酸性核质DNA结合蛋白1的研究进展
2021-05-12崔亚婷勾文峰魏会强侯文彬李祎亮
崔亚婷,勾文峰,魏会强,于 江,侯文彬*,李祎亮*
(1.天津中医药大学,天津 301617; 2.中国医学科学院 北京协和医学院 放射医学研究所天津市放射医学与分子核医学重点实验室,天津 300192)
酸性核质DNA结合蛋白1(acidic nucleoplasmic DNA-binding protein 1, And-1)主要在细胞核中表达,具备WD40重复序列(WD-repeat,WD40,WDR)和高迁移率组蛋白框结构域(high mobility group-box motif,HMG-box motif)家族的双重特性[1],其结构复杂,功能多样。BLAST数据库显示,果蝇、秀丽隐杆线虫、拟南芥、神经孢子囊、非洲爪蟾中均检测到了与人类And-1高度相似的蛋白质, 表明该蛋白家族高度保守。And-1家族基因包括来自裂殖酵母的mcl1、酿酒酵母(Saccharomycescerevisiae)的Ctf4、曲霉(Aspergillusnidulans)的SepB以及人类的And-1。该家族基因包含SepB结构域,SepB盒是该蛋白家族独有的共性特点,也是区分WD40蛋白或其他传导蛋白重要的特征。主要影响DNA复制、修复以及姐妹染色体凝聚等[2]。And-1的结构与Ctf4和 Mcl1略有不同,除N端WD40和SepB结构域外,其在C末端还有一个HMG结构域。WD40能介导蛋白与蛋白的相互作用[3],而HMG蛋白则具有DNA结合的特性[4]。因此,它的功能比Ctf4/Mcl1更复杂、多样。
1 And-1的结构特征
And-1由氨基末端WD40重复序列、中间SepB结构域和羧基末端HMG框结构域共同组成(图1A),该蛋白又名WD重复序列与HMG-box DNA结合蛋白1(WD-repeat/-domain and HMG-box/-domain DNA binding protein 1, WDHD1)。其包含1 127个氨基酸,总分子质量为125 ku,pI为5.27[5]。WD40结构域包含7个叶片结构,形成1个环状的α-螺旋桨(图1B)。SepB结构域包含一个WD40子结构域和一个螺旋子结构域,其中WD40子结构域包含6个串联刀片状结构,形成了具有间隙的环形结构(图1C);螺旋子结构域由5个α螺旋(α1-α5)组成,以螺旋束的形式位于WD40子域的底部[6]。而And-1羧基末端的HMG框结构域晶体结构目前还未被报道。
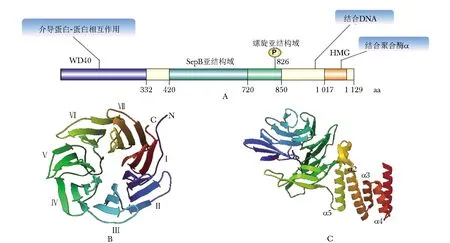
A.And-1结构域成分概述; B.WD40结构域(左,PDB ID:5GVA); C.SepB结构域(右,PDB ID:5GVB); aa.氨基酸图1 And-1蛋白结构示意图Fig 1 Structure diagram of And-1
2 And-1的结构功能
WD40结构域是真核基因组中最丰富、相互作用最强的结构域之一,其介导蛋白质与蛋白质或蛋白质与DNA的相互作用,并协调下游泛素化、组蛋白甲基化等作用。WD40结构域通常由7个β螺旋结构共同组成圆形结构域,该结构域中心具有一定的大小空腔,可以与药物小分子以较高的亲和力进行结合,介导参与关键伴侣蛋白的相互作用。此外WD40结构域的侧面和底部也可作为小分子药物结合位点,从而参与一系列生理过程[7]。以WD40结构域的蛋白作为药物靶点开发新药目前已有报道,如WDR5、EED蛋白。
SepB结构域参与介导And-1三聚体的形成。该结构域的螺旋束部分是聚合酶α相互作用的结合位点。酿酒酵母Ctf4的结构中,螺旋H2和 H4之间的半闭合槽可为酵母Pol催化亚基(Pol1)提供结合位点,二者之间的相互作用是由Pol1的CIP基序介导[8]。And-1与Polα的结合与Ctf4不同,其主要通过And-1的C端区域与Polα的特异性B亚基相互作用[6],而与Polα的CIP基序相互作用很弱。
HMG蛋白属于DNA结合特征家族。HMG-box/HMGB是由Box A N末端结构域、Box B中央结构域和酸性C末端结构域共同组成特征性结构域[9]。研究显示,Box A结构域结合DNA,Box B结构域结合并增强DNA弯曲,而C末端尾巴对DNA结合和弯曲特性起调节作用[10-11]。HMG蛋白通过调节DNA结合和弯曲活性来控制DNA复制、转录和修复的过程,从而对染色质结构和基因的转录产生影响[12]。此外,另有研究表明,And-1与DNA的结合由SepB和HMG之间的中间区域介导。推测这可能是因Polα与AND-1′C末端之间的相互作用,从而影响And-1的HMG结构域与DNA结合。
3 And-1的生化功能(图2)

图2 And-1生物功能示意图Fig 2 Schematic diagram of And-1 biological function
3.1 参与Pre-RC的组装
真核DNA的复制开始之前包括复制前复合物(pre-replication complex,pre-RC)的组装、激活和形成复制体等多步骤准备过程。And-1首先参与复制起点处的标记蛋白六亚基起源识别复合物(origin recognition complex,ORC)的组装。随后,加载微小染色体维持蛋白(minichromosome maintenance proteins,MCM)复合体的关键蛋白Cdc6和Cdt1,在G1期与ORC结合[13]。HBO1是一种H4特异性组蛋白乙酰化酶,对于Cdt1的复制许可至关重要[14]。然而And-1作为MCM2-7负载在染色质上的关键元素,可以通过独立于HBO1的方式促进Cdt1和MCM2-7之间的相互作用。加载MCM之后,And-1将Cdc45和GINS加载到pre-RC上,有助于CMG(Cdc45-MCM-GINS)复合体的形成。
3.2 参与 DNA复制
在高等真核生物中CMG复合物的形成需要SepB和HMG结构域的参与,And-1可与DNA聚合酶α、CMG解旋酶复合物相互作用,桥接CMG解旋酶与DNA聚合酶α,从而协调DNA解链和聚合酶活性,调控DNA复制[6]。缺少HMG结构域的Ctf4只能依赖于SepB结构域与复制体进展复合物(replisome promoting complex,RPC)[15]。研究表明,Ctf4和RPC之间的相互作用主要取决于Ctf4与GINS的相互作用,而GINS是RPC中存在的CMG复合物,在DNA复制过程中,Ctf4协调CMG复合物与Polα作用[16]。因此DNA复制需要CMG复合物参与,而细胞中CMG复合物的成熟需要And-1或Ctf4, 其在DNA复制中发挥重要的桥梁纽带作用[17]。
3.3 参与DNA 修复
DNA双链断裂(dobule strand break,DSB)是最致命的DNA损伤类型,可能导致有害基因的产生和染色体重排, 从而诱导癌发生。DNA双链损伤修复主要包含非同源末端连接(non-homologous end joining, NHEJ)和同源重组(homologous recombination,HR)两种方式[18]。And-1通过调节DNA末端切除来参与同源重组[19],并且And-1在同源重组过程中担负着双重职责,一种是将CtIP招募至DSB部位,另一种是在具有复制压力的细胞中参与Chk1激活。在HR修复方式中,肿瘤抑制蛋白CtIP对于DSB末端切除必不可少,并以MRN(MER11-RAD50-NBS1)依赖的方式被招募到DSB部位[20]。And-1通过与CtIP以及其他修复蛋白形成复合物,招募CtIP到DSB位点上调节DSB末端切除。此外,将And-1招募到DSB位点还依赖于BRCA1、MDC1和ATM(Ataxia telangectasia mutated)。研究表明,And-1与BRCA1和MDC1存在相互作用,而BRCA1的BRCT(BRCA1 C-terminal domain)结构域对于And-1的再生至关重要,但并不直接结合And-1[19]。
在DSB诱导后,依赖MRN/ATM的DSB末端切除导致形成ssDNA区域,该区域促进ATR激活并通过ATR引起Chk1磷酸化。在没有And-1或CtIP的情况下,Chk1的磷酸化降低且DSB末端切除受损。And-1在T826处被ATR磷酸化,这是And-1-Claspin相互作用对复制压力的反应所必需的,但其在T826处的磷酸化对And-1招募或DSB末端切除没有影响。Claspin是一种与Chk1相互作用的蛋白,可通过蛋白与蛋白之间的相互作用将Chk1带入停滞的复制叉,以使其被ATR磷酸化。ATR磷酸化And-1是响应复制应激中Chk1激活的关键[21]。
3.4 参与姐妹染色单体的内聚
姐妹染色单体的聚集对于减数分裂和有丝分裂中的染色体分离至关重要。研究显示,减数分裂中姐妹染色体分离之前,And-1耗尽的细胞停滞在前中期和中期,其对姐妹染色体的正确分离至关重要。在G1早期,着丝粒蛋白A(CENP-A)的特异性沉积需要And-1与CENP-A相互作用,And-1介导CENP-A与Holliday连接识别蛋白(Holliday junction recognition protein, HJURP)在着丝粒定位。And-1的下调能导致有丝分裂早期细胞的积累,并造成染色体收缩的缺陷;And-1过表达能增强着丝粒处的CENP-A装配。因此,And-1与HJURP一起作用,促进CENP-A向着丝粒的细胞周期特异性募集[22]。
4 And-1与癌
And-1在许多肿瘤细胞中呈过表达,尤其是在睾丸癌、恶性淋巴瘤、尿路上皮癌和恶性黑色素瘤,而在肾癌以及大多数肝癌和胰腺癌均为阴性。AKT1激酶磷酸化后能稳定癌细胞中的And-1蛋白,抑制PI3K/AKT通路会降低肺癌中And-1磷酸化的水平,提示其可能是参与癌症的AKT下游分子。选择性靶向AKT1和And-1之间的功能性相互作用,极有可能是一种有前景的癌症治疗策略[23]。研究也发现,And-1能通过促进MAPRE2在肺腺癌中的泛素化而导致顺铂耐药[24]。敲除And-1基因的细胞中添加细胞周期特异性化疗药物,如依托泊苷、PARP抑制剂AZD2281,均可有效提高药物的敏感性[25]。以上种种证据表明,And-1与潜在耐药性密切相关,选择性降解And-1,可以显著提高许多抗癌药物在癌中的治疗作用。
5 问题与展望
And-1是近年来新发现的DNA结合蛋白,在多种肿瘤细胞中过表达,并且参与DNA复制、转录和损伤修复等遗传相关的生命过程。目前人们对于该靶点的研究仍然处于刚刚起步阶段,And-1的功能机制仍需进一步探索和完善,相关小分子抑制剂的开发也需进一步探索和开发。本课题组研究发现了一类化合物直接作用And-1,抑制DNA损伤修复,在抗肿瘤和肿瘤化放疗增敏中起作用。本文对And-1的结构特点及功能进行了详细的综述,该蛋白WD40结构域的中心空腔口袋可作为小分子药物的主要靶向结合区,通过选择抑制蛋白与蛋白相互作用,发挥重要的调控功能。积极深入研究And-1在DNA遗传和损伤修复的作用机制,有助于广谱抗癌机制的药物发现;联合其他靶点的抗肿瘤药物,能协同增强抗肿瘤作用,并解决肿瘤药物耐药等难题。对该靶点药物的研发将是一个新颖且极具发展前景的抗肿瘤策略。
