施入烟草专用肥,氮肥含量为每盆1.5 g,其中m(N)∶m(P2O5)∶m(K2O)比例为1∶1.5∶2。选取生长健壮的烟苗在2018年11月25日进行移栽,每盆移栽2株,共移栽60盆。
试验采用随机区组设计,土壤相对含水量设置4个梯度,即W1:25 %~30 %,W2:45 %~50 %,W3:65 %~70 %以及W4:75 %~80 %,每个梯度设置5盆,重复3次。烟苗移栽后正常生长45 d,在烟株旺长期采用称重法进行水分调控。对供试烟株处理1周后进行光合作用测定,不同处理土壤水分调控1周内土壤相对含水量详见表1。
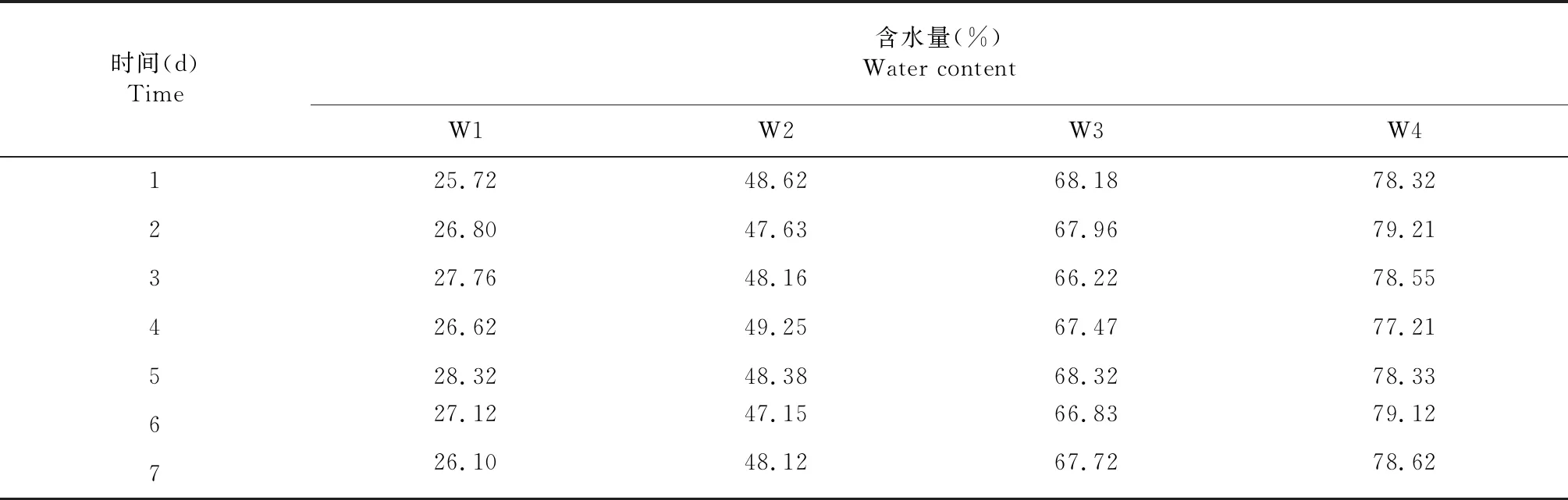
表1 各处理土壤相对含水量的变化
1.3 光合作用的测定
使用Li-6400便携式光合仪对各处理烟株进行田间活体测定。测定时各处理随机选取生长良好、长势均匀的烟株,自上而下选取烟株第6片烟叶进行测定,每个处理分别测定5片。测定时间为早上6:00至晚上18:00,每隔2 h测定1次,每个处理测定5个叶片,取其平均值。用于测定的光合指标主要包括:净光合速率(Pn,μmol CO2·m-2·s-1)、蒸腾速率(Tr,mmol H2O·m-2·s-1)、光合有效辐射(PAR,μmol·m-2·s-1)、气孔导度(Gs,mol H2O·m-2·s-1)、细胞间CO2浓度(Ci,μmol·mol-1)、田间CO2浓度(Ca,μmol·mol-1)、气温(Ta,℃)及空气相对湿度(RH,%)。气孔限制值(Ls,%)[13]根据公式Ls=1-Ci/Ca计算,叶片瞬时水分利用效率(WUE,μmol CO2mmol-1)[14]根据公式WUE=Pn/Tr计算。
1.4 气孔参数的测定
待光合作用测定完后立即采摘所测定的叶片,采用改良刮制法[15]对叶片进行处理,并将其制成永久切片。利用数码显微镜摄像系统在40倍物镜下观察叶片气孔的特征,测量气孔的长度、宽度以及气孔密度。单个气孔面积按照公式S=π×a×b进行计算[16],其中,a代表1/2倍气孔长度的,b代表1/2倍气孔宽度。
1.5 叶绿素含量的测定
采用 φ 为 80 % 的丙酮浸提比色法测定采摘叶片的叶绿素含量[17]。
1.6 数据分析
采用SPSS软件对试验数据进行方差分析和相关性分析,Excel软件进行绘图。
2 结果与分析
2.1 冬烟生长环境因子日变化规律
由图1可以看出,冬烟主要生长环境因子PAR和Ta的日变化曲线呈现出“单峰”型。早上6:00左右PAR开始升高并且上升速度较快,到中午12:00左右达到峰值,之后开始迅速下降。Ta随着PAR的变化而改变,峰值出现在下午14:00左右,比PAR的日间最大值延迟2 h,Ta与PAR呈极显著正相关(R=0.930**)。Ta的日差值较大,最高温度比最低温度高15.8 ℃。RH的变化趋势随着PAR和Ta的升高而降低,在早上10:00以前,RH下降迅速,中午12:00以后RH又开始缓慢上升,RH与PAR(R=-0.866**)和Ta(R=-0.916**)呈显著负相关。Ca的日变化也呈现出明显的规律,早上6:00左右最高,之后开始逐步减低,下午16:00达到最低值,之后又略有回升。
2.2 冬烟在不同水分条件下光合参数的日变化特征
2.2.1 净光合速率日变化 由图2可知,处理W2、W3和W4的Pn日变化均呈现“双峰”型,2个峰值分别出现在早上10:00和下午14:00左右,上午的峰值大于下午,并且2个峰值之间存在显著差异(P<0.05),中午12:00左右出现明显的光合“午休”现象。W1的Pn日变化进程与其它处理存在明显不同,日变化呈现“单峰”型,最高值出现在上午10:00左右。从Pn日变化进程的斜率分析,随着土壤含水量的降低,烟株的Pn升高和降低速率逐渐变缓。不同水分条件下烟株的Pn日均值存在明显差异,大小依次为W4>W3>W2>W1,烟株的Pn日均值随土壤水分的降低而降低,其中W4显著大于W1(P<0.05),但与W2和W3差异不显著(P>0.05)。土壤水分对烟株的Pn具有较大的影响。
2.2.2 蒸腾速率日变化 不同土壤水分条件下烟株的Tr日变化进程存在明显差异(图2)。其中处理W2、W3和W4的Tr日变化进程呈现明显的“双峰”型,而W1表现为“单峰”型。处理W2、W3和W4的Tr2个峰值分别出现在早上10:00和下午14:00,并且第二个峰值均大于第一个峰值。处理W1的峰值出现在早上8:00左右,之后逐渐下降。不同土壤水分的烟株Tr日均值表现不同,土壤含水量越低,烟株的Tr越低,W4显著大于W1和W2(P<0.05),与W3差异不显著(P>0.05)。土壤水分对烟株的Tr在不同时间段的变化速率也有一定影响,随着土壤含水量降低,烟株Tr的日变化趋势逐渐变缓。
2.2.3 气孔导度日变化 由图3可以看出,土壤水分对烟株的Gs影响较为明显。处理W2、W3和W4的Gs日变化均呈“双峰”型,但峰值出现的时间不同。W4和W3的2个峰值分别出现在早上8:00和下午14:00左右,W2的第2个峰值出现在下午16:00左右。处理W1日变化呈现“单峰”型,峰值出现在早上8:00左右。不同处理Gs的日均值存在明显差异,大小为W4 >W3 >W2 >W1。在早上8:00以前,W4和W3的Gs升高速率明显高于W1和W2,8:00-12:00随着Ta的升高以及HR的降低,各处理烟株Gs呈现下降趋势,W4降低幅度最大。晚上18:00左右,处理W1的Gs略高于其它处理。
2.2.4 胞间CO2浓度日变化 由图3可知,不同土壤水分条件下烟株的Gi在早上6:00左右达到一天中的最大值,并且数值较为接近。上午12:00以前,各处理烟株的Gi一直保持下降趋势,12:00-16:00之间处理W2、W3和W4的Gi变化趋势为先升高、再降低,而W1烟株的变化趋势截然相反,表现为先降低、再升高。各处理烟株的Gi日均值大小依次为W4 >W3 >W2 >W1。
2.2.5 气孔限制值日变化 处理W2、W3和W4的Ls呈“双峰”型,2个峰值分别出现在12:00和16:00左右;W1烟株的Ls呈“单峰”型,峰值出现在下午14:00左右。各处理Ls的日均值表现为W1 >W2 >W3> W4。
2.3 水分利用率的日变化特征
不同处理烟株的WUE日进程存在差异(图4)。处理W1和W2的WUE日变化呈现“双峰”型,两个峰值分别出现在8:00和18:00,而W4和W3在8:00左右达到峰值,之后逐步降低。不同处理烟株的WUE日均值大小依次为W2>W1>W4>W3,处理W2的WUE显著高于W4和W3(P<0.05),并且日均值是W4的1.6倍,W2与W1烟株的WUE差异不显著(P>0.05),随着土壤含水量的降低,烟株提升了土壤水分的利用效率。各处理烟株的WUE表现为上午高于下午,处理W1~W4在下午的WUE分别较上午下降82 %、69 %、46 %和55 %。
2.4 冬烟光合速率和蒸腾速率与生理生态因子的相关性分析
相关性分析表明(表2),不同土壤水分条件下烟株的Pn与PAR、Ta和Gs正相关,与HR和Gi负相关;处理W1的Pn与Gs极显著正相关,其它处理烟株Pn与Gs显著正相关;W4的Pn与PAR极显著正相关,与Ta显著正相关,其它处理烟株Pn与PAR、Ta正相关,但相关性不显著。处理W4的Tr与PAR极显著正相关,与Ta和Gs显著正相关;W2和W3的Tr与PAR、Ta和Gs显著正相关;W1的Tr与Gs极显著正相关,与PAR和Ta正相关,但相关性不显著;不同处理烟株的Tr与Ga、HR和Gi负相关,相关性不显著。这说明在水分充足的条件下影响烟株Pn和Tr的主要因子是PAR,其次是Gs和Ta,但烟株在缺水的环境下影响Pn和Tr的主要因子是Gs,并且随着土壤含水量的降低,这种表现越发明显。
2.5 水分对冬烟气孔特征及光合色素的影响
水分对冬烟气孔特征的影响较大,不同土壤水分条件下叶片气孔的大小存在明显的不同(表3)。减少水分会引起烟株气孔长度和宽度出现不同程度的降低,W1、W2、W3的气孔长度和宽度较W4分别降低12.7 %、13.5 %、10.5 %、10.0 %、2.3 %、3.4 %,并且W4与W1、W2存在显著差异(P<0.05),与W3差异不显著(P>0.05)。气孔长度和宽度的降低造成叶片单个气孔面积以及每平方毫米气孔总面积减少,但气孔密度却随着水分的减少而增加,W1和W2的气孔密度显著高于W4(P<0.05),W3略高于W4,但差异不显著。

表2 不同水分处理叶片光合速率和蒸腾速率与生理生态因子的相关性分析

表3 不同水分处理叶片气孔特征参数分析
由图5可知,土壤水分影响叶片光合色素含量,但不同光合色素对水分的响应程度存在差异。土壤水分减少,不同处理叶片Chl a、Chl b和Car的含量均降低,大小表现为W1
3 讨 论
烤烟对外界环境的变化十分敏感,研究光合作用日变化有利于明确烤烟干物质的积累与代谢能力,同时为分析环境因素对烤烟生长发育的作用提供重要手段[18-19]。本文发现,土壤水分变化会造成冬烟的Pn日变化趋势出现较大改变,不同水分条件下冬烟的Pn日变化进程出现“单峰”和“双峰”两种类型。邓仲篪等[20]表明,植物在高温以及强光照条件下会出现“午睡”现象。本研究中,处理W2、W3和W4的冬烟Pn日变化曲线表现为“双峰”型,具有明显的光合“午休”现象。这主要是因为午间光合有效辐射和温度较高,空气相对湿度维持在较低水平,植物为降低蒸腾作用,关闭大部分气孔,从而导致冬烟Pn降低。当土壤水分小于30 %时,冬烟Pn日变化呈现“单峰”型,这可能与叶片光合酶活性降低有关[21]。当处理W1的Pn达到最大值,Tr也最大,严重缺水环境下叶片水分进一步亏缺,水分亏缺导致叶肉细胞CO2扩散阻力增大,W1的胞间CO2浓度显著小于W4(P<0.05),光合酶类物质活性下降,叶片获取的激发能超过自身碳同化能力,光能发生过剩耗散,叶片对光能的利用效率减弱[22],所以造成Pn下降。12:00-18:00之间W1的Pn一直处于下降趋势,主要是因为气温较高,同时空气相对湿度一直处于较低的水平,叶片水分亏缺造成大部分气孔处于关闭状态。赵铭钦等[23]和张广富等[24]研究表明延吉地区种植的夏烟Pn日最大值分别为10.58和10.21 μmol CO2·m-2·s-1,日均值分别为7.83和7.74 μmol CO2·m-2·s-1,本文中正常水分条件下处理W4的Pn日最大值为15.21 μmol CO2·m-2·s-1,日均值为8.62 μmol CO2·m-2·s-1,冬烟的Pn高于先前报道的夏烟,这可能与冬烟种植的地理环境有关。江厚龙等[25]研究发现,烤烟的Pn与海拔密切相关,烤烟光能转换效率和光合潜能均随海拔的升高而增加。本文的试验地点云南德宏属于低纬度高海拔地区,试验地的海拔高度接近1000 m,这可能是造成冬烟净光合速率较高的重要原因。
李正风等[26]研究发现土壤相对含水量在75 %~80 %之间,不同基因型的夏烟蒸腾速率平均值在3.5 mmol H2O·m-2·s-1左右,相同水分条件下冬烟蒸腾速率平均值为2.3 mmol H2O· m-2·s-1。已有研究表明,一般情况下夏烟的蒸腾速率日均值在3.79~4.30 mmol H2O· m-2·s-1之间[22],本研究中冬烟在不同土壤水分下的蒸腾速率均低于前人所报道夏烟,这可能与当地生态条件下冬烟气孔导度始终维持在较低的水平有关。植物蒸腾作用的主要途径是气孔,空气温度和环境相对湿度影响叶片的气孔的开放程度[27]。本研究的试验地在10:00-16:00的大气平均温度超过25 ℃,空气相对湿度在35 %左右,高温低湿的外界环境造成冬烟气孔导度保持在较低的水平。同时,植物的蒸腾作用不仅与自身基因型和生理生化有关,同时还受到外界条件的影响,尤其是与PAR和Ta等环境因子密切相关[28-29]。邓云等[30]在研究巨尾桉蒸腾耗水特性时指出,Ta可以影响叶片Gs,进而对Tr造成影响。曹生奎等[31]发现胡杨的Tr与PAR、Ta呈多项式关系。本研究表明,不同冬烟的蒸腾速率与PAR、Ta及Gs呈极显著或显著正相关。但土壤水分含量不同,冬烟Tr受环境因子的影响存在差异。处理W4的烟株在水分充足的环境下,生态因子对冬烟Tr起主要作用,主要生态因子是PAR,次要生态因子是Ta;处理W1的烟株在缺水时,影响Tr的主要因子是生理因子Gs。这种现象可能是冬烟在受到逆境胁迫时,为了满足生长发育的需求而表现出的适应性和抵抗能力。
气孔作为叶片与外界环境进行气体与水分交换的门户,对植物的光合作用和蒸腾作用起着重要的调节作用[32]。气孔特征主要由遗传因素决定[33],但外界环境条件也对气孔形态具有重要的影响,尤其是土壤水分。前人研究表明,在干旱环境下小麦、白沙蒿和臭柏的叶片气孔密度会随着干旱程度的加剧而增加,同时气孔的分布也与对照有明显的差异[34-36];徐坤等[36]认为减少水分会使生姜叶片的气孔密度以及气孔面积减小;陈征等[38]发现烤烟品种中烟100在干旱的情况下气孔密度会显著增加。本研究中冬烟的气孔密度随土壤水分的减少而增加,与之前的研究结论一致。这主要与烟株在缺水的环境下烟叶开片受到影响,单位面积缩小导致气孔数目增加有关。此外,Makbul 等[39]认为植物在干旱胁迫下叶片气孔会减小。本研究W1和W2的气孔长度、宽度以及气孔面积显著低于W4,水分减少造成叶片气孔大小显著降低,这可能是由于土壤的干旱程度超过了烟株的承受能力,烟株体内的膨压和组织水势下降明显,导致气孔发育受到影响[40]。
叶片光合色素影响植物的光合作用,干旱胁迫会导致植物光合色素降低,进而造成植物的光合能力的下降[41]。本研究中,叶片光合色素含量随水分的减少而越低,但光合色素种类不同,下降程度也存在差异。Chl b的下降比例最大,其次是Chl a,Car的下降比例最小,这可能与光合色素在植物中作用不同有关。已有研究表明,叶绿素的主要作用是吸收、传递光能,并通过光敏化特性进行光化学反应,而类胡萝卜素除了具有吸收光能外的作用外,还可以在强光下逸散能量,淬灭活性氧,防止膜脂过氧化[42]。土壤水分含量减少造成叶绿素含量降低、气孔导度下降、胞间CO2减小,继而导致叶片碳同化能力降低,光照出现过饱和。为了减少强光对叶绿体的伤害,烟株通过维持相对较高的类胡萝卜素含量以保持淬灭活性氧的能力。叶片光合色素含量的变化与植物利用资源的生长策略密切相关[43],烟株控制自身色素之间的降解差异以抵御环境中不利因素,通过自我调节以适应生存环境。
4 结 论
土壤水分对旺长期冬烟的光合作用、气孔形态以及光合色素含量有较大影响。减少土壤水分会明显降低烟株的光合能力,但土壤水分降低程度不同,烟株的净光合速率、蒸腾速率、气孔导度、水分利用效率的日变化进程有较大差异。叶片的气孔长度、气孔宽度以及气孔面积随水分的减少而降低,但气孔密度随水分的减少而增高。减少水分会降低叶片光合色素含量,但不同类型光合色素降低比例不同,叶绿素b下降幅度最大,其次是叶绿素a,类胡萝卜素下降幅度最小。综上所述,旺长期水分供应不足会造成冬烟叶片功能受损,光合能力下降,影响冬烟的产量和品质,土壤相对含水量保持在65 %以上可以有效避免旺长期内水分亏缺对冬烟生长发育造成影响。
