基于线粒体Cyt b基因和CR序列的养殖貉(Nyctereutes procyonoides)种群遗传多样性和结构分析
2021-04-28魏宏达刘志平姜春明史迎秋徐艳春
魏宏达 李 波,2,3* 魏 来 刘志平 姜春明 史迎秋 徐艳春,2,3*
(1.东北林业大学野生动物与自然保护地学院,哈尔滨,150040;2.国家林业和草原局野生动植物检测中心,哈尔滨,150040;3.国家林业和草原局野生动物保护与利用工程技术研究中心,哈尔滨,150040;4.哈尔滨华隆蓝狐育种有限公司,哈尔滨,150040)
貉(Nyctereutesprocyonoides)隶属于食肉目(Carnivora),犬科(Canidae),犬亚科(Caninae),貉属,原产于东亚地区;20世纪初期作为经济动物引入欧洲[1-2]。貉分为6个亚种:俄罗斯和中国东北的乌苏里亚种(N.p.ussuriensis)、朝鲜半岛亚种(N.p.koreensis)、中国南部和越南北部的指名亚种(N.p.procyonoides)、云南的滇北亚种(N.p.orestes)、北海道亚种(N.p.albus)、日本亚种(N.p.viverrin-us)[3-5]。我国20世纪70年代开始引入野生貉种群(乌苏里亚种为主)开展规模化驯养繁殖[6]。1990年由野生型貉培育出白色型貉,命名为吉林白貉[7];2006年山东临沂饲养种群中出现了红褐色型貉[8]。这丰富了养殖貉种群的色型,提升了其经济价值,使其成为我国重要经济动物之一。但经过50年的人工饲养和繁育、无新野生个体引入、遗传漂变等因素可能导致貉种群遗传多样性水平下降,近交衰退的几率增加。为此有必要彻底调查养殖貉种群遗传多样性水平和遗传结构,溯源其母系起源,这有助于制定科学的遗传管理方案,有利于养殖种群的可持续发展。
国内对于貉种群遗传学研究包括中国貉种群的亚种分类:随机扩增多态DNA分析结果表明中国貉分为4组:广西、安徽、陕西、云南—越南[9];线粒体DNA(mtDNA)限制性内切酶分析结果说明分布于华北和陕西的貉种群应各自独立为亚种[10]。研究还显示貉毛色变化与多个基因的多态性、突变和表达相关:黑素皮质素受体1(MC1R)、鼠灰色基因(Agouti)、白色基因(KIT)[11-13]。国际上研究了野生貉种群遗传多样性和遗传结构:线粒体控制区(CR)分析表明俄罗斯貉种群可以划分为2个mtDNA世系[14];16个微卫星分析结果说明欧亚大陆种群和日本岛屿种群间有显著的遗传分化,大陆种群可分为朝鲜半岛南部、中国—俄罗斯、越南3个亚群,且其种群遗传多样性水平依次递减[15]。基于细胞色素b(Cytb)基因的貉系统地理学研究说明该物种存在多个冰期避难所,韩国和日本貉种群间没有发生基因交流[16]。总之,由于采集样本难以全面覆盖各地理种群,国内貉种群的种下分类及与国外邻近地区种群的关系还需进一步分析。本研究利用非损伤性技术收集了东北和华北地区养殖种貉的毛样,测定了其线粒体Cytb基因和CR部分序列;结合GenBank中已知的同源序列综合分析了国内貉养殖种群的遗传多样性和遗传结构,揭示了其母系起源。
1 材料和方法
1.1 材料
利用非损伤性技术收集了黑龙江养殖场野生型貉102只、红褐色型貉13只、白色型貉5只和河北养殖场野生型貉40只的毛样本。采集毛样品多数有毛囊,放入取样袋中密封、-20℃保存备用。
1.2 PCR扩增与测序
试验材料经预处理后利用蛋白酶K消化,后用基因组提取试剂盒(上海百赛生物工程技术有限公司)提取和纯化总DNA。利用引物L14724/H15915[16]和NycDLF0/NycDLR01[14]分别扩增貉线粒体Cytb基因和CR部分序列。PCR反应在50 μL体系中进行,包括2×EasyTaqSuperMix(Transgen,北京)25 μL,正、反向引物(10 pmol/μL)各1 μL,DNA模板5 μL,灭菌去离子水 18 μL。扩增采用PE9700型和2400型DNA扩增仪(PE,美国),PCR扩增程序:94℃预变性3 min,94℃变性45 s,44℃/58.5℃(Cytb/CR)退火45 s,72℃延伸90 s/60 s(Cytb/CR),扩增35个循环,最后72℃延伸10 min/5 min(Cytb/CR)。PCR扩增产物用1.5%琼脂糖凝胶电泳分离(220 v/25 min)。电泳后将PCR产物送至哈尔滨擎科生物科技有限公司,利用原扩增引物进行双向测序。
1.3 数据分析
从GenBank下载66条同源序列:乌苏里亚种CR序列30条(JF809819-JF809848),乌苏里亚种Cytb基因5条(JX099862-JX099866),朝鲜半岛亚种Cytb基因8条(JX099854-JX099861),日本亚种Cytb基因17条(JX099869-JX099870、JX099872-JX099880、JX09 9884-JX099889),北海道亚种Cytb基因4条(JX09 9871、JX099881-JX099883),指名亚种Cytb基因2条(JX099867、JX099868)。将实验所得Cytb基因和CR序列峰图导入DNAstar软件包的Seqman程序进行拼接,对内部位点进行人工校对,并与下载的貉同源序列进行对比确定变异位点。利用MEGA 5.2.1[17]软件计算Cytb基因和CR序列的碱基组成。利用DNAsp 6[18]软件分析确定单倍型、计算单倍型数(haplotype number,H)、单倍型多样性(haplotype diversity,Hd)、核苷酸多样性(nucleotide diversity,Pi)以及平均核苷酸差异(average number of nucleotide differences,K)、多态位点数(number of polymorphic sites,S)等数据。
选用最大似然法(maximum likelihood,ML)和贝叶斯推论法(Bayesian inference,BI)进行系统发育分析。在MEGA 5.2.软件中计算核苷酸最佳替代模型,并根据贝叶斯信息准则(Bayesian information criterion,BIC)选择最佳模型——Hasegawa-Kishino-Yano+G(HKY+G)[19]。利用IQ-TREE构建ML树[20],自举检验值(bootstrap,BS)由1 000次重复检验获得;使用软件MrBayes 3.2.1[21]构建贝叶斯树,主要参数包括:将马尔可夫链运行2 000 000代,每100代记录1次树,舍弃最先生成的25%树,由后验概率(posterior probabilities,PP)评价其置信度。2种树都以北极狐线粒体基因组中的同源序列作为外群(序列号NC_026529)。
2 结果
2.1 序列特征和单倍型
实验测得158条Cytb基因序列(1 140 bp)。平均碱基组成:29.58% A、13.75% G、28.64% C、28.03% T,A+T含量(57.61%)高于C+G含量(42.39%)。在1 140 bp中包含15个变异位点(1.36%),其中简约信息位点13个,单突变位点2个。实验测得145条CR序列(659 bp)。平均碱基组成:26.93% A、17.29% G、27.39% C、28.39% T,A+T含量(55.32%)高于C+G含量(44.68%)。在659 bp中包含19个变异位点(2.88%),其中简约信息位点17个,单突变位点2个。
158条Cytb基因序列定义了5个单倍型,其中有3个与俄罗斯产乌苏里亚种共享单倍型(CH1、CH2、CH3),2个新单倍型(CH4、CH5)。养殖种群优势单倍型为CH1(48.73%)。145条CR序列定义了6个单倍型,其中有4个与俄罗斯产乌苏里亚种共享单倍型(DH1、DH2、DH3、DH4),2个新单倍型(DH5、DH6)。种群优势单倍型为DH1(67.59%),单倍型CH5-DH5来自黑龙江养殖场。
2.2 种群遗传多样性
貉亚种种群及我国养殖种群Cytb基因核苷酸多样性(Pi)及平均核苷酸差异(K)等信息见表1。养殖种群的单倍型多样性为(0.637±0.020),核苷酸多样性为(0.523±0.009)%,平均核苷酸差异为5.96。国内养殖貉种群Cytb基因核苷酸多样性和平均核苷酸差异均略低于产于俄罗斯的乌苏里亚种野生种群(0.596%和6.800),高于其他4个亚种的野生种群。
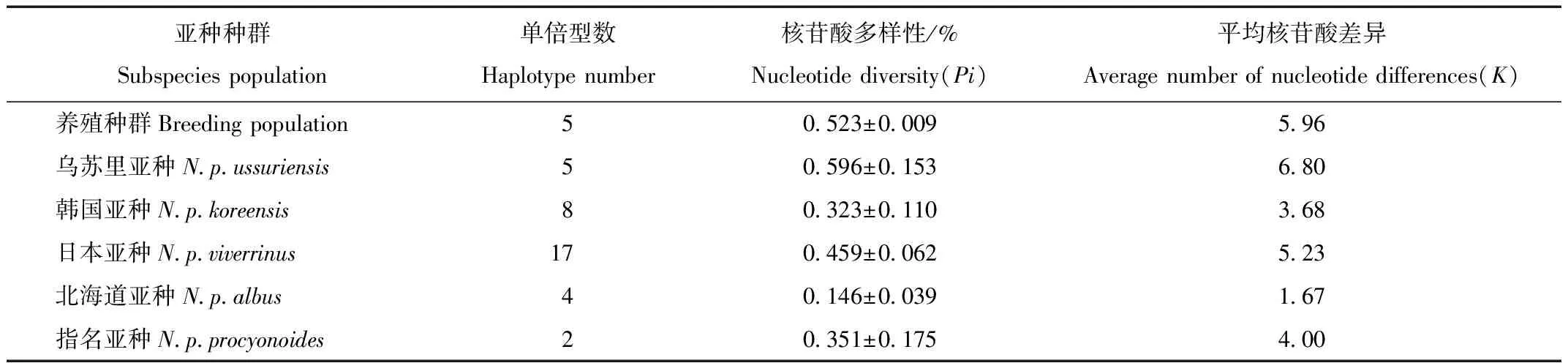
表1 貉Cyt b基因的遗传多样性参数
俄罗斯乌苏里貉野生种群及我国养殖种群CR单倍型多样性(Hd)、核苷酸多样性(Pi)及平均核苷酸差异(K)等信息见表2。国内养殖种群单倍型多样性为(0.500±0.042),核苷酸多样性为(0.982±0.071)%,平均核苷酸差异为6.46。我国东北与华北地区的貉养殖种群遗传多样性水平一致,但都明显低于同亚种的俄罗斯野生种群。
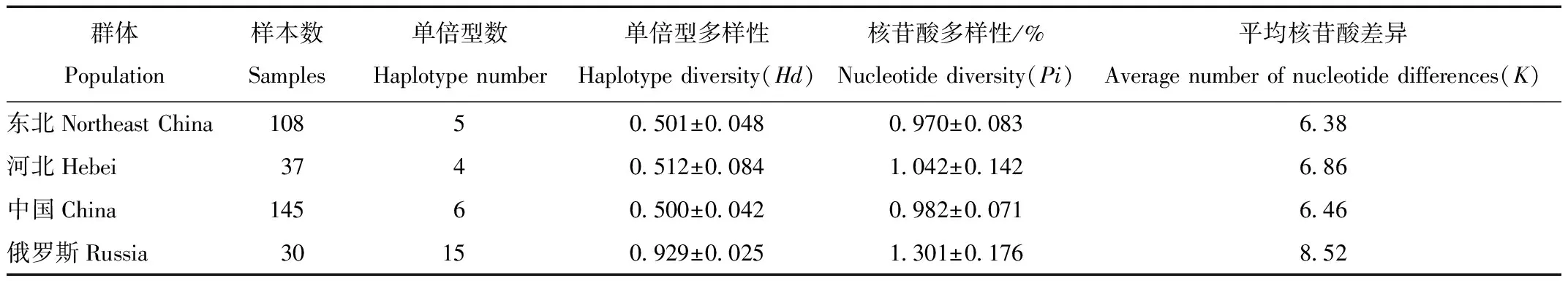
表2 貉CR序列的遗传多样性参数
国内养殖的野生型、白色型和红褐色型貉种群单倍型多样性(Hd)、核苷酸多样性(Pi)及平均核苷酸差异(K)等信息见表3。Cytb基因和CR序列2种分子标记分析结果都表明白色型遗传多样性水平略高于野生型,这应与白色型样本量偏小有关。红褐色型貉单倍型多样性和核苷酸多样性都显著低于野生型貉,涉及的Cytb基因和CR序列单倍型各自均只有2个,优势单倍型为HC2-DH2(92.30%)。这说明红褐色型貉主要是单一单倍型的毛色基因突变而来。

表3 野生型、白色型和红褐色型貉种群遗传多样性参数
2.3 种群遗传结构
基于Cytb基因和CR序列单倍型构建最大似然树(ML)及贝叶斯树(BI),除置信度不同,2种树的拓扑结构大致相同(图1,图2)。基于Cytb基因的系统发育树说明东北亚地区分布的野生貉种群可分化为4个mtDNA世系:越南的指名亚种为世系1;日本亚种与北海道亚种独自组成世系2,与大陆种群(乌苏里亚种、朝鲜半岛亚种、指名亚种)差异明显;7个朝鲜半岛亚种单倍型与4个乌苏里亚种单倍型组成世系3;1个朝鲜半岛亚种单倍型与2个乌苏里亚种单倍型组成世系4。这说明朝鲜半岛亚种与乌苏里亚种有紧密的系统发育关系。国内养殖貉种群有4个Cytb基因单倍型均匀分布与3、4世系,与俄罗斯产乌苏里貉有紧密的系统发育关系,还有1个单倍型(CH5)分布于1世系,与越南产指名亚种有更近的亲缘关系。基于CR序列的系统发育树表明俄罗斯乌苏里貉野生种群和国内养殖种群可分为A、B、C 3个mtDNA世系,B与C世系有更近的亲缘关系。A世系(BS=67%,PP=51)包含2个共享优势单倍型(DH1和DH4,频率62.87%),B世系(BS=37%,PP=71)仅有单倍型DH5,C世系(BS=96%,PP=82)含有2个共享单倍型(DH2和DH3)和1个新单倍型(DH6)。由于单倍型CH5与DH5来自同一个体,这说明Cytb基因和CR序列得出的国内养殖貉种群遗传结构已一致。同时,也表明国内养殖的貉种群多数属于乌苏里亚种,极少数个体属于指名亚种。
3 讨论
遗传变异的程度反映物种进化潜力的大小,决定了物种的生存能力[22-23]。本研究利用线粒体Cytb基因和CR序列评估国内养殖貉种群遗传多样性水平。结果显示,尽管养殖貉种群Cytb基因遗传多样性水平与俄罗斯同亚种的相近,但CR区上显著低于后者。这与2种分子标记的进化速率不同有关:Cytb基因进化速率适中,适合研究种内、种间、科间的系统发育关系[24];CR区因选择压力小而进化速率快,适合研究亲缘关系较近的群体[25]。因而,CR区的分析结果更能体现养殖貉种群遗传多样性的真实水平。Grant等[26]利用单倍型多样性(Hd)和核苷酸多样性(Pi)评估种群遗传多样性水平高低的临界值分别是0.5和0.5%。该标准也被用于研究哺乳动物种群遗传多样性,例如鹅喉羚(Gazellasubgutturosa)[24]、淮南猪(Susscrofa)[27]。根据该标准养殖貉种群单倍型多样性处于临界点、核苷酸多样性处于较高水平。这种遗传多样性模式可能主要是由奠基者效应导致的。养殖貉种群建群时个体数偏少,缺乏遗传管理的繁殖模式促使原来有限的单倍型丢失的风险增加,但遗传漂变造成核苷酸多样性丢失要缓慢得多。同时,以生产性能作为单一选育指标的繁殖模式作用下,种群内会不可避免地发生近交,从而造成遗传多样性的丢失和个体适合度下降[28-29]。
红褐色型貉遗传多样性水平显著低于其他2种色型,涉及的优势单倍型为CH2-DH2。这与该色型产生模式——由毛色基因突变获得新色型,依赖近亲繁殖扩繁种群数量[8,30]是一致的。白色型貉种群遗传多样性水平略高于野生型貉,可能是因为取样存在偏差。此外,吉林白貉的色型为杂合显性,近亲交配会出现基因纯合致死现象[31]也是不容忽视的因素。因此,为维持养殖貉种群的质量,建议在保证生产性能的基础上将遗传多样性变化引入种群遗传管理中。当种群遗传多样性和近交衰退达到临界值时考虑引入具有新基因型的野生貉改良现有种群。
单倍型分析显示,国内养殖貉种群与俄罗斯野生种群间存在3个Cytb基因和4个CR序列优势共享单倍型,这说明二者同为乌苏里亚种[32]、种群间有紧密的系统发育关系。野生貉种群易受环境因素限制,活动范围较小[33]。本研究还发现养殖种群的Cytb基因单倍型CH5与越南产的貉指名亚种单倍型亲缘关系较近。这表明早期建群引种过程中可能偶然引入了华南地区分布的指名亚种。基于Cytb基因和CR序列系统发育关系分析得出了相同的结果,即养殖貉种群来自3个mtDNA世系。部分世系划分的置信度偏低,这与选取的线粒体序列片段较小有关,增加片段长度可能会提高置信度[34]。相较于俄罗斯的乌苏里貉,国内养殖貉种群建群时个体的涵盖度较广,不同世系个体的交配繁殖,会有效抑制近交衰退的效应。
综上分析,我国养殖貉种群遗传基础较好,但明显低于俄罗斯野生乌苏里貉种群,且已经出现遗传多样性下降的迹象。需要监控种群遗传多样性的变化,引入具有新基因型的野生貉种,制定科学的交配模式,避免近交衰退。
猜你喜欢
——乌苏里白鲑
——和田盘羊
