紫茎泽兰叶片提取物对4种杂草种子萌发及幼苗生长的抑制效应
2021-04-19张丽娜杨文秀马金虎杨小环
李 颖 张丽娜 杨文秀 马金虎 杨小环
(1山西农业大学农学院,山西 太谷 030801;2烟台爱华双语学校,山东 烟台 265500;3 云南农业大学热带作物学院,云南 普洱 665000;4山西农业大学工学院,山西 太谷 030801)
农田杂草防除是农业生产中面临的主要问题之一。我国常年受杂草危害的农作物面积达0.73 亿hm2。因杂草危害,直接经济损失近千亿元[1]。目前,农田除草主要依靠喷洒化学农药,但化学除草剂对生态环境造成污染及对农作物的药害问题已引起广泛关注[2-5]。植物源农药具有毒性低、易降解、有害生物不易产生抗药性等特点[6-7]。因此,挖掘植物天然活性产物,研发新型植物源除草剂,对于农田杂草防控具有重大的研究意义[8]。
紫茎泽兰(Eupatorium adenophorumSpreng.)是一种世界性恶性杂草[9-10],于20 世纪40年代传入我国云南南部,现已蔓延到贵州、四川、重庆、广西、西藏以及台湾等地区,对当地农业生产造成了极大的危害[11-13]。紫茎泽兰植株茎叶中含有大量化感物质,对多种植物具有强烈的化感作用,对植物种子萌发和幼苗生长表现出显著的生长抑制作用。因此,紫茎泽兰植株具有开发植物源除草剂的潜力。Ma 等[14]报道,紫茎泽兰叶片浸提液对反枝苋(Amaranthus retroflexus)、灰绿藜(Chenopodium glaucum)杂草种子萌发和幼苗生长有强烈的抑制作用。马金虎等[15]研究发现,紫茎泽兰叶片提取物对稗(Echinochloa crusgalli)、灰绿藜、反枝苋根尖和根边缘细胞具有毒害作用,诱导根边缘细胞凋亡,破坏根的组织结构。杨国庆[16]研究发现,紫茎泽兰的2 个主效化感物质——泽兰二酮[4,7-二甲基-1-(丙烷-2-亚甲基)-1,4,4a,8a-四氢萘-2,6(1H,7H)-二酮]和羟基泽兰酮[6-羟基-5-异丙基-3,8-二甲基-4a,5,6,7,8,8a-六氢奈-2(1H)-酮]对旱稻幼苗具有强烈的化感作用,抑制旱稻幼苗生长。万欢欢等[17]报道,不同浓度紫茎泽兰叶片凋落物水提液对入侵地牧草白三叶(Trifoliumrepens)、辣子草(Galinsoga parvifloral) 和紫花苜蓿(Medicago sativa)种子萌发和幼苗生长有显著的化感抑制作用,且水提液浓度越高抑制效果越强。桂富荣等[18]报道,紫茎泽兰叶片水提液对鲁梅克斯(Rumex patientia)、高丹红(Sorghum vulgare)、鸭茅(Dactylis glomerata)、苕子(Vicia dasycarpa)、胡枝子(Lepedeza bicolor)等9 种牧草种子萌发均具有化感作用,高浓度水提液抑制种子的萌发和幼苗的生长。郑丽等[19]研究发现,高浓度的紫茎泽兰叶片提取液会降低细叶苦荬(Ixeris gracilis)、金盏银盘(Bidens biternata)、莎草砖子苗(Mariscus cyperinus)、无芒虎尾草(Chloris gayana)等10 种牧草种子的发芽率、发芽速率、胚轴和胚根长度,增加幼苗丙二醛含量。钟声等[20]报道,5%的紫茎泽兰茎叶榨取液对我国亚热带种植的多花黑麦草特高(Lolium multiflorumcv.Tetragold)、白三叶、紫花苜蓿等16 种牧草种子的发芽势具有显著的抑制作用。马齿苋(Portulaca oleraceaL.)、鬼针草(Bidens pilosaL.)、鹅绒藤(Cynanchum chinenseR.Br.)和猪毛菜(Salsola collinaPall.)是我国北方农田和果园常见的主要杂草,危害严重,紫茎泽兰叶片提取物对其是否也具有抑制作用尚鲜有报道。本研究以上述4 种杂草为试验材料,研究紫茎泽兰叶片提取物对其种子萌发和早期幼苗生长的的抑制效应及作用机制,旨在为紫茎泽兰叶片提取物对4 种杂草绿色防控方面的应用提供理论依据。
1 材料与方法
1.1 试验材料
1.1.1 紫茎泽兰叶片提取物的制备 采集紫茎泽兰成熟叶片(云南农业大学热带作物学院惠赠),室温下阴干,粉碎机粉碎,过40 目筛,按95%乙醇与植物干粉4 ∶1(v ∶m)浸泡48 h 后,过滤浸提液,重复浸泡3 次后将3 次浸提液混合,用RE-52AA 旋转蒸发器(上海亚荣生化仪器厂)经减压浓缩,除去大部分乙醇,再置于低温真空干燥箱中蒸发掉残留乙醇,至提取物浓缩至膏状且重量不再变化为止[12]。提取物于4℃低温避光保存备用。取适量紫茎泽兰叶片提取物,先用5 mL丙酮溶解,再分别配制成试验所用浓度的紫茎泽兰叶片提取物水溶液。
1.1.2 试验植物 马齿苋、猪毛菜、鬼针草、鹅绒藤种子采自山西农业大学实践教学基地。种子干燥、精选后置冰箱保存备用。
1.2 试验设计
1.2.1 不同浓度紫茎泽兰叶片提取物对杂草种子萌发生长的抑制效应 前期预试验发现,不同的杂草对紫茎泽兰叶片提取物的胁迫敏感程度不同,故紫茎泽兰叶片提取物对不同杂草种子萌发生长抑制效应试验采用了2 种浓度梯度(表1)。挑选均匀饱满的4 种杂草种子适量,先用0.1% HgCl2消毒10 min,然后用蒸馏水冲洗2 次,滤纸吸干种子表面水分。分别取马齿苋、猪毛菜种子约100 粒,鬼针草、鹅绒藤种子各15粒,均匀摆放在18 cm×25 cm 的2 层滤纸中上部,小心将滤纸卷成纸卷(马齿苋、猪毛菜各6 卷,鬼针草、鹅绒藤各3 卷),用橡皮筋将6 卷扎为一组,置于300 mL的烧杯中。分别向烧杯中加100 mL 不同浓度(表1)的紫茎泽兰叶片提取物水溶液,并以加等体积去离子水为对照(CK)。将烧杯置25℃培养箱中黑暗下进行种子发芽试验。种子萌发过程中,隔天更换一次处理液。试验设3 次重复。

表1 紫茎泽兰叶片提取物浓度Table 1 The concentration of E. adenophorum leaf extract
1.2.2 杂草幼苗生长胁迫试验 采用营养钵盆栽试验。取未耕种的黄土状母质上发育的碳酸盐褐土(0 ~20 cm 土层),将土壤晾干捣碎,过3 mm 筛,按土壤和沙7 ∶3加入干净的细河沙混匀,按照2 g·kg-1施入复合肥(N ∶P ∶K=15 ∶15 ∶15)作为土壤基质。称取600 g 土壤基质装入直径14 cm、高16 cm 的塑料营养钵内。播种前每钵先浇200 mL 自来水。取适量均匀饱满、无破损的4 种杂草种子均匀播于营养钵内,然后每钵覆2 cm(约150 g)厚的土壤基质,再浇100 mL 自来水。将营养钵置于室外,在自然条件下进行杂草生长培养。种子出苗后逐渐间苗,每营养钵约留10 株。幼苗生长至苗高10~15 cm 时,将4 种杂草幼苗各分成4 组,每组8 盆。由于紫茎泽兰浸提液对不同杂草抑制强度不同,为了保证杂草能生长,统一选用600 mg·L-1紫茎泽兰叶片提取物每天透灌一次进行胁迫处理,并以自来水透灌作空白对照。分别于胁迫1、2、3、4、5、6 d,取不同处理杂草幼苗中部的成熟叶片,用去离子水冲洗干净,滤纸吸干表面水分,将叶片剪碎混匀用于各项生理指标的测定。试验设3 次重复。
1.3 测定项目与方法
1.3.1 发芽指标测定 种子萌发过程中,每天统计种子的发芽数,直至没有新增种子发芽为止,以种子露白记为萌发。种子发芽结束(6 d),每处理和重复取20株生长一致的幼苗,统计各杂草幼苗的根长、鲜重并根据公式计算各杂草种子的发芽势、发芽率、发芽指数(germination index,GI)、活力指数(vitality index,Ⅵ)、根长抑制率,其中发芽势于萌发试验第3 天计算:

式(3)中,Gt 为t d 内发芽种子数,Dt 为发芽天数;式(4)中,W 为20 株幼苗鲜重。
1.3.2 幼苗叶片生理指标测定及方法 超氧化物歧化酶(superoxide dismutase,SOD)活性测定采用氮蓝四唑法,过氧化物酶(peroxidase,POD)活性测定采用愈创木酚法,过氧化氢酶(catalase,CAT)活性测定采用H2O2紫外吸收法,丙二醛(malondialdehyde,MDA)含量测定采用硫代巴比妥酸法,脯氨酸含量测定采用茚三酮显色法[21]。超氧阴离子自由基(superoxide anion free radical,)含量测定参照李忠光等[22]的方法。还原型抗坏血酸(ascorbic acid,AsA)测定参照邹琦[23]的方法。谷胱甘肽(glutathione,GSH)含量测定参照马金虎[24]的方法。
1.4 数据统计及分析
数据统计分析均采用DPS 6.5 软件完成,采用Duncan 新复极差测验法进行数据差异显著性检验(α=0.05)。利用Excel 2003 软件作图。试验数据均为3次重复的平均值±标准差。
2 结果与分析
2.1 紫茎泽兰叶片提取物对杂草种子萌发和幼苗生长的影响
由表2 可知,随着紫茎泽兰叶片提取物浓度的升高,4 种杂草种子的发芽势、发芽率、发芽指数、活力指数和幼苗鲜重均呈降低趋势,幼苗根长抑制率呈增加趋势。当紫茎泽兰叶片提取物浓度高于400 mg·L-1时,显著抑制了鬼针草、马齿苋、猪毛菜、鹅绒藤种子的萌发生长。与空白对照相比,紫茎泽兰提取物浓度为600 mg·L-1时,鬼针草种子的发芽势和发芽率分别降低了11.5 和14.0 个百分点,发芽指数、活力指数和幼苗鲜重分别降低了56.2%、60.2%和8.7%,根长抑制率达11.0%;马齿苋种子的发芽势和发芽率分别降低了14.5 和10.2 个百分点,发芽指数、活力指数和幼苗鲜重分别降低了32.0%、41.2%和12.0%,根长抑制率达到46.7%;猪毛菜种子的发芽势和发芽率分别降低了44.8 和19.5 个百分点,发芽指数、活力指数和幼苗鲜重分别降低了42.2%、50.0%和16.0%,根长抑制率达28.9%;鹅绒藤种子的发芽势和发芽率分别降低了71.0 和6.5 个百分点,发芽指数、活力指数和幼苗鲜重分别降低了40.4%、67.0%和45.0%,根长抑制率达到62.0%。上述结果表明,紫茎泽兰叶片提取物对4种杂草种子萌发和幼苗生长均有抑制作用。
2.2 紫茎泽兰叶片提取物对杂草幼苗抗氧化系统的影响
2.2.1 紫茎泽兰叶片提取物对杂草幼苗叶片SOD 活性的影响 试验用600 mg·L-1紫茎泽兰叶片提取物对杂草幼苗进行胁迫处理,分别在胁迫处理1、2、3、4、5、6 d 时,检测幼苗叶片SOD 活性。结果表明,提取物胁迫处理2、3、4、5、6 d 时,马齿苋幼苗叶片SOD 活性均显著高于其空白对照,其中胁迫处理3 d 时,叶片SOD活性最高,较其空白对照提高91.11%(图1-A);提取物胁迫处理1、3、4、5 d 时,猪毛菜叶片SOD 活性分别较其空白对照显著提高16.35%、38.93%、39.83%和45.37%(图1-B);提取物胁迫处理3、4、5、6 d 时,鬼针草叶片SOD 活性分别较其空白对照显著提高了21.48%、41.01%、54.43%和83.26%(图1-C);提取物胁迫处理1、4、5、6 d 时,鹅绒藤叶片SOD 活性分别较其空白对照显著提高了23.07%、12.46%、34.03%和25.48%(图1-D)。4 种杂草经紫茎泽兰叶片提取物处理,在不同处理时间幼苗叶片SOD 活性出现显著差异。综上,紫茎泽兰叶片提取物对4 种杂草均产生了胁迫伤害,诱导了杂草幼苗叶片SOD 活性的升高。
2.2.2 紫茎泽兰叶片提取物对杂草幼苗叶片POD 活性的影响 试验用600 mg·L-1紫茎泽兰叶片提取物对杂草幼苗进行胁迫处理,分别于胁迫1、2、3、4、5、6 d时,检测幼苗叶片POD 活性。结果表明,提取物胁迫处理1、2、3、4、5 d 时,马齿苋叶片POD 活性均显著高于其空白对照,胁迫处理3 d 时,POD 活性较其空白对照提高了48.72%(图2-A);提取物胁迫处理5、6 d时,猪毛菜POD 活性分别较其空白对照提高了17.12%和18.76%(图2-B);提取物胁迫处理1、3、4、5、6 d 时,鬼针草叶片POD 活性分别较其空白对照显著提高了 35.22%、 72.01%、 40.32%、 66.86% 和59.39%(图2-C);提取物胁迫处理1、4 d 时,鹅绒藤叶片POD 活性较其空白对照显著提高了47.80%和22.98%(图2-D)。4 种杂草经紫茎泽兰叶片提取物处理,在不同处理时间幼苗叶片POD 活性出现显著差异。综上,紫茎泽兰叶片提取物对4 种杂草均产生了胁迫伤害,诱导杂草幼苗叶片POD 活性升高。
2.2.3 紫茎泽兰叶片提取物对杂草幼苗叶片CAT 活性的影响 试验用600 mg·L-1紫茎泽兰叶片提取物对杂草幼苗进行胁迫处理,分别在1、2、3、4、5、6 d 时,检测幼苗叶片CAT 活性。结果表明,提取物胁迫处理1、2、3、4、5、6 d 时,马齿苋叶片CAT 活性分别较其空白对照显著提高了32.02%、57.27%、30.41%、51.51%、81.34%和43.37%(图3-A);提取物胁迫处理1、2、6 d时,猪毛菜叶片CAT 活性分别较其空白对照显著提高了32.42%、47.11%和34.00%(图3-B);提取物胁迫处理3 d 时,鬼针草叶片CAT 活性较其空白对照显著提高了17.05%(图3-C);提取物胁迫处理1、2 d 时,鹅绒藤叶片CAT 活性分别较其空白对照显著提高了23.42%和16.88%(图3-D)。4 种杂草经紫茎泽兰叶片提取物处理,不同处理时间幼苗叶片CAT 活性出现显著差异,说明紫茎泽兰叶片提取物对4 种杂草均产生胁迫伤害,诱导杂草幼苗叶片CAT 活性升高。
2.2.4 紫茎泽兰叶片提取物对杂草幼苗叶片抗坏血酸(AsA)含量的影响 由图4 可知,紫茎泽兰叶片提取物处理后,4 种杂草幼苗叶片AsA 含量在不同时间段均高于其空白对照。提取物胁迫处理下,马齿苋叶片AsA 含量与其空白对照相比均未达显著水平(图4-A);猪毛菜在提取物胁迫处理1、2、3、4、5、6 d 时,其叶片AsA 含量均显著高于其空白对照,分别提高了41.22%、 105.0%、 54.14%、 96.33%、 46.78% 和90.71%(图4-B);提取物胁迫处理1、6 d 时,鬼针草叶片AsA 含量分别较其空白对照提高了28.33%和33.48%(图4-C);提取物胁迫处理1、2、4、5、6 d 时,鹅绒藤叶片AsA 含量分别较其空白对照显著提高了17.10%、19.68%、44.00%、43.43%和60.74%(图4-D)。幼苗叶片AsA 含量出现显著差异,说明紫茎泽兰叶片提取物对4 种杂草均产生了胁迫伤害,诱导杂草幼苗叶片AsA 含量升高。
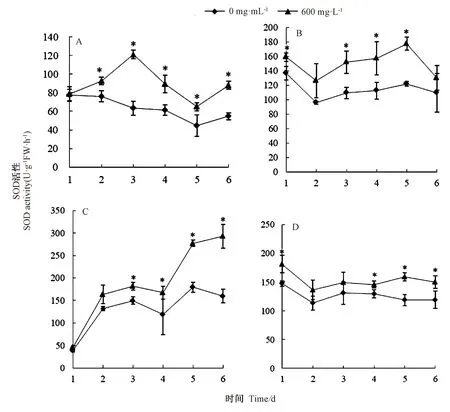
图1 紫茎泽兰叶片提取物对4 种杂草幼苗叶片SOD 活性的影响Fig.1 The effect of E. adenophorum leaf extract on SOD activity in leaf of four spices of weed

图2 紫茎泽兰叶片提取物对4 种杂草幼苗叶片POD 活性的影响Fig.2 The effect of E. adenophorum leaf extract on POD activity in leaf of four spices of weed

图3 紫茎泽兰叶片提取物对4 种杂草幼苗叶片CAT 活性的影响Fig.3 The effect of E. adenophorum leaf extract on CAT activity in leaf of four spices of weed

图4 紫茎泽兰叶片提取物对4 种杂草幼苗叶片AsA 含量的影响Fig.4 The effect of E. adenophorum leaf extract on AsA content in leaf of four spices of weed
2.2.5 紫茎泽兰叶片提取物对杂草幼苗叶片谷胱甘肽(GSH)含量的影响 由图5 可知,紫茎泽兰叶片提取物处理后,供试的4 种杂草幼苗叶片GSH 含量均高于其空白对照。提取物胁迫处理1、2、3、4、6 时,马齿苋叶片GSH 含量均显著高于其空白对照,其中胁迫处理4 d 时较其空白对照提高了30.58%(图5-A);猪毛菜在提取物胁迫处理1、2、3、4、5、6 d 时,GSH 含量分别比其对照显著提高24.21%、39.30%、21.73%、37.53%、57.49%和45.97%(图5-B);鬼针草在提取物胁迫处理下,GSH 含量均显著高于其空白对照,其中胁迫处理6 d 时较其空白对照提高了60.60%(图5-C);提取物胁迫下,鹅绒藤的GSH 含量较其空白对照均未达到差异显著水平(图5-D)。紫茎泽兰叶片提取物对4 种杂草均产生了胁迫伤害,诱导杂草幼苗叶片GSH 含量升高。
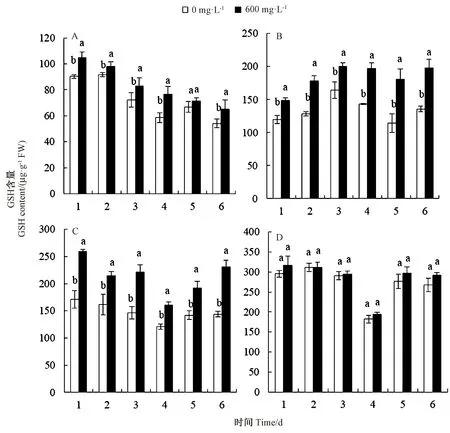
图5 紫茎泽兰叶片提取物对4 种杂草幼苗叶片GSH 含量的影响Fig.5 The effect of E. adenophorum leaf extract on GSH content in leaf of four spices of weed
2.2.6 紫茎泽兰叶片提取物对杂草幼苗叶片MDA 含量的影响 由图6-A 可知,紫茎泽兰叶片提取物胁迫处理1、4 d 时,马齿苋MDA 含量分别较其空白对照提高了64.58%和30.56%;提取物胁迫处理后,猪毛菜MDA 含量均较其空白对照高,但差异不显著(图6-B);提取物胁迫处理3、4、6 d 时,鬼针草MDA 含量分别较其空白对照提高了19.05%、21.55%和27.61%(图6-C);提取物胁迫处理3、4、6 d 时,鹅绒藤MDA含量分别较其空白对照提高了44.19%、69.70%和60.95%(图6-D)。综上,紫茎泽兰叶片提取物对4 种杂草均产了生胁迫伤害,诱导杂草幼苗叶片MDA 含量升高。

图6 紫茎泽兰叶片提取物对4 种杂草幼苗叶片MDA 含量的影响Fig.6 The effect of E. adenophorum leaf extract on MDA content in leaf of four spices of weed
2.2.7 紫茎泽兰叶片提取物对杂草幼苗叶片超氧阴离子(含量的影响 如图7 所示,紫茎泽兰叶片提取物胁迫处理后,供试的4 种杂草幼苗叶片含量均高于其空白对照处理。提取物胁迫处理2、5、6 d时,马齿苋含量分别较其空白对照提高了34.63%、29.81%和31.93%(图7-A);猪毛菜在提取物胁迫处理4、5、6 d 时,含量分别较其空白对照提高了25.02%、39.78%和47.58%(图7-B);鬼针草在提取物胁迫处理3、4 d 时,含量分别较其空白对照提高了21.55%和19.04%(图7-C);鹅绒藤在提取物胁迫处理1、2、3、4、5、6 d 时,含量较其空白对照高,但差异均不显著(图7-D)。4 种杂草经紫茎泽兰叶片提取物处理,在不同处理时间幼苗叶片含量出现显著差异。说明紫茎泽兰叶片提取物对4 种杂草均产生了胁迫伤害,诱导杂草幼苗叶片含量升高。
2.2.8 紫茎泽兰叶片提取物对杂草幼苗叶片脯氨酸含量的影响 由图8-A 可知,提取物胁迫处理2、5 d时,马齿苋脯氨酸含量分别较其空白对照提高了86.12%和89.06%;提取物胁迫处理1、2、3 d 时,猪毛菜脯氨酸含量分别较其空白对照显著提高了248.24%、74.40%和68.91%(图8-B);提取物胁迫处理3、4、5、6 d 时,鬼针草脯氨酸含量分别较其空白对照显著提高了190.90%、 80.41%、 210.70% 和570.10%(图8-C);提取物胁迫处理后,鹅绒藤脯氨酸含量均显著高于其空白对照,其中胁迫处理1、3 d 时,脯氨酸含量分别比对照提高了92.53%、91.00%(图8-D)。4 种杂草经紫茎泽兰叶片提取物处理,在不同处理时间幼苗叶片脯氨酸含量出现显著差异。紫茎泽兰叶片提取物对4 种杂草均产生了胁迫伤害,诱导杂草幼苗叶片脯氨酸含量升高。
3 讨论

图7 紫茎泽兰叶片提取物对4 种杂草幼苗叶片含量的影响Fig.7 The effect of E. adenophorum leaf extract oncontent in leaf of four spices of weed
化学农药本身固有的缺陷和人们长期不合理的使用,不仅对生态环境造成严重污染,其农药残留也对农产品质量和人类健康带来了一定的安全隐患。因此,亟待发掘有同样作用的天然活性物质——绿色植物源农药。据报道,全球具有控制有害生物作用的高等植物有6 300 多种,其中,除草植物有1 000 多种[25]。研究发现,紫茎泽兰叶片提取物对旱稻、牧草等植物种子萌发和幼苗生长发育有显著的抑制作用[16-18]。本研究结果表明,随着紫茎泽兰叶片提取物浓度的增加,鹅绒藤、猪毛菜、马齿苋和鬼针草4 种杂草的发芽率、发芽势、发芽指数、活力指数均逐渐降低,幼苗的根长抑制率增大,幼苗鲜重降低,说明紫茎泽兰叶片提取物对4 种杂草种子萌发和幼苗生长发育均有明显的抑制作用,提取物可用于植物源除草剂的研发。当紫茎泽兰叶片提取物浓度为600 mg·L-1时,鹅绒藤和猪毛菜的幼苗鲜重分别比其空白对照下降45.0%和16.0%,而马齿苋、鬼针草幼苗的鲜重则比其空白对照下降了12.0%和8.7%,说明4 种杂草对紫茎泽兰提取物的敏感性不同。相对来说,鹅绒藤、猪毛菜对提取物比较敏感,而鬼针草、马齿苋对提取物有较强的耐性。这与钟声等[20]和桂富荣等[18]的研究结果相似。究其原因,可能是4 种杂草分属不同的4 个科属,其生物学特性存在差异的结果。
当植物受到外界逆境胁迫时,细胞中活性氧(reactive oxidative species,ROS),如等含量会升高,诱发细胞膜发生膜脂过氧化反应,破坏膜的完整性,同时产生大量MDA。因此,细胞内MDA 含量可反映细胞膜氧化损伤的程度[26-27]。孙娜娜[12]研究发现,1 000 mg·L-1紫茎泽兰叶片提取物水溶液处理可对黄瓜根系产生氧化胁迫伤害,破坏根的组织结构,显著诱导黄瓜根系O-·2 、MDA 含量升高。本试验结果也发现,600 mg·L-1的紫茎泽兰叶片提取物胁迫下,4 种杂草幼苗叶片、MDA 含量在不同处理时间段均较其空白对照高,说明提取物同样对杂草幼苗也产生了氧化胁迫伤害。
SOD、POD、CAT 等抗氧化酶构成了植物的抗氧化保护系统。适宜环境条件下,植物体内抗氧化酶活性维持在较低的水平,当植物遭受逆境胁迫时,抗氧化酶活性会升高,保护植物免受ROS 的伤害[28]。SOD 是植物抗氧化系统的第一道防线,它可以使梅勒反应中产生的ROS 转化成H2O2,然后通过POD、CAT 等将H2O2转化为H2O 和O2,从而有效地阻止和H2O2对细胞膜产生更大的伤害[29]。本研究发现,4 种杂草在紫茎泽兰叶片提取物处理后,不同杂草、不同胁迫时间段SOD、POD、CAT 活性均比其空白对照高,是杂草主动抵御胁迫伤害的一种表现,这与前人的研究结果一致[30]。随着紫茎泽兰提取物胁迫时间的延长,4 种杂草SOD、POD、CAT 活性有的表现出活性进一步升高,有的则迅速降低,这可能是因为不同杂草对紫茎泽兰提取物胁迫敏感程度不同,这从紫茎泽兰提取物胁迫下4 种杂草萌发生长指标变化中也能得到印证,说明紫茎泽兰提取物胁迫对4 种杂草均造成了不同程度的胁迫伤害。
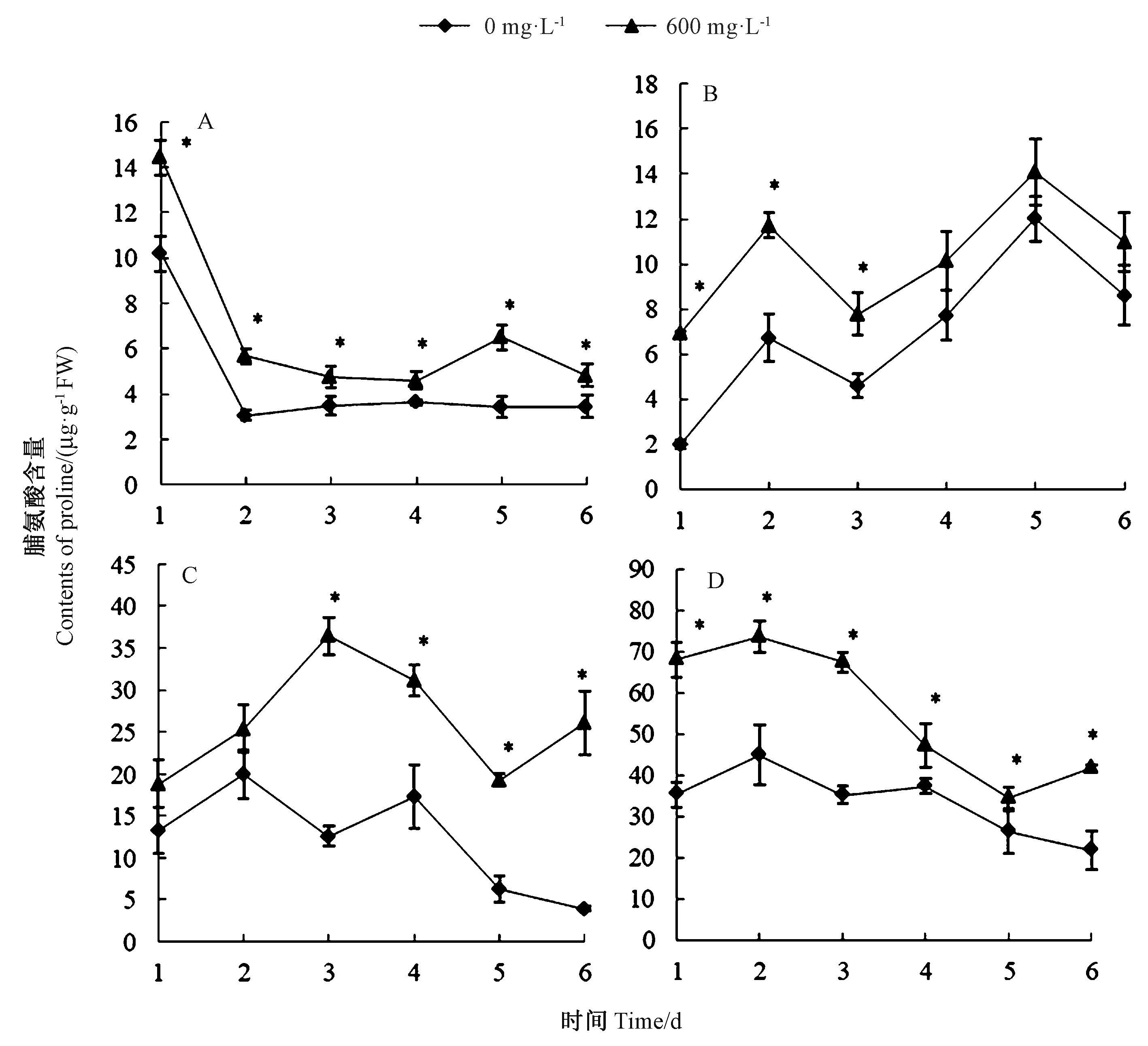
图8 紫茎泽兰叶片提取物对4 种杂草幼苗叶片脯氨酸含量的影响Fig.8 The effect of E. adenophorum leaf extract on proline content in leaf of four spices of weed
植物体内除酶促活性氧清除系统外还有非酶类抗氧化物质,如GSH、AsA 等,可有效抵抗逆境胁迫所造成的伤害[31]。当植物受到逆境胁迫时,ROS 含量会逐渐升高,激发植物细胞中谷胱甘肽和抗坏血酸防御系统启动反应,产生更多的GSH 和AsA,抵抗逆境胁迫伤害。本研究中,紫茎泽兰叶片提取物胁迫处理后,在不同胁迫时间段,4 种杂草幼苗GSH 和AsA 含量均较其空白对照高,进一步印证了提取物处理对杂草幼苗产生了逆境胁迫伤害。
逆境胁迫下,积累渗透调节物质也是植物应对外界胁迫所导致的细胞内外渗透压失衡的方法之一,脯氨酸则是其中一种主要渗透调节物质[32-34]。本研究结果显示,紫茎泽兰叶片提取物胁迫处理后,在不同胁迫时间段,4 种杂草幼苗体内脯氨酸含量均较其空白对照高,说明杂草在受到提取物胁迫时可自身积累一定量的脯氨酸来调节细胞渗透压,减缓环境胁迫的影响。这也从脯氨酸的渗透调节作用方面,证明了提取物对杂草有胁迫伤害作用。
4 结论
紫茎泽兰叶片提取物对鹅绒藤、猪毛菜、马齿苋和鬼针草4 种杂草种子萌发和幼苗生长均有抑制作用,提取物浓度越高抑制作用越强。此外,这种抑制效应存在差异,表现为鹅绒藤>猪毛菜>马齿苋>鬼针草。紫茎泽兰叶片提取物对4 种杂草的氧化胁迫作用,是导致种子萌发率降低和幼苗生长受抑的内在机制,但紫茎泽兰叶片提取物对不同杂草的抑制程度不同,这一现象是否与植株属于不同科属相关,还有待进一步研究证实。
