异迟眼蕈蚊生物生态学特征及防治策略
2021-05-12苟玉萍刘长仲张克信李春春张强艳张艳蕾
苟玉萍,毛 亮,刘长仲,张克信,李春春,张强艳,张艳蕾
(1. 甘肃农业大学植物保护学院 / 甘肃省农作物病虫害生物防治工程实验室,甘肃 兰州 730070;2. 临洮县林业和草原局,甘肃 临洮 730500)
异迟眼蕈蚊(Bradysia impatiens Johannsen)是常见于蘑菇房、设施大棚、花盆、苗圃等场所的重要害虫,俗称菌蛆,隶属于双翅目(Diptera) 长角亚目(Nematocera)眼蕈蚊科(Sciaridae) 迟眼蕈蚊属(Bradysia)[1-4]。该虫食性极广,主要以幼虫为害韭菜( Allium tuberosum )、葱(A. fistulosum)、百合(Lilium brownii)、胡萝卜(Daucus carota)、一品红(Euphorbia pulcherrima)、蝴 蝶 兰(Phalaenopsis aphrodite)、草莓(Fragaria × ananassa)、桉树(Eucalyptus robusta)等植物的根、茎、叶、花,甚至整棵幼苗[5-7],也可以真菌和腐殖质为食,具有半腐生性[8]。此外,实验室内常观察到异迟眼蕈蚊可对紫花苜蓿(Medicago sativa)造成严重为害,但相关研究尚未报道。
关于异迟眼蕈蚊的学名多有争议,已报道的有Bradysia (Chaetosciara) tristicula var. difformis Frey、Sciara (Lycoriella) hardyi Shaw 、 Bradysia paupera Tuomikoski 以 及Bradysia agrestis Sasakawa 等,而Bradysia difformis Frey 是被广泛认可的[9],我国学者也多采用此名[1,10-13]。然而,近年来Mohrig 等[14]将Bradysia difformis 归为Bradysia impatiens 的异名,由Johannsen 于1912 年首次报道于北美,幼虫模式标本采自于凤仙花(Impatiens balsamina)根部附着的土壤中。Ye 等[15]通过DNA 编码和系统发育分析,证实了Bradysia impatiens 为该种的学名。Sueyoshi和Yoshimatsu[16]通过对日本地区采集到的1 738 头迟眼蕈蚊属成虫的标本进行形态鉴定,进一步明确Bradysia difformis 为异迟眼蕈蚊Bradysia impatiens的异名。
该虫分布范围极广,属世界性害虫,美国、南非、日本、荷兰、巴西、夏威夷群岛、英国、俄罗斯等国家均有报道,已对农林经济造成巨大损失[2,8-9]。2009 年,因该虫对中国云南省茶树菇(Agrocybe aegerita)、平菇(Pleurotus ostreatus)等食用菌造成危害而被首次记录,并已成为该地区食用菌害虫的优势种[10],以幼虫取食菌丝体造成出菇量下降。异迟眼蕈蚊与韭菜迟眼蕈蚊(Bradysia odoriphaga Yang et Zhang)形态相似,主要区别在于雄性鞭毛以及平衡棒的生殖刺突和着色[16],两者常混合发生,是甘肃天水和辽宁沈阳等地区韭菜害虫的优势种[17-18]。本文对异迟眼蕈蚊生物生态学特性及防治方法的相关研究进展进行总结和阐述,以期为该虫的进一步研究提供基础的理论依据。
1 生活习性及为害
1.1 成虫及产卵
成虫不取食,羽化也无明显的昼夜节律,多发生在傍晚至次日上午,雄虫羽化高峰集中在17:00 -23:00,而雌虫则在16:00 - 21:00,雌 ∶ 雄 > 1 ∶ 1,交尾时呈“一”字形,时长约为3 min,且雄虫可多次交尾[13]。其寿命长短与环境条件密切相关,一般随温度升高逐渐缩短(20~28 ℃),雌虫历期比雄虫长,25 ℃时雌虫历期约为2.65 d,雄虫历期约为1.85 d[17]。
卵趋向隐蔽黑暗场所,多产于土壤缝隙或植物基部与土壤间的空隙处,单雌产卵量随寄主和环境条件的不同而异,如果环境不适宜则产卵量减少或不产卵[17]。适宜温度范围内产卵量随温度的升高而增加,25 ℃时最高,单雌产卵量可达111.06 粒,显著高于10、15、20 和30 ℃[5]。以韭菜为寄主植物的繁殖力显著高于葱、蒜(Allium sativum)、百合、蚕豆(Vicia faba)、 生 菜(Lactuca sativa)、 白 菜(Brassica pekinensis)、甘蓝(Brassica oleracea)等植物[6]。此外,光和雌雄性比等因素也可对异迟眼蕈蚊的产卵量造成不同程度的影响。Liu 等[5]研究表明,光周期为L ∶ D = 16 h ∶ 8 h 和光强度为588 lx 条件被证实更有利于异迟眼蕈蚊的繁殖。Gou 等[1]研究表明,异迟眼蕈蚊雌雄成虫性比为1 ∶ 1 时繁殖力显著高于1 ∶ 2、1 ∶ 3 和3 ∶ 1 时。由于雄虫一生可多次交配,所以即使局部性别比例失调也可繁衍,这也是造成异迟眼蕈蚊局部发生严重的原因之一[19]。此外,经室内饲养观察,雌虫产卵时移动性小,随产卵的进行腹部逐渐收缩,产完卵便在卵堆处死亡,或死亡时腹部仍留有卵。
1.2 幼虫为害
异迟眼蕈蚊是杂食性昆虫,幼虫多钻蛀到食用菌、药用菌和野生菌的菌料中,一般先从基部为害,取食菌料中的菌丝体和有机质[10],也常在菌褶内为害,严重时,将菌柄吃成海绵状,菌盖只剩上面一层表皮,进而枯萎腐烂。当异迟眼蕈蚊蛀食植物根部,主要对植物表皮和皮层组织造成伤害,导致植物根系受损而影响水分和养分的吸收,植物生长后期地上部分则表现出萎蔫、活力丧失、营养减少和落叶等症状,并且根部磨损处会出现棕褐色的小虫疤[20-21]。此外,植物根系受损还会限制碳水化合物的储存量,削弱幼苗的结构支撑。因此,幼苗易倒伏,并进一步成为异迟眼蕈蚊的食料来源[21]。Shin 等[22]根据迟眼蕈蚊属的食性,将其幼虫栖息地分为3 种不同类型:活植物(啃食植物根茎叶)、腐木(存活于枯木或树皮下)和植物凋落物(存活于腐殖质、落叶层或土壤中)。幼虫以咀嚼大多数植物器官为食,尤其是幼嫩发育的根尖系统、嫩根和根毛,对根系造成严重损害[9]。幼虫还可以损害植物木质化的部分[23]。拟南芥(Arabidopsis thaliana)幼苗明显比成熟植株更易受到迟眼蕈蚊属的为害,从而导致受影响幼苗的生长和存活率降低[20]。幼虫不仅直接取食和钻入植物根茎对植物造成物理伤害[21-24],而且可以间接性地为植物病原体的进入、传播、转移提供途径,例如腐霉病、葡萄球菌病、柱状芽孢杆菌病、菌核病和根腐病等[25-26](表1)。

表 1 异迟眼蕈蚊的为害方式及症状Table 1 Damage patterns of Bradysia impatiens
2 发生规律
在温室和塑料大棚内,异迟眼蕈蚊可终年发生危害,有世代重叠现象,3 月下旬、5 月中下旬及9 月下旬是成虫种群数量高峰期的3 个阶段,幼虫的发生盛期则在5 月上中旬和10 月上旬以后[11-12,27]。在温室外或非保护地,该虫一年发生4~6 代,以幼虫在4 cm 左右处的韭菜假茎基部及周围土壤越冬,成虫从3 月下旬 - 4 月中旬数量陆续上升,5 月中下旬、6 月下旬 - 7 月上旬以及9 月下旬为发生高峰期;幼虫的高峰期则在5 月下旬 - 6 月中旬和10 月上旬以后[11]。全世代发育历期一般为23~27 d,其中卵期为6.5 d 左右,幼虫期为13.5 d 左右,蛹羽化为成虫需要5.5 d 左右,成虫期平均为3~7 d[17]。研究表明,异迟眼蕈蚊在葱田以3 龄幼虫在15 cm深的葱根上越冬,并在韭菜、葱、蒜等寄主间迁移为害,幼虫孵化后便向下转移,若近地面有烂叶、伤口,或湿度较大、寄主本身含水量较高,极易成为幼虫的侵入点,老熟幼虫多远离寄主,寻找隐蔽处化蛹[18]。在生长季节田间各虫态均有,但应以占优势的虫态为准来确定防治适期和防治策略。因此,幼虫应在5 月下旬、6 月中旬和10 月上旬各防治一次,而成虫应在5 月中下旬、6 月下旬 - 7 月上旬以及9 月下旬各防治一次。
3 发生与环境条件的关系
3.1 寄主植物
寄主植物可对植食性昆虫的种群数量变化产生影响[28]。取食不同寄主植物不仅影响昆虫的生长发育、繁殖及体内物质[29-31],还会影响其对温度的耐受性以及对药剂的敏感性[32-35],并且两者在某种程度上是协同进化的。昆虫选择寄主植物与其自身的遗传、生活习性以及外界环境有关[36],同时,与寄主植物的挥发性物质息息相关[37]。昆虫通过嗅觉器官、或视觉器官、或两者的协同作用来定位寄主、检测植物[38]。异迟眼蕈蚊可为害百合科蔬菜,尤其对韭菜的嗜食性和适应性较好,种群扩增速度显著优于大葱、蒜茎、百合和蒜瓣[39]。通过构建异迟眼蕈蚊在20 个韭菜品种上的实验室种群生命表,发现该虫对‘平韭二号’的选择性和适应性较好,内禀增长率(rm) 为0.22 d-1,‘冀研小韭园’和‘791 韭菜王’ 次之(rm= 0.20 d-1和rm= 0.19 d-1),而‘万源紫根宽叶王’和‘万源汉中冬韭一号’相对较弱(rm= 0.09 d-1和rm=0.07 d-1)[40]。此外,通过对百合科(韭菜)、豆科(蚕豆)、十字花科(白菜和甘蓝)、菊科(生菜)植物的寄主选择性相关研究,发现该虫对韭菜和蚕豆具有选择偏好性,取食后平均世代周期较短(20.78 和19.56 d)、繁殖力较强(产卵量为91.60 和78.07 粒);生菜次之(世代周期为21.32 d和产卵量为64.13 粒);而对白菜和甘蓝选择性较弱,取食后平均世代周期较长(23.86 和25.17 d)、成虫繁殖力减弱(产卵量为63.73和60.67 粒)[6]。上述结果可用于防治,即选择对该虫嗜食性差的韭菜品种种植或者避免韭菜和蚕豆轮作来抑制种群扩增速率,减轻为害。
3.2 温度
昆虫是一种典型的变温动物,易受环境因子胁迫,尤其是温度,其繁衍和发生与温度密切相关[41]。当温度超过或低于昆虫的适宜温度,短时间的温度胁迫不会使其迅速死亡[42]。即使能够存活,其后续发育仍会受阻,对其发育速率、繁殖和生理生化等产生抑制效应[43-44]。各虫态发育历期随温度的变化而不同,在适温范围内随温度的升高而缩短。成虫在10~30 ℃时平均寿命由10.08 d 降至1.28 d;卵期由12.15 d 降至3.75 d;幼虫期由44.64 d 降至9.13 d;蛹期由14.80 d 降至2.92 d[17]。温度也影响产卵量,适宜范围内随温度的升高而增加,25 ℃时最高,达111.06 粒。有研究表明,当温度高于36 ℃时各虫态迅速死亡,且存活幼虫个体发育期延长,成虫寿命缩短,产卵量下降,但蛹的耐热性强于幼虫和成虫[12]。综上所述,温度过高或过低均不利于各虫态发育和存活,温度在25 ℃左右时,有利于该虫种群繁殖和扩增,而高温影响产卵及各虫态存活,这就是该虫春、秋季发生严重的缘由之一。
3.3 光
光并非昆虫生存的必要条件,但却对其发育历期、存活率、滞育及繁殖等生物学特性、生活习性和行为有显著影响[45-48]。昆虫通过视觉器(单眼和复眼)的共同作用感受光信号,且表现出一定的趋光性,这与光的性质即颜色或波长有关[47]。光的照度,包括光照强度和光周期,则对昆虫的起飞迁出、取食及其体色等产生影响[5]。Liu 等[5]研究了异迟眼蕈蚊成虫在5 种光照强度和光周期下的历期,结果表明,在光周期L ∶ D = 16 h ∶ 8 h 和光强度588 lx 条件下,成虫寿命平均为2.30 d。光对成虫产卵也有显著影响,光周期L ∶ D = 16 h ∶ 8 h 时产卵量为101.54粒,而L ∶ D = 0 h ∶ 24 h 时为73.33 粒,L ∶ D = 24 h ∶ 0 h时为69.5 粒;光强度为588 lx 时,产卵量为101.54 粒,1 275 lx 时 降 到94.65 粒,1 574 lx 时 降 到79.36 粒,2 072 lx 时降到70.78 粒,3 323 lx 时降到69.78 粒[5]。异迟眼蕈蚊幼虫喜欢于阴暗隐蔽处活动,但成虫喜弱光,产卵量随光强度的升高而降低。由此可见,可利用长时间强光照射来防治该虫。异迟眼蕈蚊对黄色或绿色的发光二极管(light-emitting diode, LED)以及紫外(ultraviolet, UV)照射较敏感,已被应用于该虫的防治[48]。
4 防治策略
4.1 农业防治
异迟眼蕈蚊已成为甘肃天水、武山、甘谷等地韭菜温室大棚中的优势种,严重威胁该地农业发展,而选择抗性较强的韭菜品种,如‘万源紫根宽叶王’和‘万源汉中冬韭一号’等,是农业防治最安全有效的手段[40]。此外,培育韭菜苗时选择无害虫的土壤,移栽韭菜时清除带虫的韭菜苗,选择卫生条件良好的栽培田地,也可有效避开虫源,降低虫口密度[49]。增效剂与杀虫剂联用防治异迟眼蕈蚊可起到增效减量的作用,更有益于绿色农业的发展[50]。在食用菌和药用菌培育中加强管理,定期对培养料进行消毒灭菌、使用杀菌剂对菇房进行熏蒸灭菌等,均可有效抑制虫害的发生[51]。
4.2 物理防治
高温对昆虫的影响极其显著,容易引发昆虫系统性细胞死亡,最后导致虫体死亡[52]。据各地实践,在6 月 - 9 月的高温关键季节,采用日晒高温闷棚或高温覆地膜法,提高大棚内部或土壤的温度,不仅可以熟化土壤,改善土壤质量,而且对各种土传病和地下害虫的虫卵和蛹有较好的灭杀效果。据报道,在高温闷棚或覆地膜前进行灌溉,可有效降低植株在闷棚期间遭受的损害[53]。史彩华[54]利用“日晒高温覆膜法”防控韭蛆,结果显示,该法对韭菜迟眼蕈蚊各龄期幼虫的致死率高达100%,而对韭菜的长势和根系发育无明显影响。目前,高温覆膜法主要应用于韭菜迟眼蕈蚊的防治,对异迟眼蕈蚊的报道较少,但此法成本低、见效快且绿色环保,值得借鉴和推广。
4.3 生物防治
4.3.1 昆虫病原线虫
昆虫病原线虫(entomopathogenic nematodes)是一种以昆虫为专性寄主的致病性微生物,它们常从昆虫的自然孔口,如口器、气门或肛门等进入虫体,或直接穿过节间膜侵入虫体,不仅对虫体造成机械损伤,也会随即释放所携带的共生细菌,使虫体得败血症而亡[55-57]。因其主动搜寻寄主、安全高效而成为世界性的生物杀虫剂[58]。该微生物主要通过识别昆虫排泄物中的尿酸和尿囊素等物质、粪便散发出的异味以及呼吸释放的CO2等方式来主动寻找寄主昆虫[58]。钱秀娟等[55]发现夜蛾斯氏线虫(Steinernema feltiae) 0619HT品系与吡虫啉联用后,可提高对异迟眼蕈蚊的防治效果(表2)。Ernesto等[59]研究表明,亚马逊小杆线虫(Heterorhabditis amazonensis) LPV498 品系对异迟眼蕈蚊的防效较好,并且温室优于大田(表2)。Acharya 等[60]采用印度小杆线虫(Heterorhabditis indica)和小卷蛾斯氏线虫(Steinernema carpocapsae)来防治异迟眼蕈蚊,结果显示,印度小杆线虫的致死率显著高于小卷蛾斯氏线虫,并且对4 龄幼虫的效果显著高于其他龄期(表2)。
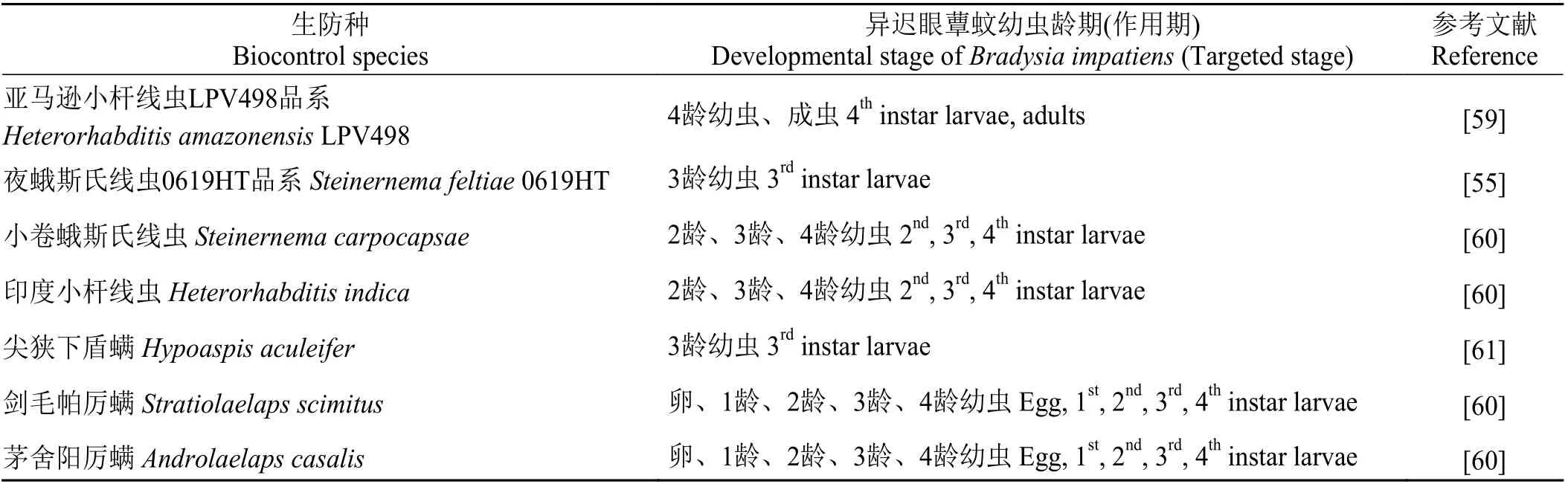
表 2 对异迟眼蕈蚊高效的昆虫病原线虫和捕食螨种类Table 2 Effective species of entomopathogenic nematodes and predatory mites against Bradysia impatiens
4.3.2 捕食螨
异迟眼蕈蚊的卵和幼虫均可成为捕食螨的捕食对象,其捕食的有效性与其生命周期及在土壤中的活动能力相关[62]。已有研究报道,将尖狭下盾螨(Hypoaspis aculeifer)作用于异迟眼蕈蚊幼虫,效果良好且持续时间久,尤其是每间隔7 d 接1 次螨,连续接3 次,防效可高达100% (表2)[61]。此外,Acharya等[60]追踪了剑毛帕厉螨(Stratiolaelaps scimitus)和茅舍阳厉螨(Androlaelaps casalis)对异迟眼蕈蚊的捕食情况,结果显示,剑毛帕厉螨的捕食率优于茅舍阳厉螨,并且对1 龄幼虫的捕食效果显著高于其他虫态(表2)。将昆虫病原线虫(小卷蛾斯氏线虫和印度小杆线虫)与捕食螨(剑毛帕厉螨和茅舍阳厉螨)联用,分别测定对异迟眼蕈蚊卵、1 龄、2 龄、3 龄和4 龄幼虫的致死率,结果表明,印度小杆线虫与剑毛帕厉螨联合显著提高了异迟眼蕈蚊的致死率[60],可作为一种改良的生物防治策略。
4.3.3 苏云金芽孢杆菌
苏云金芽孢杆菌(Bt)的芽孢会释放出具有杀虫活性的晶体蛋白,昆虫取食后中毒死亡[63-64],且对非靶生物安全和人畜无害,因此,Bt 已成为一种应用广泛的微生物杀虫剂。王帆帆等[65]研究表明,Bt 菌株70-4 为毒蛋白基因cry11 + cyt1 的复合型,对异迟眼蕈蚊的杀虫活性显著高于Bt 菌株69-5 和70-2 的复合型毒蛋白基因cry/10 + cry11 + cyt1 + cyt2。
4.4 化学防治
4.4.1 化学杀虫剂
化学杀虫剂因其成本低、效率高而被广泛应用于农业生产中。目前在韭菜、食用菌和药用菌生产中,防治韭菜迟眼蕈蚊和异迟眼蕈蚊等根蛆类害虫常见的化学药剂包括:1)有机磷类,毒死蜱和辛硫磷等;2) 新烟碱类,噻虫嗪和噻虫胺等;3) 复配剂,20%灭蝇胺·噻虫胺悬浮剂和48%毒·辛乳油等。曲绍轩等[66]测定了苦皮藤素乳油和印楝素乳油对异迟眼蕈蚊的毒力,结果显示,两种杀虫剂的校正死亡率均达80%以上,均具有良好的灭杀效果。沈登荣等[67]利用4 种苯甲酰基脲类杀虫剂测定了对异迟眼蕈蚊的致死率,结果表明,氟啶脲和氟虫脲对异迟眼蕈蚊的温室和田间防治效果较好。有研究报道,增效剂与杀虫剂联用可提高对异迟眼蕈蚊的防效,溴氰菊酯和高效氯氟氰聚酯的增效作用强于吡虫啉和噻虫啉[50]。
4.4.2 植物源杀虫剂
植物源杀虫剂因其安全、低毒、低残留、易降解且抗药性低而被广泛应用于害虫防治[68-69]。程东美等[7]采用浸虫法和混合法对除虫菊素、印楝素、鱼藤酮和苦参碱4 种植物源杀虫剂防治异迟眼蕈蚊的作用效果进行了评价,发现除虫菊素的生物活性较好,尤其对2 龄幼虫的效果优于3 龄和4 龄幼虫。
5 研究展望
目前,异迟眼蕈蚊猖獗为害是韭菜和食用菌(平菇、香菇、茶树菇等)生产中面临的主要瓶颈。异迟眼蕈蚊作为一种世界性农林害虫,针对其生物学特性、行为生态学及防治技术的研究逐渐得到关注,但是深度和广度还不够,未来可以从以下几方面开展研究:1)该虫各虫态对湿度的适应性差异尚未见研究报道。一般来讲,各虫态均选择相对湿润隐蔽的环境,但高湿或过度干燥会影响其存活和发育。研究中常观察到幼虫在高湿环境中身体会拉长,增加死亡率;卵过于干燥会出现干瘪,影响孵化,但具体湿度需进一步研究。2) 高温抑制该虫的生长发育,但机理尚不清楚,需要从生理生化、基因组学、代谢组学和蛋白组学等方面深入探究。3)在防治策略方面,一直以幼虫为重点研究对象,缺少对成虫的防治研究。熏蒸剂是灭杀成虫的有效方法之一,选择合适的植物源熏蒸剂,可达到安全有效、低残留的效果。此外,该虫主要发生于温室、大棚和菇房中,为熏蒸剂的使用提供了可能,在这方面有待取得突破。
