10份小麦持久抗性资源和6份新品系抗条锈病的遗传特点分析
2021-04-30魏婷朱孟杰郎晓威王震顾紫琼潘伟釉王科宇贾湘冯晶王凤涛郭青云蔺瑞明
魏婷 朱孟杰 郎晓威 王震 顾紫琼 潘伟釉 王科宇 贾湘 冯晶 王凤涛 郭青云 蔺瑞明
关键词小麦条锈病;持久抗性;抗性鉴定;遗传分析
小麦条锈病是由条形柄锈菌小麦专化型Puc-cinia strii formis f.sp.tritici(Pst)侵染引起的真菌病害,主要危害小麦叶部,也能侵染叶鞘、麦芒和颖片。它是典型的跨大区流行性气传病害,病原菌群体的毒性变异频率高,可随高空气流远距离传播。全球各小麦栽培区域均有条锈病发生,但在我国条锈病危害尤为严重,严重威胁着我国粮食生产安全。由于条锈菌新优势小种不断产生,主栽品种抗病基因较单一,遗传背景较为相似,导致新审定的小麦品种大面积推广应用几年后便会“丧失”抗条锈性,不仅缩短了品种有效使用期限,还极易引起条锈病大流行。最新出现的条锈菌CYR34小种已克服了Yr10、Yr24和Yr26等已知抗病基因,是目前毒性谱最宽、毒性最强的小种,能侵染大部分绵麦系、绵农系、蜀麦系、川育系、川麦系、兰天系、天选系、中梁系、洮字系和小偃系等我国西北和西南麦区小麦主栽品种,可能在今后一段时间内仍将是主要优势小种之一。其他品种如洛类衍生系、Sull系、阿夫系、陇鉴系和繁6衍生系等大批品种也对CYR32和CYR33优势小种感病。选育并广泛使用持久抗病品种是防治小麦条锈病最经济有效的措施。目前多数已知抗条锈病基因已被病原菌克服,故需要不断发掘新的抗病基因并引入育种体系,是实现主栽品种中抗病基因多样化和持久抗病性的有效途径。作物品种的持久抗病性为现代育种指明了一个延缓抗性丧失的育种新方向,特别是对于持久稳定控制小麦条锈病尤为重要。
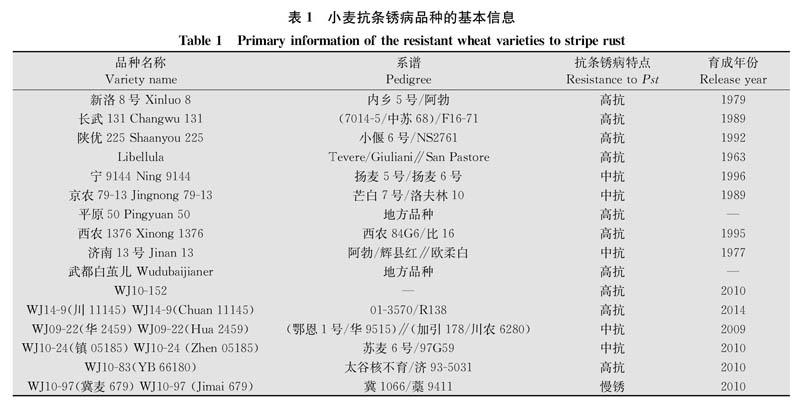
抗病基因的遗传特点及互作模式决定基因的功能和持久性。我国部分利用多年的小麦种质资源至今仍保持稳定的抗条锈性,是重要的抗源材料,但其对当前条锈菌流行小种CYR32的抗性遗传模式尚不清楚。本研究中分析具有稳定抗条锈性的10个代表j生品种(‘新洛8号‘长武131‘陕优225‘Libellula‘宁9144‘京农79-13‘平原50‘西农1376‘济南13号和‘武都白茧儿)和近年育成的6个新品系(‘WJ10-152‘wJl4-9‘WJ09-22‘WJ10-24‘WJ10-83和‘WJ10-97)的遗传组分和遗传模式,初步明确这些品种的基本遗传特点,为发掘抗病新基因提供材料,对进一步丰富我国小麦抗条锈病基因资源具有重要的理论意义和潜在的应用价值。
1材料与方法
1.1材料
具有持久稳定抗条锈病的小麦品种及新育成的抗病品系(表1)作为抗病基因供体父本,感病材料‘Taichung29和‘铭贤169作为感病受体母本,采用常规有性杂交方法配制组合,获得F1代杂交种,经自交和回交分别得到F2和BCI代种子。利用条锈菌小种CYR32的單孢菌系在成株期进行田间接种鉴定,遗传分析群体及亲本的抗病性。
1.2成株期抗病鉴定方法
将构建群体的亲本,F1、F2和BC1代材料播种于中国农业科学院植物保护研究所廊坊试验基地抗病鉴定圃,小区行长3m。亲本播种1行,F1和BC1播种2行,F2群体播种20行,每行播种30粒种子。小区四周种植感病品种‘铭贤169作为诱发行。采用小麦条锈菌小种CYR32的单孢菌系接种用于抗病性鉴定。于4月上旬田间喷雾接种诱发行‘铭贤169,接种前对待接种的诱发行浇水。在三角瓶中配制条锈菌夏孢子悬浮液母液(含0.025%吐温20),稀释后在气温较低的傍晚(低于18℃)喷雾接种诱发行,接种后用塑料薄膜覆盖保湿过夜。接种后应及时浇水,保证地面潮湿,促进条锈菌再侵染传播扩散。在5月下旬,当感病对照‘铭贤169发病充分,上部叶片病害严重度达80%以上时进行抗病性调查,间隔1周后进行第2次调查。成株期侵染型调查采用“0~4”级共划分为6级的基本标准,即0、0;、1、2、3和4,并用“+”和“-”进一步详细划分为12级,调查倒数第2叶、倒数第3叶和旗叶的侵染型及严重度。依据双亲,F1、F2及BC1代植株侵染型级别及各级侵染型的株数划分抗感类型,计算卡方(42)值,根据F1代植株抗病表型以及BCi代植株和F2代群体抗感分离情况进行遗传统计分析。
2结果与分析
2.1抗性持久的种质资源抗条锈病遗传分析
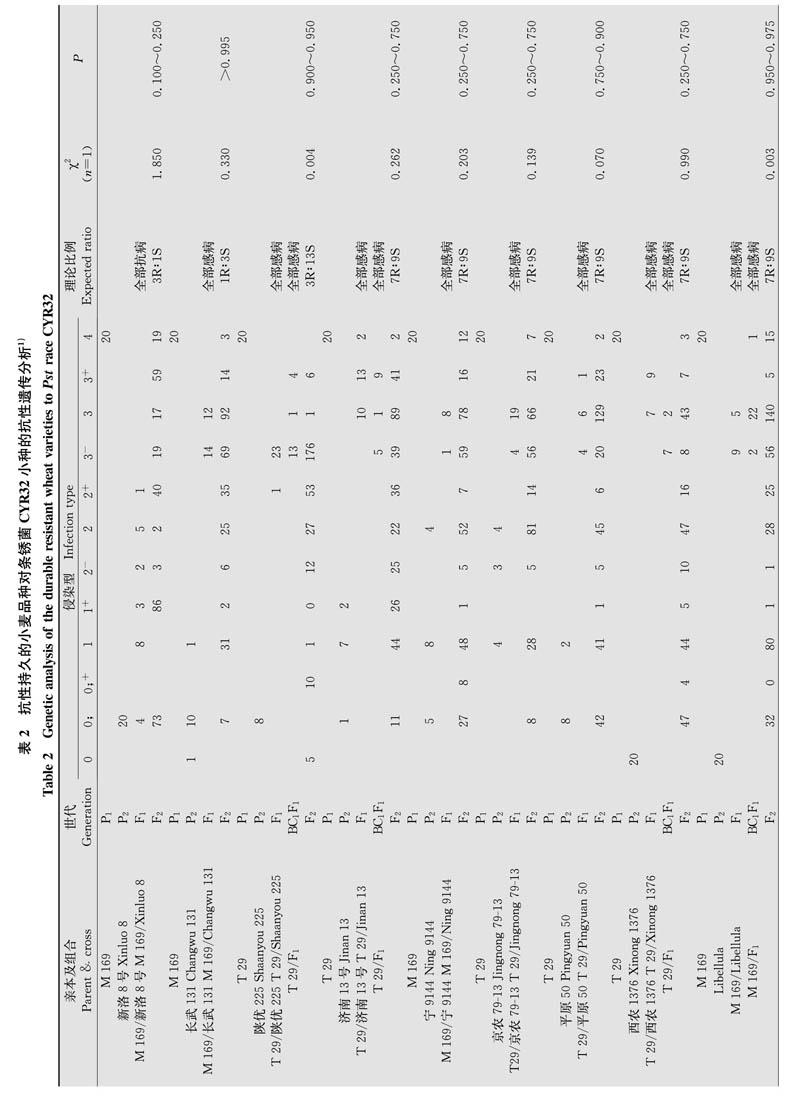
对一些已经验证的具有持久稳定抗条锈病特点的品种进行遗传分析结果显示,‘济南13号‘京农79-13‘宁9144‘平原50‘西农1367和‘Libellu-la对CYR32小种的抗病性均由2对隐性基因控制,‘长武131由1对隐性基因控制,‘陕优225由1对显性基因和1对隐性基因互补作用控制,‘新洛8号和‘武都白茧儿对条锈病抗性分别由1对和2对显性基因控制。10个抗性老品种中,8个品种对CYR32小种的抗性受隐性遗传基因控制,同时有8个品种对CYR32小种的抗性受2对基因控制。因此,多数抗性持久稳定的品种对CYR32小种抗病性由1对或2对隐性遗传抗病基因控制(表2,表4)。
2.2新育成小麦品系的抗条锈病遗传分析
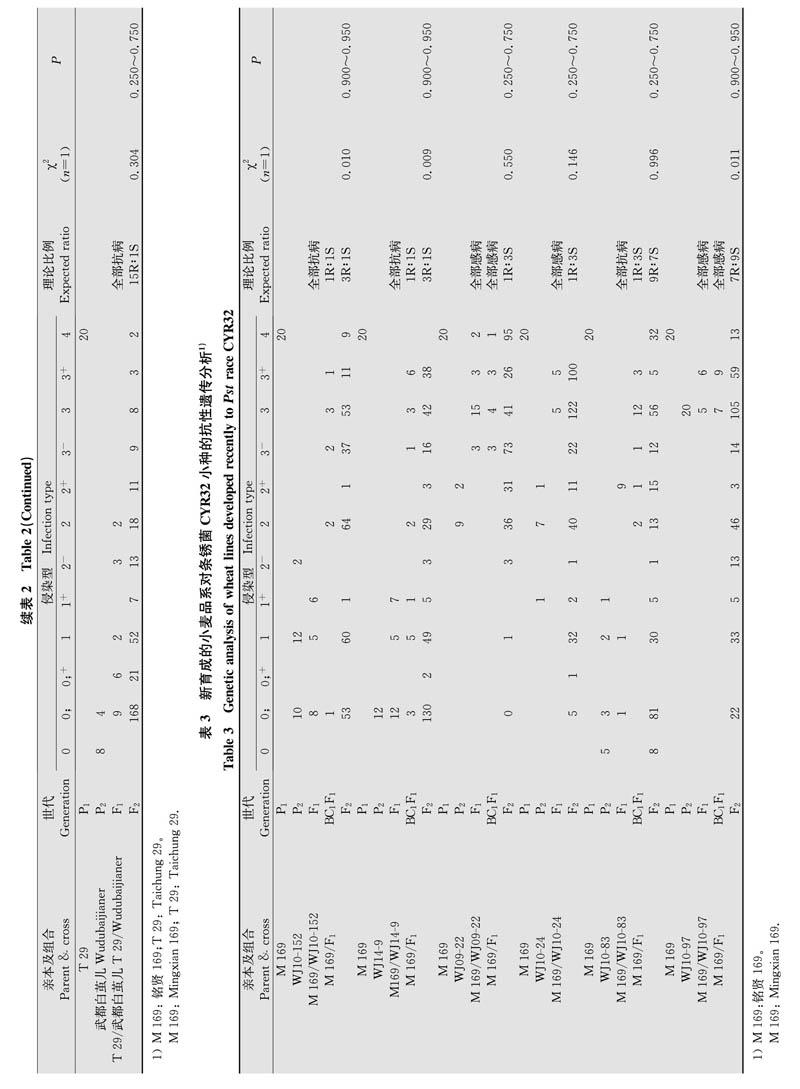
对新育成的6个小麦品系遗传分析结果显示,‘WJl0-152和‘WJl4-9对供试菌系的抗条锈性均由1对主效显性基因控制,‘WJ09-22和‘WJl0-24抗条锈性受1对主效隐性基因控制,‘WJ10-83抗条锈性由2对显性基因互补作用控制,‘WJ10-97的抗条锈性由2对隐性基因重叠或独立作用控制。因此,3个新育成品系对CYR32小种的抗病性受隐性遗传基因控制,另外3个新育成品系对CYR32小种的抗性受显性遗传基因控制;4个新育成品系的抗病性由1对主效基因控制,其他2个品系由2对基因调控(表3,表4)。因此,供试新育成品种对CYR32生理小种的抗性主要受单基因控制,抗病基因显性或隐性遗传特点的偏向性不明显。
3讨论
根据病害发生和流行特点以及流行区域的生态气候条件差异,可将我国小麦条锈病流行区划分为8个主要区域,其中甘肃天水、陇南以及四川西北部是条锈菌关键的越夏区和主要的秋季菌源地,也是病原菌的冬繁区域。病原菌群体在局部生态区连续侵染和大量扩繁非常有利于突变个体的存活。另外,流行区自然环境条件的多样性极大增加了条锈菌群体毒性变异频率。目前生产上主栽品种中抗病基因过度单一化增加了对病原菌群体毒性变异定向选择的压力,从而加速新致病类型的产生及其大规模流行,引发条锈病大规模流行,造成了严重的经济损失。因此,发掘具有抗病谱宽而且抗性持久稳定的新基因位点、培育抗性持久稳定的新品种是有效防控条锈病首选措施。另外,抗性持久稳定的小麦基因型有利于延缓条锈菌毒性变异进程,减少经济损失。因此,分析抗源材料的遗传组成及其遗传特点可对培育抗性持久稳定的新品种提供重要的参考信息。
本研究中对具有抗性稳定持久的小麦品种和近年来选育的抗条锈病新品系进行遗传组分和遗传特点分析,并比较这两类材料的抗条锈病遗传特点差异。例如10个抗性稳定持久的小麦老品种中,多数品种对CYR32小种的抗性是由隐性遗传基因控制,而且其中8个品种抗条锈病至少由2对主效基因控制(表2,表4);4个新育成品系的抗病性由1对主效基因控制,其他2个品系由2对基因调控(表3,表4);而新育成品种对CYR32生理小种的抗病性主要受单个主效基因控制,显性或隐性遗传的偏向性不突出(表3,表4)。对条锈病抗性持久稳定的小麦基因型的遗传分析表明,这类材料具有主效抗病基因多为隐性遗传而且含有2对或多对主效抗病基因的特点,与我国抗条锈病地方品种的遗传特点相似,如‘小红芒对CYR32小种的成株抗条锈性是由1对隐性基因控制,‘红疙瘩和‘疙瘩糙对CYR32小种的抗性分别由3对隐性互补基因和1对隐性基因控制。农家品种具有丰富的抗条锈等位基因,其群体遗传多样性水平较高。目前多数抗性持久稳定的品种是从农家品种经多代系选或杂交改良后再经系选方式获得,其抗条锈基因遗传组成及遗传特点对农家品种具有一定的继承性。
mlo是隐性遗传的持久抗病基因的范例,已在欧洲及世界其他大麦种植区域广泛应用多年,特别是在欧洲利用该基因已基本控制了白粉病的发生和流行。这些自然突变的隐性遗传抗病基因可能是丧失功能的感病因子的等位位点,目前正在从麦族及其他作物中鉴定出这类隐性遗传基因。大麦广谱抗秆锈病基因rpg以及水稻抗稻瘟菌多个生理小种的抗病基因pi21等隐性遗传基因都具有抗性持久稳定的特点。相对于隐性遗传抗病基因,显性遗传的抗病基因更易被克服从而丧失抗性。
此外,在对条锈菌CYR32小种成株期抗病性鉴定中发现,‘WJlo-97侵染型为3型,严重度为5%,具有高侵染型、低严重度和产孢量少的典型慢条锈性抗性特点。慢条锈性作为小麦成株期一種特殊的抗病类型,对病原菌毒性变异的选择压力较小,也可以降低病害所造成的经济损失。因此,慢条锈品种‘WJlO-97可作为抗性品种选育的重要种质资源。同时应该注意的是,小麦慢条锈品种的抗病性受环境影响较大,其抗病性的研究可能因鉴定地区、鉴定年份的不同而有所差别。所以,对慢条锈品种的研究应以多地、多年的大量数据为基础。此外,在病原菌的自然选择压力条件下,具有持久抗性特点的地方品种及少数人工选育的品种中抗病基因隐性遗传方式与抗条锈病持久稳定性之间的关联性,还有待进一步深入研究。
