青藏高原东南缘陇蜀杜鹃当年生小枝茎叶性状对海拔变化的响应
2021-04-25何家莉王金牛周天阳宋怡珂NiyatiNaudiyal
何家莉, 王金牛, 周天阳, 宋怡珂, 查 琳, Niyati Naudiyal, 石 凝, 吴 彦
1 四川师范大学生命科学学院, 成都 610101 2 中国科学院成都生物研究所, 成都 610041 3 中国科学院武汉植物园, 武汉 430074
植物为了适应外界环境通常表现出一定的表型性状,即表型可塑性是植物克服环境压力的重要途径之一,具有重要的适应及生态进化意义[1-2]。枝条作为植物体运输水和有机物的重要通道,且支撑着其上叶、花及果实生长发育。通过调整枝条结构能改变叶、花和果实的空间排布方式,进而提高光的捕获效率、传粉授粉的成效和种子的扩散等[3- 7],反映了物种对资源的利用和适应策略[8- 10]。
植物整个生命周期均离不开光照、水分等环境因子的影响[3,11- 12],不同环境下植物体枝茎结构的调整反映了对可利用资源如光、热等环境因子的响应以及相邻个体间竞争状态[7, 13- 15]。植物枝条的结构可以用枝条的茎长、茎粗和茎纤细率表示,茎长和茎纤细率直接影响其上叶片的空间排布以及资源利用获取的效率,茎粗则影响机械支撑强度与水分和养分在器官间的分配效率。海拔作为重要的地形因子,使得温度、降水和光照强度等环境因子在小范围内呈梯度性变化[16-20]。海拔作为综合环境因子,上升到一定高度后所导致的低温以及大风雪等胁迫都可能影响植物体抗逆性和适应性,功能小枝的形态及资源分配相应的调整有助于减轻环境压力且充分利用养分资源。高海拔的植物倾向于增加小枝茎的投资,如较粗的茎能够支撑较重的繁殖与光合构件,同时也能抵抗较大的风力,而较长的茎有利于植物体捕获光能、花的授粉效率、种子的扩散等[3, 5-6]。但是,高寒地带的大风强雪等外界环境对植物体作用的机械拉力限制了小枝不能随意过度延伸,所以小枝茎的结构在纵向与径向上具有一定的权衡特征。近年来国内外有关小枝功能性状研究主要集中在小枝-叶大小关系上,包括茎构型或茎生物量与叶大小关系在不同物种或森林群落间的差异[21- 27]或单一物种枝叶关系在生境梯度上的变化[2, 22,28]。陇蜀杜鹃(Rhododendronprzewalskii)广泛分布于青藏高原东南缘高山林草交错带,研究其位于不同山体的当年生小枝茎叶性状和茎性状间相关关系,通过综合分析小枝功能性状沿着海拔变化的空间异质性响应,对于理解高寒环境梯度下灌丛植物的适应性特征以及生态学内涵非常关键。
以陇蜀杜鹃为研究对象,通过分析三处不同山体沿着海拔梯度陇蜀杜鹃灌丛小枝茎叶性状的变化,基于小枝形态和生物量的相互关系变化以及小枝茎叶性状与土壤因子的关系,拟阐明如下三个问题:(1)小枝茎、叶性状在不同山体海拔梯度下的空间异质性变化;(2)小枝茎功能指标之间的相关性;(3)个体水平的不同海拔小枝茎叶性状与土壤因子的关系。从而探究高寒灌丛陇蜀杜鹃个体尺度上小枝茎叶性状沿着海拔综合环境梯度变化的空间可塑性及其生态适应性意义。
1 材料和方法
1.1 研究区域概况
试验地位于岷江源区,该区地处青藏高原的东南部边缘地带,以卡卡山和祁让沟为南界,斗鸡台为北界,地理位置为103°39′—103°43′E,33°02′—33°28′N之间。岷江源区地势高亢,地貌类型多样,海拔基本在3400 m以上,受东南季风控制,属于山地季风气候。年均气温5—10℃,最低气温为-10—-20℃,最高气温30℃。年降水量为700 mm左右,全年降雨不均,雨季降雨量占全年降雨量的70%左右,积雪期长达4—6个月[29- 31]。
岷江源区为浩瀚的原始森林,植被类型多样,组成成分复杂,植被垂直带结构分明[29]。陇蜀杜鹃作为岷江源区主要的高山伴生灌丛之一,主要分布在海拔3600 m以上的开阔草甸区域,在卡卡山的林草交错带及针阔叶混交林下也有分布(海拔3600 m以上)。
1.2 试验设计
在岷江源区选择斗鸡台、弓杠岭、卡卡山三座山作为取样地,在陇蜀杜鹃的自然分布地取样,在斗鸡台3834 m和3779 m、弓杠岭4040 m和3963 m、卡卡山3921 m和3836 m处取样,具体样地信息见表1。在每个海拔设置一条宽度为30 m的水平样带,设置3个10 m×10 m的样方,随机选取样方内长势良好的3个杜鹃灌丛,每个灌丛取叶片完好的当年生小枝两枝,装入自封袋后带回实验室。这里的当年生小枝为枝条最末端无分支的当年生枝条,包括其上的叶、花和果实[17]。在每个海拔取样的灌丛附近使用环刀(规格100 cm3)对地表10 cm以下处各取3个土样,将采集的土壤放入自封袋,带回实验室进行测定。

表1 取样样地基本信息
1.3 测定方法
取样时,使用精度为0.01 m卷尺测定取样灌丛的高度以及冠幅。样品取回后使用精度为0.01 mm游标卡尺,测量新鲜小枝的茎长和茎粗。使用平板扫描仪(CanoScan5600F,Canon,Tokyo,Japan)扫描新鲜叶后用Photoshop CC计算叶面积。随后在75℃条件下烘干小枝及叶片至恒重,使用精度为0.001 g的电子天平称量小枝的茎生物量与叶生物量。获得的指标包括:茎性状(茎长(Stem length,SL);茎粗(Stem diameter,SD);比茎长(Specific stem length,SSL);茎生物量(Stem Biomass,SB))、叶性状(总叶质量(Total leaf mass,TLM);比叶面积(Specific leaf area, SLA))和灌丛大小(高度(Shrub height,SH);冠幅(Shrub width,SW))。其中比茎长(SSL)=茎长(SL)/茎生物量(SB)。
土壤指标测定:利用烘干称重法测定土壤含水量(soil variable water content,VWC)及土壤容重(Soil bulk density,SBD)。土壤有机碳(Soil organic carbon,SOC)含量的测定采用重铬酸钾外加热法,全碳(Total carbon,TC)和全氮(Total nitrogen,TN)含量采用元素分析仪(vario EL III,Elementar,Langenselbold,Germany)测定。全磷(Total phosphorus,TP)含量采用HNO3-H2O2微波消解—钼锑抗比色法测定,土壤pH利用METTLER TOLEDO pH计测定(水土比为2.5∶1)。
2 数据分析
对所取得的小枝数据使用Levene′s test检验方差齐性,对不符合正态分布的数据进行对数转换使其满足正态分布,使用单因素方差分析比较海拔及样地间小枝茎叶性状的差异,若方差不齐,使用非参数检验进行比较分析。根据相关生长模型y=bxa对三个样地不同海拔的小枝SL、SD和SB进行相关生长分析,方程可转化为log(y)=log(b)+a log(x),x和y代表特征参数,包括小枝SL、SD和SB,b为标准化常数,a为异速生长指数,a=1为等速生长关系,a≠1为异速生长关系[32],在双对数直角坐标系中,a表示斜率,log(b)为截距。相关生长模型的参数确定,采用标准化主轴估计(Standardized major axis estimation,SMA),利用R语言SMATR包完成[33-34]。
为了直观地反映陇蜀杜鹃小枝茎叶性状与土壤因子之间的相关关系,使用约束排序分析性状与环境因子之间的关系,采用典范对应分析(Canonical correspondence analysis, CCA)。用前选法(forward selection)和Monte Carlo转置法确定对茎叶性状有重要且独立作用的土壤因子变量组合,使用该组合用于CCA中并绘制排序图。植物性状与土壤环境的CCA排序图中,箭头表示环境因子,箭头越长表明对研究样本的影响越大,箭头连线与排序轴上的投影表明与排序轴相关性大小。环境变量间的夹角代表其相关性,夹角越小,相关性越大,研究对象到环境变量箭头的距离决定了其相关性大小,距离越大,关联性越小[35]。
数据分析在SPSS 19.0下进行,柱状图与散点图使用Origin 9.0绘制,CCA分析及出图均使用Canoco for Windows 5.0 软件完成。
3 结果与分析
3.1 不同山体土壤养分在海拔间的变化
研究结果表明,除弓杠岭外,卡卡山和斗鸡台海拔间土壤因子差异显著(表2)。具体表现为卡卡山位点高海拔C/N(10.28±0.21)显著小于低海拔(12.86±0.13)(P<0.05),土壤的TP(1.36±0.18)g/kg比低海拔TP(0.88±0.02)g/kg多0.48 g/kg,差异显著(P<0.05),其余养分指标则无显著性差异(P>0.05)。斗鸡台位点N%、TN(g/kg)、VWC%在海拔间无显著差异(P>0.05),高海拔土壤的C(7.08±0.60)%、SOC(3.87±0.36)%、TC(70.78±6.05)g/kg、TOC(38.76±3.64)g/kg、C/N(10.58±0.87)、pH值(5.01±0.09)均显著小于低海拔(16.96±1.59;8.73±0.37;169.57±15.97;87.31±3.70;16.93±0.77;6.41±0.29)(P<0.05),SBD(1.00±0.06)g/cm3及TP(1.01±0.01)g/kg则显著大于低海拔(0.54±0.07;0.59±0.05)(P<0.05)。相似海拔不同位点比较结果则表明,斗鸡台高海拔土壤的TP(1.01±0.01)g/kg显著大于卡卡山低海拔(0.88±0.02)(P<0.05),其土壤pH值(5.01±0.09)也显著大于卡卡山位点(4.43±0.04)(P<0.05)。卡卡山高海拔土壤pH值(4.37±0.06)与C/N(10.28±0.21)则显著小于弓杠岭低海拔(5.41±0.13;11.36±0.19)(P<0.05),SBD(0.99±0.02)显著大于弓杠岭位点的SBD(0.87±0.03)g/cm3(P<0.05)。

表2 不同山体土壤环境因子在海拔间的变化
3.2 不同山体植物功能小枝性状在海拔间的变化
斗鸡台、弓杠岭和卡卡山三处样地的灌丛小枝SL(mm)、SD(mm)、SB(g)和SSL(mm/g)、TLM(g)在不同海拔间均无显著性差异(P>0.05,P>0.05,P>0.05)(图1)。除卡卡山位点高海拔SLA(cm2/g)显著大于低海拔外,斗鸡台和弓杠岭的SLA在海拔间并无显著差异(P>0.05)。弓杠岭(3963 m)和卡卡山(3921 m)性状比较结果表明(图1),位于弓杠岭低海拔的灌丛小枝SL显著大于卡卡山高海拔(P<0.05),其余性状(SD、SB、SSL、SLA和TLM)均无显著差异(P>0.05)。卡卡山低海拔(3836 m)和斗鸡台高海拔(3834 m)的海拔位置相似但生境类型不同,卡卡山低海拔为林草交错带,斗鸡台高海拔为开阔草甸。以生境类型作单因素方差分析发现,林草交错带的灌丛小枝SL、SD、SB、SSL、SLA和TLM与开阔草甸的小枝性状并无显著差异(P>0.05)。

图1 陇蜀杜鹃当年生小枝功能性状在三处样地海拔间的比较。Fig.1 Comparison of the functional traits of the twigs of R. przewalskii in different elevations in three plots不同小写字母表示在同一样地不同海拔间差异显著(P<0.05);▽,▼分别表示不同样地的相似海拔,不同大写字母表示在相似海拔不同样地间差异显著(P<0.05)
3.3 陇蜀杜鹃功能小枝不同性状之间的相关关系
小枝性状间的相关性分析结果表明(表3),弓杠岭小枝SL、SD和SB两两之间均具有显著的正相关关系(低海拔SB-SLR2=0.384,P<0.01; SB-SDR2=0.281,P<0.01; SL-SDR2=0.172,P<0.05; 高海拔SB-SLR2=0.425,P<0.01; SB-SDR2=0.514,P<0.01; SL-SDR2=0.329,P<0.01),斗鸡台样点其小枝SB和SL有显著的正相关关系(R2=0.512,P<0.05;R2=0.580,P<0.01),卡卡山低海拔SB与SD有显著正相关关系(R2=0.381,P<0.01)。进一步对具有显著相关性状进行异速生长分析发现(表3),斗鸡台小枝SL与SB在高、低海拔均为异速生长关系,且SL增加速率慢于SB累积的速率,其SMA斜率分别为1.577(CI=1.060, 2.346)和1.847(CI=1.074, 3.178)。弓杠岭高海拔小枝SL与SB为异速生长关系,且SL增加速率小于SB累积速率,其SMA斜率为1.322(CI=1.010, 1.732),低海拔则为等速生长关系(P>0.05, 表3);SD和SB在高、低海拔均为异速生长关系,其SMA斜率分别为4.145(CI=3.270, 5.252)和3.928(CI=2.768, 5.574),SB增加速率大于SD增加速率;SL与SD在高、低海拔均为异速生长关系,且SL增加速率远大于SD增加速率,SL增加的速率约SD增加速率的2.8倍。卡卡山低海拔SB与SD为异速生长关系,其SMA斜率为5.404(CI=3.736, 7.816),表明SB累积速率比SD增加的速率快。

表3 陇蜀杜鹃功能小枝不同植物性状之间的异速生长分析
3.4 不同山体处植物性状与关键环境因子的CCA排序
由表4可知,不同山体CCA排序前两轴的累计解释拟合变异均达到98%以上,说明CCA前两轴已能够较好地反映植物性状与土壤环境因子之间的关系。
由图2可以看出,斗鸡台高海拔的植物性状受C%和C/N影响,C%对SB影响最大,其次为SD与SL,C%与C/N对SLA的作用大小差别不大。此外,SH也受到C/N的影响。低海拔性状则受C%、SOC%和TP影响。其中SH与C%关联性最大,其次是SOC%。SW、SB、SL、SD和SLA则受TP影响,TP对SL、SD影响最大,对SB、SW和SLA影响较小。

表4 植物性状与环境因子CCA排序
弓杠岭高海拔植物性状受SOC%、C/N和SBD影响,SBD对SL的影响作用最大,其次为SLA。SH则受到SOC%和C/N影响。C/N对SH的影响最大,其次为SW,对SB影响作用最小。低海拔植物性状则受C%与SOC%的影响,SLA受C%和SOC%影响最大,SB和SH则主要受SOC%影响,SL主要受C%影响(图2)。
卡卡山高海拔植物性状受C%、SBD和SOC%影响。其中SLA主要受SBD影响,SOC%其次。C%对性状的影响SH>SW>SD>SL>SB。低海拔植物性状受SOC%和C/N影响,其中SLA主要受SOC%影响,C/N其次。SOC%对SW的影响最大,C/N则影响SL和SD(图2)。
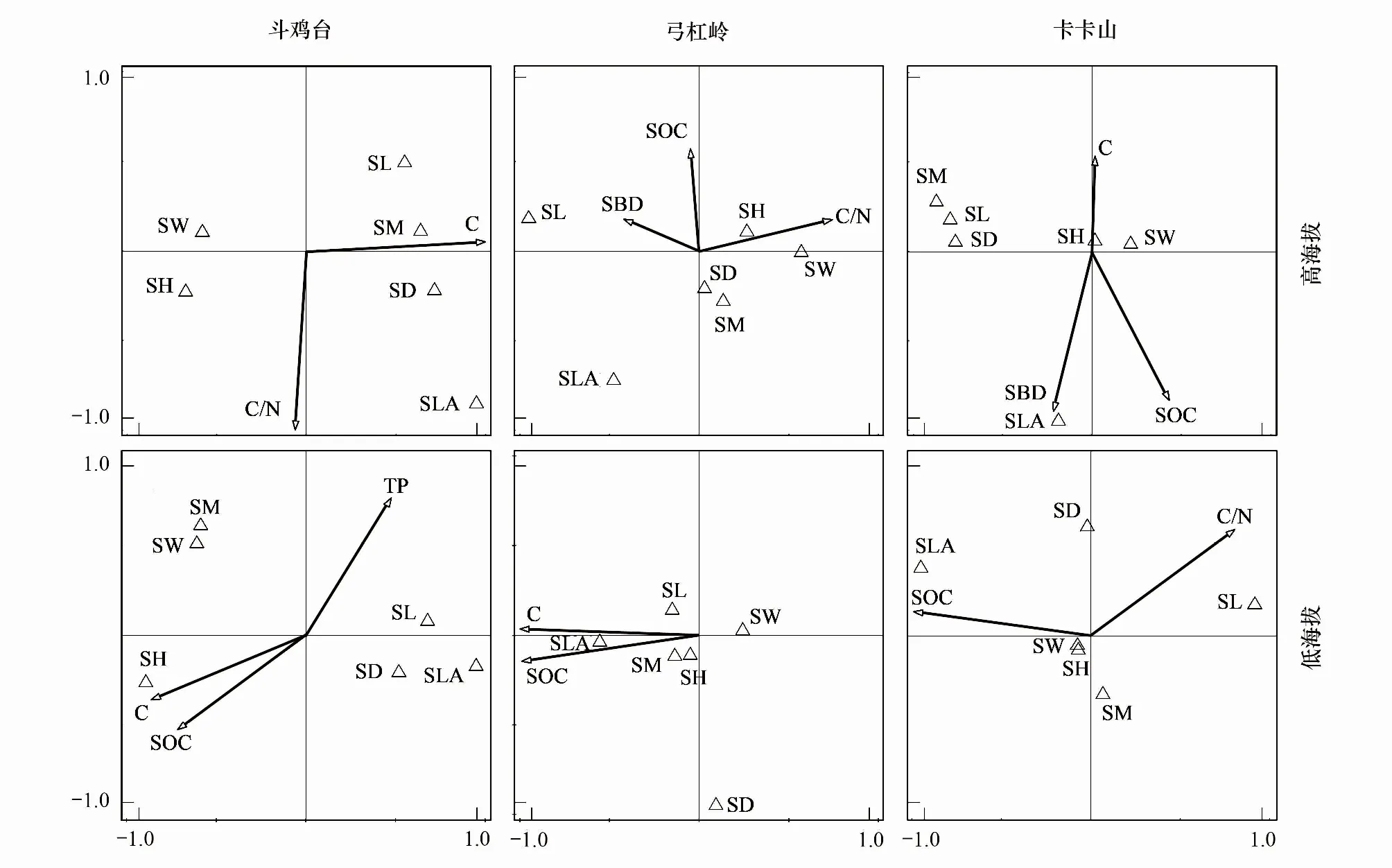
图2 三处陇蜀杜鹃植物性状与关键环境因子的CCA排序图Fig.2 CCA ordination diagram of plant traits and significant environmental factors at three sitesSH:灌丛高度 Shrub height;SW:灌丛大小 Shrub width
4 讨论
植物性状在面对环境胁迫时其表型具有高度的可塑性,对提高在胁迫环境中的适合度有重要作用[36-37]。山地木本植物其小枝性状在海拔梯度的变化已被广泛讨论,但结果存在一定的差异。本研究发现小枝茎的形态在海拔间无显著差异,且茎结构及其生物量并未表现出随海拔上升的变化,但Zhu等[38]在研究武夷山海拔梯度上三种森林群落的小枝与叶片关系变化时发现,中海拔混交林的小枝茎生物量小于其他海拔的常绿林与阔叶林。李曼等[39]在研究黄山松时发现,随着海拔上升,小枝茎长以及茎粗有上升的趋势,而卢宏典[40]关于黄山松小枝性状与海拔以及季节的关系研究时发现,尽管在生长末期茎长、茎粗以及茎纤细率在海拔间无显著差异,随着季节更替植物处于生长旺季时则表现出高海拔茎长、茎粗以及茎纤细率大于低海拔的现象。本研究与前人结果有异可能是与前人研究植物生活型及群落组成的不同,其次海拔跨度小于前人所研究的海拔跨度。此外,已有研究表明较长的小枝茎有助于增加光的获取以及提高繁殖成效[5-6]。本研究发现弓杠岭(3963 m)的小枝茎显著长于卡卡山(3921 m)的茎,CCA排序结果表明弓杠岭低海拔小枝长受土壤碳含量和有机碳含量影响,而卡卡山高海拔小枝长受土壤碳含量和容重影响。而两位点的土壤碳氮比、pH值和容重有显著差异,故土壤容重可能是造成小枝茎长差异的原因。从样地信息看(表1),弓杠岭取样地在山坡,较卡卡山取样的山顶受到的风、雪等外界环境压力较小,因此小枝茎增加受到的限制较小,也可能是两个位点的小枝茎长有显著差异的原因。叶性状仅卡卡山位点比叶面积在海拔间有显著差异,表现为低海拔大于高海拔。CCA排序表明卡卡山两个海拔的比叶面积均受土壤有机碳含量的影响,而卡卡山海拔间有机碳含量并无显著差异,说明土壤有机碳含量并不是造成比叶面积差异的原因。有研究表明[41]植物在高海拔较低海拔具有较小的比叶面积,高海拔的比叶面积较小可能是对有限资源的适应性表现,此外高海拔较大的风力容易给植物体造成损伤,叶片面积减小使得比叶面积较小。CCA排序结果表明茎叶性状受土壤因子影响,且海拔间及样地间受影响程度及因子有差异。
向双[42]关于亚热带阔叶林植物的研究得出茎生物量增长与茎长的异速生长指数约1.5,本研究结果与其大体一致,但茎生物量与茎粗的异速生长指数约为4.5,显著大于前人的研究结果值[40]。已有研究表明[43]较高的木密度有助于增加茎对病原体以及外界物理伤害的抵抗力,陇蜀杜鹃的茎生物量累积速率远大于径向生长速率可能是对于处在高寒生境下的一种保护机制。研究还发现茎结构与茎生物量在不同海拔以及样地表现出不完全一致的相关生长关系,即具有相对高的空间异质性。如在弓杠岭位点,随着海拔降低小枝茎长与茎生物量由异速生长关系转变为等速生长关系,茎粗与茎生物量成异速生长关系,且随海拔降低,小枝茎生物量累积速率减慢。卡卡山低海拔茎粗与茎生物量呈异速生长关系,至高海拔则变为不相关。可见茎结构与茎生物量相关关系较为复杂,在海拔以及不同的样地间相关关系差异较大,既与植物自身的物种属性和发育阶段有关,也受到所处的生境的微环境以及种间关系等外部干扰作用。植物纵向的生长与捕获光的效率以及种子的扩散有关,小枝纵向生长的同时需要进行径向生长以增强其机械支撑,以及增加水分及养分的运输[5],所以二者之间的权衡是研究植物生活史对策重要内容之一[43-44]。本研究发现陇蜀杜鹃当年生小枝纵向生长与径向生长也存在显著性异速生长的权衡关系,且小枝的纵向生长速率约为径向生长速率的2.8倍,远远大于陈国鹏[45]研究的沙柳丛生枝其基径与枝长的异速生长指数结果(值为0.70),这多半是因为选取的不同研究植物物种以及小枝与丛生枝的差别所导致。其研究中上端茎纵向生长的速率远大于径向生长的速率,这就与本研究涉及的当年生小枝非常类似。当年生小枝茎长的增加是上部竞争资源的需要,植物茎干在群落中所处的空间位置决定了其竞争能力和对资源的获取能力[46],而使得植物生长过程中,当年生小枝纵向生长速率较高。因此当年生小枝作为代谢活跃的构件在纵向生长的速率远远大于径向生长。但是,斗鸡台与卡卡山的小枝茎长与茎粗并未表现出明显的相关关系,这多半与植物发育阶段以及所处群落结构有关,已有研究表明植物的异速生长指数并非固定不变[47-49]。
5 结论
沿着海拔梯度所产生的土壤因子变化是高寒地带陇蜀杜鹃灌丛的小枝茎叶性状产生差异的原因之一。小枝茎生物量的累计速率大于茎长和茎粗的增加速率,提高了小枝的机械支撑作用,避免在伸长以及增粗的过程中受到外界的机械损伤。当年生小枝作为代谢活跃的构件,为了占取空间上的有利位置,其纵向生长的速率远大于径向生长。
