巴哈雀稗苗期对不同浓度磷胁迫的生理适应机制
2021-05-07黄莉娟王普昶赵丽丽
刘 芳, 黄莉娟, 王普昶, 赵丽丽,3*
(1.贵州大学动物科学学院草业科学系, 贵州 贵阳 550025; 2.贵州省草业研究所, 贵州 贵阳 550006; 3.国家喀斯特石漠化防治工程技术研究中心,贵州 贵阳 550001)
巴哈雀稗(Paspalumnotatum)为禾本科雀稗属的多年生草本,耐贫瘠、适应性强;不仅营养丰富、饲用价值优良,还被人们用于水土保持、防风固沙、改良土壤理化性质和生态环境的防护[18-19]。其“一草多用”的功能对贵州发展畜牧业和保持水土流失具有重要作用。目前对巴哈雀稗的研究较少,国内的研究主要集中在盐和干旱的胁迫[20],种子休眠机理及破除方法[21];国外主要集中在致敏蛋白[22]、泡状丛枝菌根真菌的研究[23],而关于其耐低磷的研究较少。贵州磷素的缺乏限制了巴哈雀稗的推广种植,因此,本文以砂培方式人工模拟不同供磷水平,探究巴哈雀稗在磷胁迫下的根系形态、叶片形态和生理生化的适应性变化,发掘巴哈雀稗生长的适宜磷浓度,对减少土壤有限磷养分的消耗、提高磷的利用率以及从生物学途径缓解因土壤缺磷或施磷过多富集对巴哈雀稗生长的不利影响具有重要意义[24]。
1 材料与方法
1.1 试验材料与设计
选取健康饱满的巴哈雀稗种子,用75%的乙醇消毒5 min,然后用蒸馏水冲洗干净,置于铺满2层滤纸的培养皿中,放入恒温恒湿培养箱中25℃恒温培养,待出苗7 d后,选取整齐、发育一致的巴哈雀稗苗移栽到石英砂塑料盆中(长26 cm、宽12 cm、高15 cm),在1/2全营养液培养15 d后全营养液再培养10 d,待其长至三叶一心时进行胁迫处理。供试营养液为改良的Hoagland与阿农微量元素混合液[25],营养液配方为:KH2PO40.2 mmol·L-1,K2SO40.75 mmol·L-1,MgSO4·7H2O 0.65 mmol·L-1,Ca(NO3)2·4H2O 2 mmol·L-1,EDTA-Fe 0.1 mmol·L-1,H3BO41.0×10-3mmol·L-1,MnSO4·H2O 1×10-3mmol·L-1,ZnSO4·7H2O 1.0×10-3mmol·L-1,CuSO4·5H2O 0.5×10-3mmol·L-1,(NH4)6MO7O24·4H2O 0.05×10-4mmol·L-1,营养液pH值为6.20。全营养液中的磷(KH2PO4)水平分别设置2 μmol·L-1(P1),20 μmol·L-1(P2),200 μmol·L-1(P3),600 μmol·L-1(P4),1 000 μmol·L-1(P5)5个处理,每个磷浓度3盆,共移栽15盆,每盆22株。利用K2SO4调节营养液中K+浓度,使K+浓度维持在1 700 μmol·L-1。在胁迫10 d,20 d和30 d时测量生理指标,试验30 d结束时,分别测量地上和地下的形态指标。
1.2 试验方法
1.2.1植株形态指标 叶形、株高、生物量、根系,每个指标10次重复。其中,叶形指标用叶片扫描仪测量叶长、叶宽、叶长宽比及叶面积,将扫描后的图片用Win RHIZO软件进行分析。株高用直尺量取地上部分拉直后的植株高度。生物量测定是取出整植株,将地上部分和地下部分分离后,分别称量地上和地下鲜重,之后置于105℃杀青30 min,65℃烘干至恒重测其干重。根系指标测定是用ScanMaker 9800XL plus扫描根系,获取总根长、根表面积、根体积、根平均直径、根尖数、根分枝数。
1.2.2生理指标 取第2~3片叶片,进行各生理指标的测量,每个指标3次重复,叶绿素(chlorophy,Chl)采用丙酮-乙醇混合液法[26]测定;电导率(conductivity)采用浸泡法[27]测定;可溶性糖(soluble sugar,SS)含量采用蒽酮比色法[28]测定;脯氨酸(proline,PRO)用酸性茚三酮比色法[29]测定;丙二醛(malonaldehyde,MDA)采用硫代巴比妥酸法[30]测定;可溶性蛋白(soluble protein,SP)采用双缩脲法[31]测定;过氧化物酶(peroxidase,POD)用活性愈创木酚法[32]测定;过氧化氢酶(catalase,CAT)用紫外分光光度法[33]测定;超氧化物歧化酶(superoxide dismutase,SOD)用氮蓝四唑光还原法[34]测定;根部酸性磷酸酶活性(root acid phosphatase,ACP)选取根部,采用对硝基苯磷酸二钠法测定[35]。
1.3 试验数据处理与分析
用Microsoft Excel 2010进行图表的绘制,采用SPSS 20统计软件进行单因素方差分析,采用Duncan方法进行多重比较。参照郑清岭等[36]的研究,采用主成分分析结合模糊隶属函数公式对试验处理效果进行综合分析。计算公式如下:
μ(Xi)=(Xi-Ximin)/(Ximax-Ximin)(i=1,2,3)
(1)
(2)
(3)
其中,μ(Xi)为各处理Xi的隶属函数值,Xi为各处理综合指标i的值,Ximax和Ximin分别为各处理Xi的最大值和最小值,Pi为综合指标i的贡献率,Wi为综合指标i的权重,D为各处理的综合评价值。
2 结果与分析
2.1 磷对巴哈雀稗形态特征的影响
2.1.1根系形态特征 根系是植株在土壤中吸收磷素的重要部位,植株体内的磷素很大程度上取决于根系吸收磷的含量。根系形态在5个不同供磷水平下的差异明显。由表1可知,在低磷或者高磷水平,根表面积、总根长、总根体积、根分枝数和根尖数均有不同程度的下降,且均在P2水平值最大,与各个处理间差异显著(P<0.05);而根平均直径随着供磷水平的升高而升高,在P5磷水平直径最大。
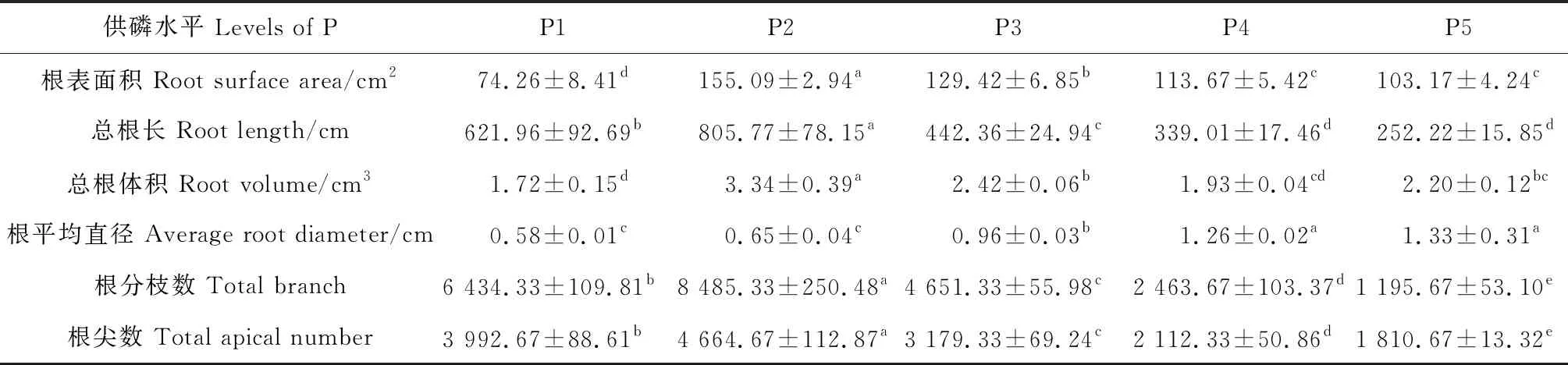
表1 不同供磷水平对巴哈雀稗根形态的影响
2.1.2叶片形态特征 如表2所示,随着供磷水平的升高,叶面积、叶长、叶宽、叶周长均呈现先升高再降低的趋势,在P4磷水平达到最大值,P1水平下最低。在P1,P2磷水平处理间各个指标差异不显著;在P2和P3磷水平处理间叶面积、叶长、叶宽的差异显著(P<0.05);在P3和P4磷水平处理间叶长、叶周长差异显著(P<0.05);在P4和P5磷水平处理间叶面积、叶长、叶宽、叶周长差异显著(P<0.05);长宽比和叶形系数在各个处理间差异不显著。

表2 不同供磷水平对巴哈雀稗叶片形态的影响
2.1.3生物量与根冠比 生物量与根冠比可以反映植物生长发育的情况。由表3可知,随供磷水平的提高,茎叶鲜重和干重呈现先上升再下降的趋势,在P4磷水平为最大值,显著高于其他供磷水平(P<0.05);根系鲜重和干重在P2磷水平下为最高值,且显著高于其他供磷水平(P<0.05);鲜重根冠比和干重根冠比均呈现先上升再下降的趋势,在P2磷水平为最大值,显著高于其他供磷水平(P<0.05);株高在P4磷水平为最大值,高于其他供磷水平。

表3 不同供磷水平对巴哈雀稗株高及生物量的影响
2.2 磷对巴哈雀稗生理特性的影响
2.2.1脯氨酸(Pro)、可溶性蛋白(SP)、可溶性糖(SS)、叶绿素(Chl)含量 脯氨酸、可溶性蛋白和可溶性糖是植物体内的渗透调节物质,在同一供磷水平下的含量均随胁迫时间的持续呈现上升趋势。由图1可知,在10 d,20 d,30 d时,脯氨酸含量均随着供磷水平的升高而降低,在P1供磷水平下最高(5.96 g·mL-1,14.34 g·mL-1,30.56 g·mL-1),在P5供磷水平时最低(1.44 g·mL-1,1.73 g·mL-1,17.55 g·mL-1)。在10 d,20 d,30 d时,可溶性蛋白和可溶性糖含量均随供磷水平的升高呈现先升高后降低的趋势,在P3,P4磷水平高于其他磷水平,P1磷水平时最低。在磷胁迫的3个时期,叶绿素的含量呈现先升高后降低的趋势,且均在P4磷水平时值最大、P1磷水平含量最低,叶绿素含量排序为:P4>P3>P2>P5>P1。
由此可见,磷元素的丰缺影响植株脯氨酸、可溶性蛋白、可溶性糖和叶绿素的合成。

图1 不同供磷水平对脯氨酸、可溶性糖、可溶性蛋白和叶绿素含量的影响
2.2.2丙二醛(MDA)含量及电导率 丙二醛和电导率反映逆境对细胞膜的伤害程度。由图2可知,在10 d,20 d,30 d时,丙二醛和电导率的含量随着供磷水平的升高呈现先下降后升高的趋势,P3,P4磷水平处的电导率显著低于其他3个磷水平(P<0.05),且随着胁迫时间的延长,丙二醛含量变化呈升高趋势,表明过低或者过高磷浓度会破坏细胞膜,对细胞膜的伤害程度随胁迫时间的延长而增加。
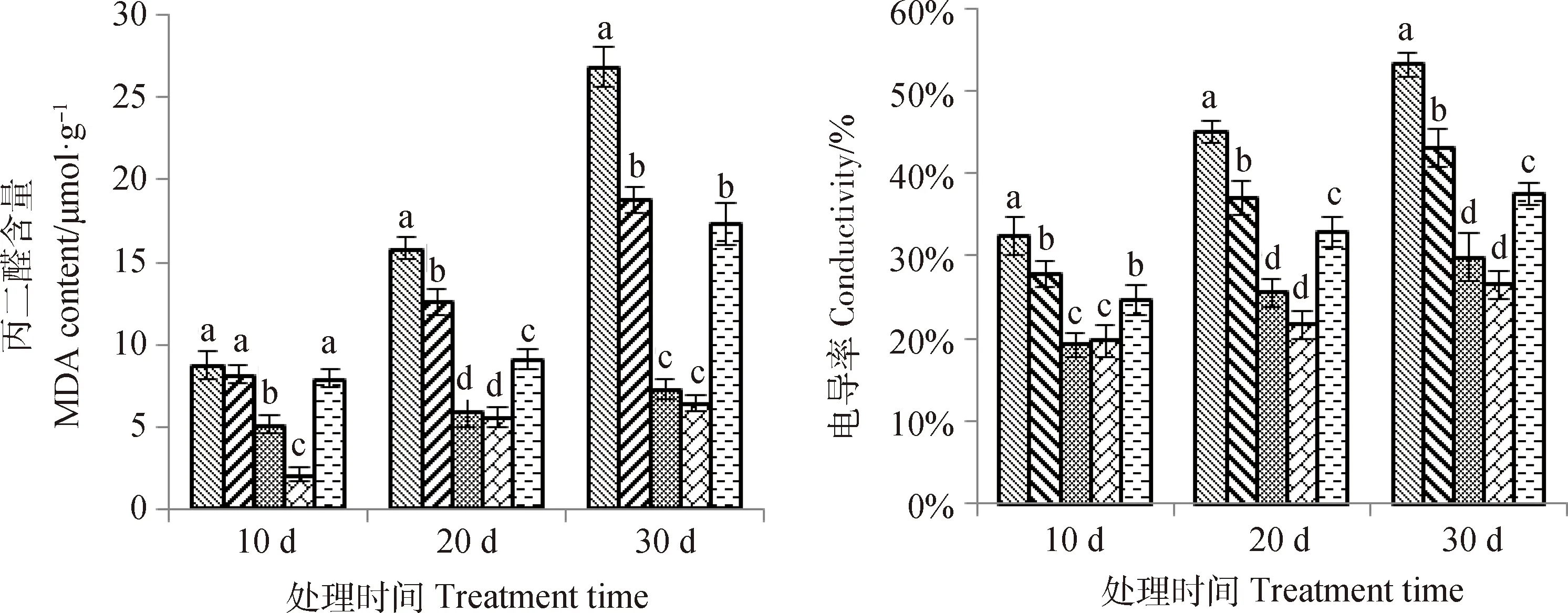
图2 不同供磷水平对丙二醛和电导率的影响
2.2.3酶活性 由图3可知,过氧化氢酶(CAT)、过氧化物酶(POD)、超氧化物歧化酶(SOD)的活性在3个时期变化趋势大致相同,均随供磷水平的增加表现先降低后升高的趋势,且随着胁迫时间的持续酶活性升高。CAT在P1,P2处活性较高,在P3,P4磷水平下较低。POD和SOD在高于或者低于P4磷水平呈上升趋势,POD在P4磷水平时活性最低,SOD在P3,P4磷水平时较低。根系分泌的酸性磷酸酶(ACP)随着供磷水平的升高呈现下降趋势,ACP活性最高和最低分别是P1和P5磷水平。综上分析,P3,P4磷水平下巴哈雀稗的自由基较少,需要的酶活性较低。
2.3 巴哈雀稗适应不同供磷水平的综合评价
单一指标不能反映巴哈雀稗的对不同磷浓度适应的积极性,主成分分析是利用降维的方法,将多个变量简化为较少的变量,不仅简化了数据,又体现了变量之间的相关性。为了探究不同供磷水平对巴哈雀稗形态变异和生理影响的总效应,采用主成分分析法对第30 d的各形态、生理共27个指标进行了主成分提取,将27个指标转化为3个新的综合指标,提取的3个主成分的特征值均大于1,累积贡献率达93.43%,对数据的变异具有较高的解释。其中主成分1(PC1)到主成分3(PC3)分别解释了57.05%,22.25%,14.13%的变异。第1主成分中,叶片鲜重、叶片干重、CHL,SS的载荷值相对较大。第2主成分中,Pro、根分枝数、根尖数、ACP的载荷值相对较大。第3主成分中,根系鲜重、总根体积、根总面积、根系干重的载荷值相对较大,是对磷元素供给敏感的指标(表4)。

图3 不同供磷水平对保护酶(SOD,POD,CAT,ACP)活性的影响
由主成分分析结果可得3个综合指标的线性方程组。其中,X1,X2,X3分别代表了3个主成分,以各指标的载荷值为系数,x1—x27分别代表总根长、根表面积、总根体积、根平均直径、根尖数、根分枝数、叶面积、叶长、叶宽、叶长宽比、叶周长、叶形系数、株高、茎叶鲜重、根系鲜重、茎叶干重、根系干重、CAT,POD,SOD,Pro,SS,SP,MDA,CHL、电导率、ACP的27个指标,分别用5个供磷水平下的各个指标的平均值乘以相应指标的系数,得到关于5个供磷水平下的3个主成分的综合指标值(表5)。
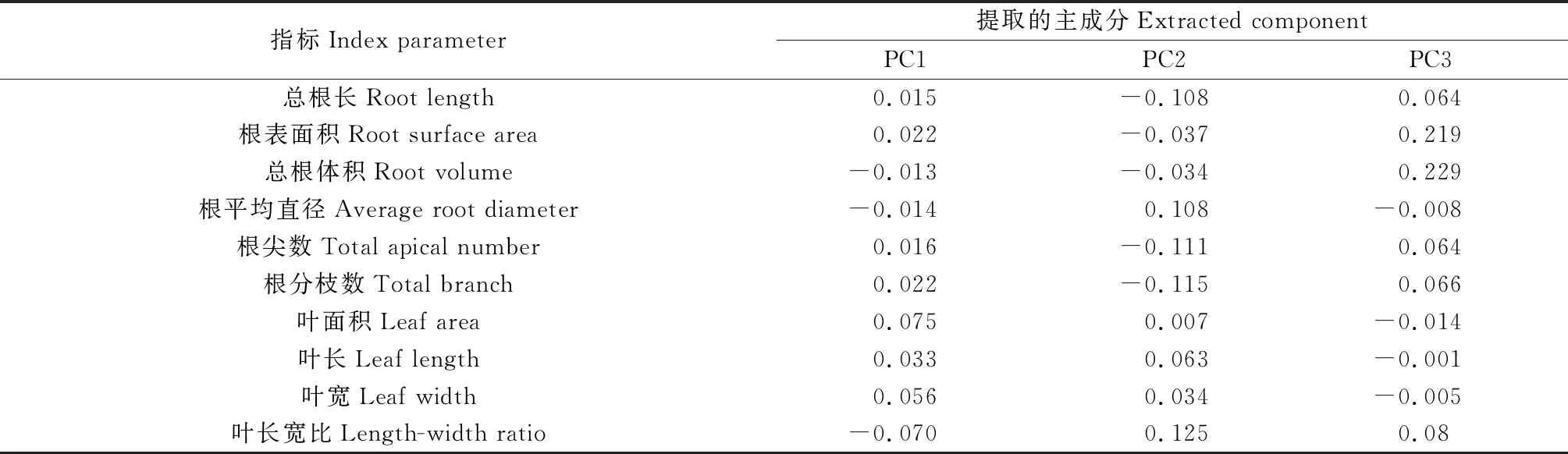
表4 主成分载荷矩阵及贡献率

续表4
X1=0.015x1+0.022x2-0.013x3-0.014x4+0.016x5+0.022x6+0.075x7+0.033x8+0.056x9-0.07x10+0.074x11+0.033x12+0.111x13+0.099x14+0.007x15+0.095x16+0.041x17+0.013x18+0.039x19+0.073x20-0.073x21+0.091x22+0.076x23+0.086x24+0.092x25+0.067x26-0.025x27。
X2=-0.108x1-0.037x2-0.034x3+0.108x4-0.111x5-0.115x6+0.007x7+0.063x8+0.034x9+0.125x10+0.000x11+0.038x12-0.081x13-0.064x14-0.012x15-0.043x16-0.047x17+0.076x18+0.052x19+0.012x20+0.136x21-0.034x22-0.020x23-0.013x24-0.037x25+0.017x26+0.11x27。
X3=0.064x1+0.219x2+0.229x3-0.008x4+0.064x5+0.066x6-0.014x7-0.001x8-0.005x9+0.08x10-0.065x11+0.067x12-0.009x13-0.04x14-0.237x15-0.025x16-0.218x17+0.062x18-0.007x19-0.035x20+0.042x21+0.028x22+0.074x23-0.009x24+0.032x25+0.010x26+0.065x27。
通过模糊隶属函数公式求出不同供磷水平对巴哈雀稗的模糊隶属函数值、各指标的权重以及综合评价值。综合评价值越大表明对巴哈雀稗的生长作用越强,结果表明,P4的综合评价值最大(0.785),综合排名为第1,表明P4磷水平对巴哈雀稗的生长最有利(表5)。

表5 不同供磷水平对巴哈雀稗幼苗形态和生理指标的隶属函数值
3 讨论
3.1 巴哈雀稗对不同供磷水平的形态适应
植物吸收磷的效率与供磷水平有关,即吸收效率随供磷量的增加而升高,为了适应不同磷环境,植物在进化过程中形成了包括根系形态、叶片形态和生理生化等一系列复杂的适应性调节机制。
巴哈雀稗的根系形态和叶片形态对不同供磷水平有不同的响应,根是感受外源磷变化进而调节根尖分生组织的部位,接收并传递胁迫信号后作出反应,再反馈调节作用于根部发生适应性调整。磷饥饿时,分生组织细胞减少[37],根毛通过增加根表面积和以耗费最小的代谢能吸收磷而获取竞争优势[37-38]。巴哈雀稗在P2磷水平下的根系生物量最大,根表面积、总根长、总根体积、根分枝数和根尖数明显高于其他水平,随着供磷水平的上升,以上根系指标均逐渐变小,说明在较低的磷水平下(P2),巴哈雀稗为了吸收更多的磷营养,把吸收到的养分更多用于发展根部,通过缩小根直径,增加侧根、丛生根、延长根系长度,形成根系网络,扩大根部与土壤的接触面积,进而促进根尖分生区对磷素的吸收。这与蔡丽平等对类芦[39]、张恩和等对春蚕豆[40]的研究结果一致。而在P1磷水平下的根系发育反而比P2磷水平差,可能是因为供磷水平过低,植物生理代谢无法调节造成根系发育受阻。株高和茎叶的鲜重、干重随磷水平的升高而先增加后降小,在P4磷水平发育最好。有研究表明,紫花苜蓿在低磷胁迫的根冠比中磷要高[41]。本文研究中根冠比不论是鲜重还是干重均在P2,P3磷水平较高,P4,P5磷水平较低,这一现象又进一步验证了在较低磷水平,植物会通过调节地上和根系生物量的生长,增强根系的发育来适应供磷不足的逆境。
3.2 巴哈雀稗对不同供磷水平的生理适应
在正常情况下,细胞内自由基的产生和清除处于动态平衡状态,当植物受到胁迫打破平衡时,逆境会加速植物组织中膜脂过氧化链式反应,导致自由基(O2-)、羟基自由基(OH-)、过氧化氢(H2O2)、单线态氧(O2)等超氧化物阴离子积累过多造成一系列生理生化紊乱。MDA不仅能与细胞内各种物质发生强烈反应,造成细胞功能不可修复的丧失和不正常的死亡,还会破坏细胞中许多生物功能分子,比如蛋白质、核酸等[42]。本研究中,巴哈雀稗在P1,P2和P5供磷水平下3个时期的电导率和MDA含量均较高,表明细胞膜脂过氧化程度更高,膜透性遭到破坏导致细胞液外渗。前人研究表明,磷的缺乏或者过剩导致蛋白质合成代谢受阻[43-44],可溶性糖含量减少[45]。在本研究中,3个胁迫时期下,巴哈雀稗在较低或较高的磷水平时叶绿素合成受阻,合成有机物的速率减慢,相应的SP和SS的合成也减少。原因可能是巴哈雀稗在磷素缺乏或者过多时产生了伤害,抑制可溶性糖和可溶性蛋白的合成。为保持不同磷环境的渗透势,除了可溶性糖、蛋白质以外,植物会调节体内脯氨酸的含量。脯氨酸是最有效的渗透调节物质,在逆境条件下对植物可能起到兼容性渗透调节剂、蛋白质和亚细胞结构稳定剂等作用[46]。3个胁迫时期下,在较低磷水平(P1,P2),巴哈雀稗通过促进自身合成脯氨酸来降低渗透势进而吸收更多的磷,而随着磷浓度的升高,介质中能吸收到的磷元素增多,脯氨酸合成减少以稳定渗透势保持膜系统的完整。由此可见,在P3,P4磷水平更适合巴哈雀稗的生长发育;在较低或者较高磷水平不利于细胞内叶绿素、可溶性糖、可溶性蛋白的合成积累;巴哈雀稗通过调节可溶性糖、可溶性蛋白及脯氨酸的含量来共同维持细胞渗透势以适应外部磷水平变化的环境。
磷胁迫不仅能升高细胞活性氧水平,还能诱导植物保护系统的建立[47],POD,SOD,CAT这些保护酶清除细胞内产生的活性氧,维持细胞功能正常发挥[37]。POD具有消除过氧化氢和酚类、胺类毒性的双重作用,在低磷水平(P1)和高磷水平(P5)下巴哈雀稗POD酶活性显著增强以分解H2O2(图3)。超氧化物歧化酶(SOD)可以与产生过量的自由基发生反应,产生H2O2和O2,在低磷水平(P1)和高磷水平(P5)显著增强(图3)。H2O2主要由CAT清除,最后分解形成H2O和O2,在低磷水平(P1、P2)的巴哈雀稗CAT酶活性显著增强(图3),且活性远远高于POD,SOD活性,原因可能是CAT在磷胁迫中起着最为重要的保护作用,细胞中通过SOD途径产生的和本身存在于细胞内的H2O2都需要CAT的清除。随着自由基含量下降,保护酶活性也下降,总体来说,在P3,P4磷水平时各种酶活性较低,表明需要消除的有害物质较少,活性氧物质通过过氧化氢酶、过氧化物酶、超氧化物歧化酶和各种还原酶等共同的作用而清除[47]。根系内源酸性磷酸酶活性增强促进根系对周围磷的活化、增加有效磷进入细胞[48-49]。巴哈雀稗ACP随磷水平增加而降低,说明在磷水平低的情况下,巴哈雀稗增强酸性磷酸酶的活性,活化土壤中难溶性磷进而加快磷素代谢来弥补磷缺乏,提高磷利用效率,而高磷水平则抑制巴哈雀稗根部酸性磷酸酶的分泌。
3.3 巴哈雀稗适应不同供磷水平的综合评价
巴哈雀稗对不同供磷水平的适应是一个复杂的反应,从形态、生理的某个指标不能反映巴哈雀稗的生长情况,故利用主成分分析和模糊隶属函数结合的方法对不同供磷水平下巴哈雀稗幼苗各指标进行综合评价。由表5可知,各磷水平胁迫处理下巴哈雀稗幼苗各生长和生理指标的综合评价值大小为P4>P3>P5>P2>P1,其中P4,P3综合评价值较高,表明在P4,P3磷水平较适合巴哈雀稗的生长。P5和P2磷水平比P4,P3磷水平综合得分低,并且得分相近,这表明在高磷(P5)和低磷(P2)胁迫下均影响了巴哈的生长发育;高磷水平(P5)所提供的磷浓度能够满足巴哈雀稗对磷的需求,而较低磷水平(P2)下巴哈雀稗通过根系形态的变化改变对磷的吸收来提高磷的吸收效率促进生长,进而与P5高磷水平生长发育相当。
4 结论
在低磷条件下,巴哈雀稗通过减少叶面积、叶长、叶宽、叶周长、茎叶干重、鲜重等地上部分生物量,增加根面积、总根长、根分枝数和根尖数等地下部分生物量的比例来适应磷缺乏的逆境。在较低或者较高的磷水平,膜透性遭到破坏,叶绿素合成受阻,可溶性蛋白和可溶性糖减少;脯氨酸与可溶性糖、可溶性蛋白的含量发生不同的变化共同维持细胞内环境的稳定;保护酶活性升高以清除细胞内过量的ROS物质。综合评价的结果得出600 μmol·L-1(P4)磷水平最适合巴哈雀稗的生长;相比叶片形态和生理上的差异,各个供磷水平间根系有较大的差异性,根系发育良好为巴哈雀稗竞争吸收到更多的磷提供了重要的优势。
