寄主对云斑白条天牛幼虫取食行为及其体内酶活性的影响
2021-05-03李贝娜彭思颖郭春晖杨振德
吴 广 徐 蕾 李贝娜 彭思颖 郭春晖 杨振德
(广西大学林学院,广西 南宁 530004)
云斑白条天牛(Batocera lineolata)属鞘翅目(Coleoptera)天牛科(Cerambycidae),是一种寄主范围广、适应性强的蛀干害虫[1]。它可危害100多种阔叶林和果树[2-4],是高度危险的林业害虫[5]。
云斑白条天牛成虫在补充营养期具有转换寄主补充营养,然后回到原寄主产卵的习性,如杨树(Populussp.)[6-8]和 桉 树(Eucalyptussp.)[9]的云斑白条天牛种群。但以核桃(Juglans regia)[4]、栎树(Quercus linn)[10]和白蜡树(Fraxinus chinensis)[11-12]等为寄主的云斑白条天牛,补充营养和产卵阶段均在同一寄主上进行。云斑白条天牛对于不同寄主表现出习性差异,与其较强的转换寄主和代谢适应能力有密切关系,这对于云斑白条天牛种群的扩散具有重要意义。云斑白条天牛适应新寄主时,天牛在生理生化方面的变化尤为明显。云斑白条天牛成虫以嗜食寄主蔷薇(Rosa multiflora)为补充营养源时,谷胱甘肽-S-转移酶(GSTs)活性较低;而当取食非嗜食寄主杨树、柳树(Salix babylonica)和构树(Broussonetia papyrifera)时,谷胱甘肽-S-转移酶的活性较高[13]。云斑白条天牛成虫阶段是其唯一转换寄主选择阶段[4]。当雌成虫产卵于新寄主中,新孵化的幼虫主要通过生理生化上的变化来适应新寄主,从而生长发育成虫,逐渐形成新的寄主种群,实现种群的分化和扩散。目前对云斑白条天牛的研究主要集中在成虫方面,关于云斑白条天牛寄主谱扩大的内在机制尚不清楚,并且不同寄主对云斑白条天牛幼虫取食行为和酶活性的影响也研究较少。
本研究以核桃树和桉树为原寄主的云斑白条天牛种群幼虫为研究对象,通过人工更换新寄主饲养的方法研究云斑白条天牛幼虫对核桃和桉树的取食选择行为,通过测定更换新寄主前后幼虫体内消化酶、解毒酶和保护酶活性变化探讨其对核桃或桉树不同寄主的适应机制,以利于预估云斑白条天牛在核桃和桉树间的潜在危险性,也有助于研究针对云斑白条天牛种群扩散的防治手段,为云斑白条天牛的有效防治提供参考。
1 材料与方法
1.1 幼虫来源及其饲养
分别于广西壮族自治区河池市巴马县核桃林和百色市那坡县桉树林中采集云斑白条天牛的初孵幼虫或卵粒,卵粒置于垫有湿润滤纸的培养皿中,在光照培养箱(温度27 ℃,光照12 h/d,相对湿度80%~85%)中培养孵化。初孵幼虫分别采用原寄主木段饲养法[14]饲养3 d和14 d后备用。寄主植物桉树和核桃木段均采自广西大学林学院校内教学实践基地。
1.2 幼虫对不同寄主取食选择行为的测定
取食选择行为测定装置由直径9.0 cm的两格或三格一次性培养皿改造而成。为了便于幼虫的爬行,测定时先在培养皿底部垫一层滤纸,再将新鲜的不同饲料放入不同的区域,然后取1头预饥饿2 h的幼虫放入装置中心区(见图1),置于暗处2 h后观察记录幼虫对不同食物的选择情况。每种类型幼虫的选择试验20次重复,计算趋向率,其公式为:


图 1 云斑白条天牛幼虫取食选择行为测定装置示意图Fig. 1 Schematic diagram of a device for determining the feeding behavior of the larva of B. lineolata
1.3 幼虫体内酶活性的测定
分别取不同寄主木段饲养14 d后的3龄幼虫,冰冻致死后迅速置于已灭菌4 ℃预冷的解剖盘上解剖中肠,清洗中肠内含物,滤纸吸干表面液体后称量备用。
幼虫中肠乙酰胆碱酯酶(AchE)、谷胱甘肽-S-转移酶(GST)、羧酸酯酶(CarE)、多酚氧化酶(PPO)等酶活性的测定均采用南京建成生物工程研究所生产的试剂盒(编号分别为A024、A004、A133和A136)说明书进行。
超氧化物歧化酶活性参照任晓亚等[15]的方法测定;过氧化氢酶和过氧化物酶活性参照李周直等[16]的方法测定;类胰蛋白酶活性参照李娟[13]的方法测定;淀粉酶活性参照周琼等[17]的方法测定;纤维素酶活性参照李晓娟[18]的方法测定;酶蛋白含量测定采用考马斯亮蓝法[19]测定。
以上酶活性单位采用相应参考文献中的定义。各种酶活性测定试验均6次以上重复。
1.4 数据分析方法
采用SPSS 22.0和Excel 2016进行数据统计与分析。利用卡方检验分析天牛幼虫选择某个寄主的最多次数和其他寄主选择次数间有无显著差异;采用Duncan"s极差法比较各组数据间的差异显著性(P<0.05),试验数据最终用平均值±标准误差来表示。
2 结果与分析
2.1 幼虫对寄主的取食选择
从表1可以看出:在选择性取食条件下,取食原寄主3 d后的天牛幼虫仍趋向于选择取食原寄主,其选择趋向率均达80%以上。将天牛幼虫强迫取食新寄主3 d后再进行选择性取食测试,结果发现幼虫选择寄主的能力下降,选择的趋向率均低于70%。其中,原寄主为桉树的幼虫强迫取食核桃寄主3 d后,明显倾向于选择取食桉树与核桃的混合饲料(P<0.05),而原寄主为核桃的幼虫强迫取食桉树寄主3 d后,对于各饲料取食的倾向性无明显差异(P>0.05)。

表 1 云斑白条天牛幼虫对不同寄主的取食选择性Table 1 Determination of the selective tropisms of different plants for the larvae of B. lineolata
2.2 取食不同寄主的幼虫体内解毒酶活性
从表2可以看出:原寄主为核桃的云斑白条天牛幼虫取食新寄主桉树14 d后,其体内乙酰胆碱酯酶活性显著升高,而谷胱甘肽-S-转移酶、羧酸酯酶和多酚氧化酶活性均显著降低(P<0.05)。天牛幼虫取食新植物桉树后的解毒酶活性中以多酚氧化酶活性变化最大,云斑白条天牛核桃种群幼虫取食原寄主核桃后的酶活性是取食桉树后酶活性的9.38倍。
原寄主为桉树的幼虫取食新寄主核桃后,其体内多酚氧化酶和谷胱甘肽-S-转移酶活性均显著降低(P<0.05),而乙酰胆碱酯酶活性则显著升高(P<0.05),羧酸酯酶活性无显著差异。
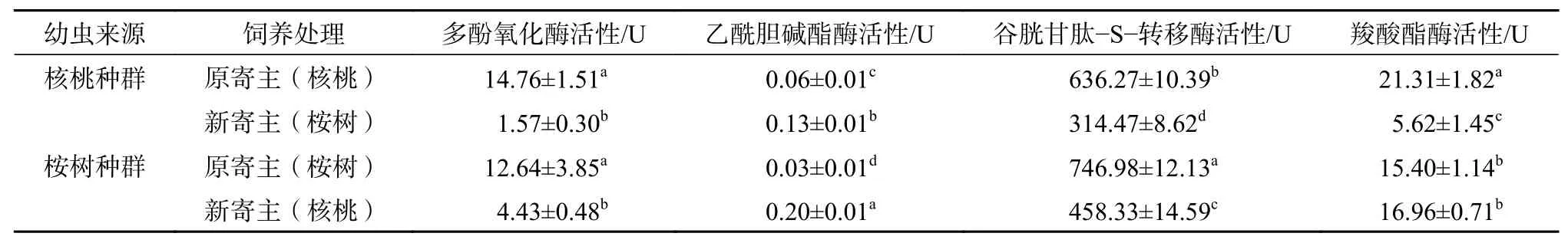
表 2 取食不同寄主的云斑白条天牛幼虫体内解毒酶活性的变化Table 2 Changes of detoxifying enzymes activity in larvae of B. lineolata fed with J. regia or Eucalyptus sp.
2.3 取食不同寄主的幼虫体内保护酶活性的影响
从表3可以看出:云斑白条天牛核桃种群幼虫转换桉树饲养14 d后,幼虫保护酶活性显著低于原寄主核桃饲养后的酶活性(P<0.05)。其中以过氧化氢酶(CAT)差异性最大,云斑白条天牛核桃种群幼虫的CAT酶活性是桉树种群幼虫酶活性的2.05倍。
云斑白条天牛桉树种群幼虫转换核桃饲养14 d后,幼虫保护酶活性除了CAT酶活性与原寄主桉树饲养后的CAT酶活性没有明显差别外,超氧化物歧化酶(SOD)和过氧化物酶(POD)活性分别与原寄主桉树饲养后的相应酶活性有显著差异(P<0.05),即原寄主桉树饲养后的SOD酶活性显著高于核桃饲养后的SOD酶活性,而POD酶活性显著低于核桃饲养后的POD酶活性(P<0.05)。云斑白条天牛在取食原寄主桉树14 d后,其中肠内的SOD酶活性是取食核桃树后SOD酶活性的4.80倍。

表 3 取食不同寄主的云斑白条天牛幼虫体内保护酶的活性变化Table 3 Changes of protective enzymes activity in larvae of B. lineolata fed with J. regia and Eucalyptus sp.
2.4 取食不同寄主的幼虫体内消化酶活性的影响
从表4可以看出:云斑白条天牛核桃种群幼虫取食原寄主核桃14 d后纤维素Cx酶活性和桉树种群幼虫取食原寄主桉树后纤维素Cx酶活性没有明显差别。虽然核桃种群幼虫取食核桃后的类胰蛋白酶活性明显低于桉树种群幼虫取食桉树后的类胰蛋白酶活性(P<0.05),但淀粉酶活性却显著高于桉树种群(P<0.05),是桉树种群的10.13倍。
转换植物取食后,除了云斑白条天牛桉树种群幼虫取食核桃后其体内淀粉酶活性与取食原寄主桉树后的淀粉酶活性无明显差别外,类胰蛋白酶和纤维素Cx酶均显著低于取食原寄主后的相应酶活性(P<0.05)。另外,云斑白条天牛核桃种群幼虫取食桉树后的类胰蛋白酶活性和桉树种群幼虫取食核桃树后类胰蛋白酶活性没有明显差异,但淀粉酶活性以及纤维素Cx酶均明显高于桉树种群幼虫相对应的消化酶活性(P<0.05)。

表 4 取食不同寄主的云斑白条天牛幼虫体内消化酶的活性变化Table 4 Changes of digestive enzymes activity in larvae of B. lineolata fed with J. regia and Eucalyptus sp.
3 结论与讨论
植食性昆虫在与植物长期的协同进化过程中,一方面植物为保护自身免受虫害而产生许多次生代谢产物进行化学防御[20];另一方面植食性昆虫通过解毒代谢、避毒行为等机制来不断适应植物体内的次生防御物质[21]。对于云斑白条天牛而言,幼虫无法选择寄主,主要依靠成虫选择新寄主产卵后初孵幼虫再进行新寄主的适应,建立新的寄主种群,从而扩大寄主范围[22]。
本研究结果表明云斑白条天牛幼虫偏好取食原寄主,但强迫幼虫取食新寄主能降低幼虫对原寄主取食的趋向率。这与阎雄飞在研究光肩星天牛对原寄主的“记忆”效应时,天牛并没有完全对原寄主表现出“记忆”效应相似[23]。原寄主为桉树的幼虫强迫取食核桃寄主3 d后,明显倾向于选择取食桉树与核桃为1∶1的混合饲料,这说明天牛的取食经历可能会改变天牛的取食嗜好,从而使云斑白条天牛幼虫对不同寄主的混合饲料具有较强的选择性。提示自然桉树林中的天牛成虫“误产卵”到邻近的核桃树上,其孵化的幼虫具有适应新寄主的潜力。
昆虫可以通过改变体内酶活性来适应寄主的变化,昆虫体内不同酶系的功能不同,不同酶对于新寄主环境的反应速度也不同。云斑白条天牛核桃种群和桉树种群幼虫更换新寄主后其体内解毒酶活性发生显著变化,乙酰胆碱酯酶活性显著升高,谷胱甘肽-S-转移酶和多酚氧化酶活性均显著降低,这表明乙酰胆碱酯酶可能是云斑白条天牛幼虫适应新寄主的关键解毒酶。
核桃种群幼虫更换新寄主桉树后,幼虫体内的三大保护酶(CAT、SOD、POD)活性均显著下降;但是,桉树种群幼虫更换新寄主核桃后,幼虫体内的CAT活性相对比较稳定,POD活性显著升高。此外,体内羧酸酯酶活性也略有升高,说明相对于更换新寄主的核桃种群幼虫,桉树种群幼虫具有对新寄主核桃更强的自我保护适应能力和解毒适应能力。
一般来说,昆虫的消化酶活性大小在一定程度上体现出昆虫正常生长发育时所需营养物质的种类以及寄主植物中各类营养物质被昆虫吸收利用的状况[24]。取食新寄主后,云斑白条天牛幼虫的类胰蛋白酶和纤维素Cx酶活性均明显降低,推测新寄主中可能含有抑制幼虫蛋白酶和纤维素酶的物质,抑制了酶活性,从而延缓了幼虫的生长发育。核桃种群幼虫更换新寄主桉树后,幼虫体内的淀粉酶活性显著降低,说明核桃种群幼虫较难适应新寄主桉树;但是,桉树种群幼虫更换新寄主核桃后,幼虫体内的淀粉酶活性无明显变化,且酶活性较低,说明淀粉酶在桉树种群幼虫适应新寄主过程中的作用较弱,即淀粉酶不是桉树种群幼虫寄主转换的主要限制因素。
天牛体内消化酶活性的变化,可在一定程度上反映出天牛对寄主植物营养物质的嗜好程度。在云斑白条天牛幼虫体内类蛋白酶、淀粉酶和纤维素Cx酶等三大消化酶中,核桃种群幼虫体内的淀粉酶活性显著高于桉树种群幼虫,但类胰蛋白酶活性却显著低于桉树种群幼虫。提示云斑白条天牛幼虫可能以不同消化酶活性构成模式适应不同的寄主,形成不同的寄主种群。此外,本研究只测定了更换新寄主14 d这个时间点云斑白条天牛幼虫体内酶活性的变化情况,随着时间的推移,酶活性会怎么变化还有待于进一步研究。
综上所述,云斑白条天牛幼虫对原寄主具有“记忆”效应,更换寄主对其体内酶活性产生明显影响,乙酰胆碱酯酶在适应新寄主中可能发挥重要作用。相对于核桃种群的天牛幼虫对新寄主桉树的适应,桉树种群幼虫具有对新寄主核桃更强的适应能力。提示桉树种群的云斑白条天牛具有较大的转移危害核桃树的风险,应注意桉树林和核桃经济林的间隔种植和防护,尤其是桉树林应当避免配置核桃树这种适宜寄主。
