A new species of Kurixalus (Anura, Rhacophoridae)from Guizhou, China
2021-04-15JuanZeng,Ji-ShanWang,Guo-HuaYu等
DEAR EDITOR,
We describe a new species of the genusKurixalus,Kurixalus raoisp.nov., from Guizhou Province, China, based on morphological and molecular evidence.Phylogenetically, the new species is sister toK.idiootocus, but is distinguishable from all known congeners by a combination of the following characters: small body size (snout-vent length 28.2-32.2 mm in males; 38.6 mm in female); snout rounded, with no prominence on tip; single internal vocal sac; dorsal surface brown, rough, scattered with several small warts; chin clouded with blackish marking; pair of large symmetrical dark blotches on chest; vomerine teeth present; iris brown; tibiotarsal articulation reaching center of eye; nuptial pad slight; flank rough; mandibular symphysis weak; throat skin granular; and toes moderately webbed, formula I2-2II1.5-3III2-3IV3-2V.The genetic distances between the new species andK.idiootocuswere 2.9% and 5.4% for 16S rRNA and COI,respectively.
KurixalusYe, Fei & Dubois, a genus within the family Rhacophoridae, is widely distributed from northeastern India,eastwards to Indochina, southern mainland China, adjacent continental islands (Hainan, Taiwan), and the Ryukyu archipelago, and southwards to the Sunda and Philippine archipelagos (Frost, 2020).Currently, a total of 19 species are recognized in the genusKurixalus, nine of which have been recorded from China, namely:K.beryllinirisWu, Huang, Tsai,Li, Jhang & Wu,K.eiffingeri(Boettger),K.hainanus(Zhao,Wang & Shi),K.idiootocus(Kuramoto & Wang),K.lenquanensisYu, Wang, Hou, Rao & Yang,K.naso(Annandale),K.odontotarsus(Ye & Fei),K.wangiWu,Huang, Tsai, Li, Jhang & Wu, andK.yangiYu, Hui, Rao &Yang (Frost, 2020; Yu et al., 2017a, 2018).
At present, a single species of the genus (K.hainanus) is known from Guizhou Province, China (Yu et al., 2017a).During our recent field surveys in southwest Guizhou, we collected several small-sized tree frog specimens resembling members ofKurixalusbased on various characters, notably presence of serrated dermal fringes along outer edge of forearm and tarsus and saddle-shaped mark on dorsum.Phylogenetically, these specimens represent a new lineage,which is sister toK.idiootocus(currently endemic in Taiwan),but morphologically distinguishable fromK.idiootocusand other members ofKurixalusby a series of characters.Therefore, we formally describe this lineage as a new species ofKurixalus.
Field surveys were conducted in May 2019 at Qingshuihe Nature Reserve, Southwest Guizhou Autonomous Prefecture,Guizhou, China (Figure 1A).Specimens were photographed,euthanized, fixed, and then stored in 75% ethanol.Liver tissues were preserved in 99% ethanol.Specimens were deposited at Guangxi Normal University (GXNU), Guangxi,China.
Sequences encoding three mitochondrial genes (12S rRNA,16S rRNA, andCOI) were amplified and sequenced using the primers and experimental protocols of Yu et al.(2020).Homologous sequences of otherKurixalusspecies were obtained from GenBank.All new sequences were deposited in GenBank under Accession Nos.MW345613-MW345628 and MW346332 -MW346339 (Supplementary Table S1).Phylogenetic relationships among the genusKurixaluswere inferred using Bayesian inference (Supplementary Materials and Methods).Uncorrected pairwise distances between species were calculated in MEGA 7 (Kumar et al., 2016).
Morphometric data were taken using digital calipers to the nearest 0.1 mm.Morphological terminology followed Fei et al.(2017) (Supplementary Materials and Methods).Comparative morphological data of otherKurixalusspecies were taken from previous publications (Annandale, 1912; Bossuyt & Dubois,2001; Boulenger, 1893; Günther, 1858; Inger et al., 1999;Kuramoto & Wang, 1987; Mediyansyah et al., 2019; Nguyen et al., 2014a, 2014b; Smith, 1924; Taylor, 1962; Van Nguyen et al., 2020; Wu et al., 2016; Yu et al., 2017b, 2018; Zhao et al., 2005).Multivariate principal component analysis (PCA)was conducted using SPSS 17.0 (SPSS Inc., USA) based on a correlation matrix of size-standardized measurements (all measurements divided by SVL).Scatter plots of the first two PCA factors were used to examine the differentiation between the new species andK.idiootocus.Specimens ofK.idiootocuswere collected from Xinbei, Taiwan, China, and can be easily distinguished from otherKurixalusspecies in Taiwan by the presence of two large brownish rounded blotches on the chest (Wu et al., 2016).

Figure 1 Collection site (A), habitat (B), phylogenetic position (C), and scatterplot of morphometric data of Kurixalus raoi sp.nov.(D)
The obtained sequence alignments were 401 bp, 873 bp,and 807 bp for 12S rRNA, 16S rRNA, andCOIgenes,respectively.Phylogenetic analysis of the three genes strongly supported the new specimens as a distinct lineage of the genusKurixalusand sister toK.idiootocus(Figure 1C).The genetic distances between the new lineage andK.idiootocuswere 2.9% and 5.4% for 16S rRNA andCOI, respectively(Supplementary Table S2).
Morphometric data are summarized in Table 1.We retained the first two principal components, which accounted for 65.97% of total variance and had eigenvalues above 1.0.Loadings for PC1 were positive, except for distance between nostril and eye (DNE), and were most heavily loaded on head length (HL), upper eyelid width (UEW), and eye diameter (ED)(Table 2).Differentiation was found along the PC1 axis between the new lineage andK.idiootocus(Figure 1D).The PCA results indicated that the new lineage differs morphometrically from its sister species (K.idiootocus) by shorter head, narrower UEW, and smaller eye.In addition, the new lineage possesses a combination of morphologicalcharacters different from all known congeners (Supplementary Table S3).Therefore, based on molecular and morphological evidence, we describe the specimens as a new species of the genusKurixalusbelow.
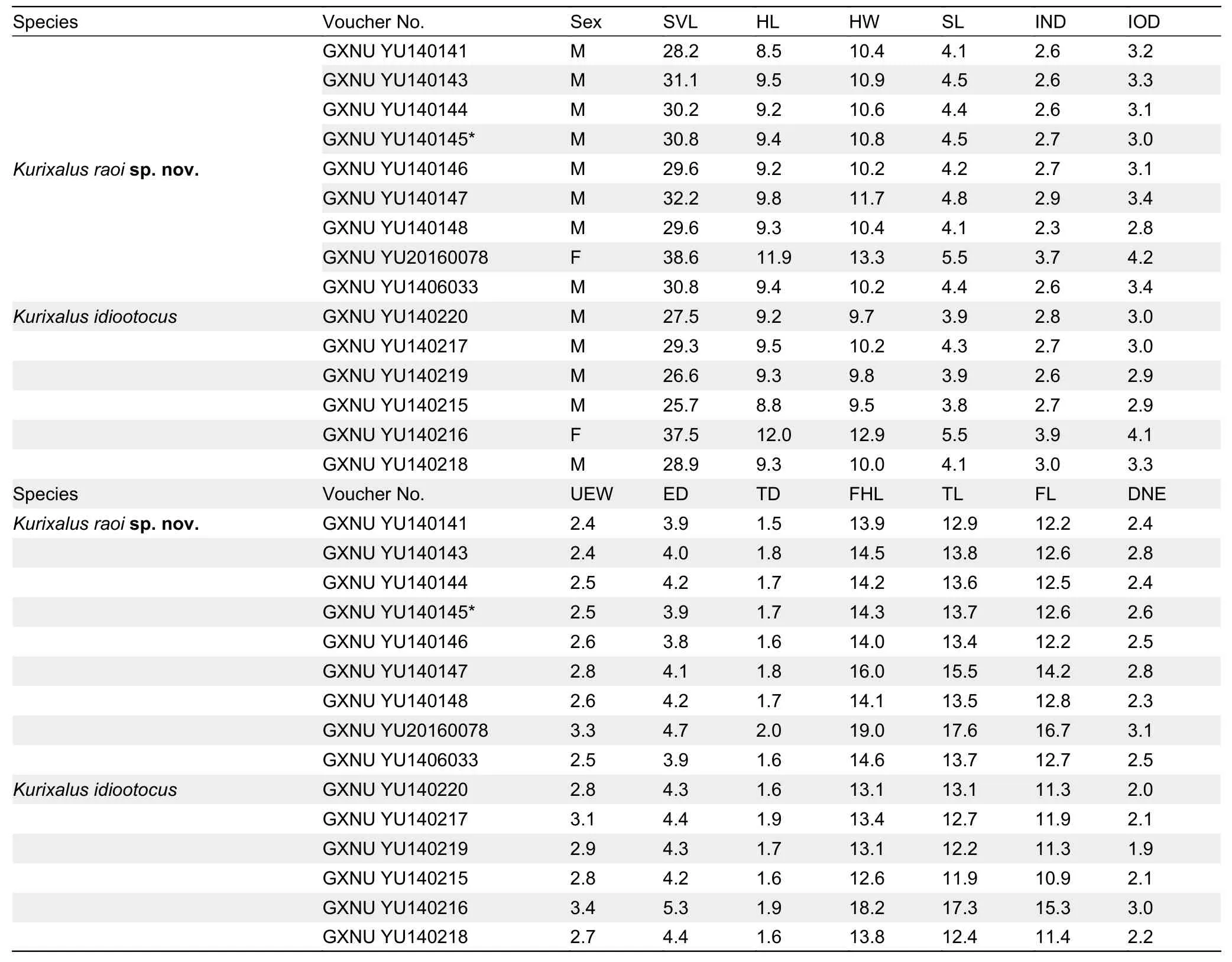
Table 1 Measurements (in mm) of Kurixalus raoi sp.nov.and K.idiootocus collected from Xinbei, Taiwan, China
Taxonomic account
Kurixalus raoisp.nov.(Figure 2)
Rao’s frilled treefrog/Ráo Shì Yuán Zhǐ Shù Wā (饶氏原指树蛙)
Holotype:GXNU YU140145, adult male, collected on 13 May 2019 by Guo-Hua Yu from Qingshuihe Nature Reserve,Southwest Guizhou Autonomous Prefecture, Guizhou Province, China (N25°18", E104°57", 1 290 m a.s.l.).
Paratypes:Seven adult males (GXNU YU140141, GXNU YU140143, GXNU YU140144, GXNU YU140146-YU140148,and GXNU YU1406033) and one adult female (GXNU YU20160078) with same collection information as holotype.
Etymology:The species name is dedicated to Dr.Ding-Qi Rao from the Kunming Institute of Zoology, Chinese Academy of Sciences, for his support of our research over the years.
Diagnosis:The new tree frog species is assigned toKurixalusbased on the following characters: small body size (SVL<50 mm); finger webbing poorly developed and toe webbing moderately developed; serrated dermal fringes along outer edge of forearm and tarsus; inverted triangle-shaped dark brown mark between eyes; dorsal brown “) (” saddle-shaped marking; coarse dorsal and lateral surface with tubercles(Nguyen et al., 2014a, 2014b; Yu et al., 2017b, 2018).Phylogenetic analysis supported that the new species belongs to the genusKurixalusand is sister toK.idiootocus(Figure 1C).
Kurixalus raoisp.nov.can be distinguished from its congeners by a combination of the following characters: small body size (mean SVL 30.3 mm (28.2-32.2 mm) in males, SVL 38.6 mm in a single female); snout obtuse, no prominence ontip; canthus rostralis curved; dorsal surface brown, rough,scattered with several small warts; chin clouded with blackish marking; ventral surface granular, no large dark spots on ventral surface; vomerine teeth present; iris brown; single internal vocal sac; dermal fringes present along outer edge of limbs; tibiotarsal articulation reaching center of eye; nuptial pad slight; flank rough; mandibular symphysis weak; toes moderately webbed, formula I2-2II1.5-3III2-3IV3-2V.
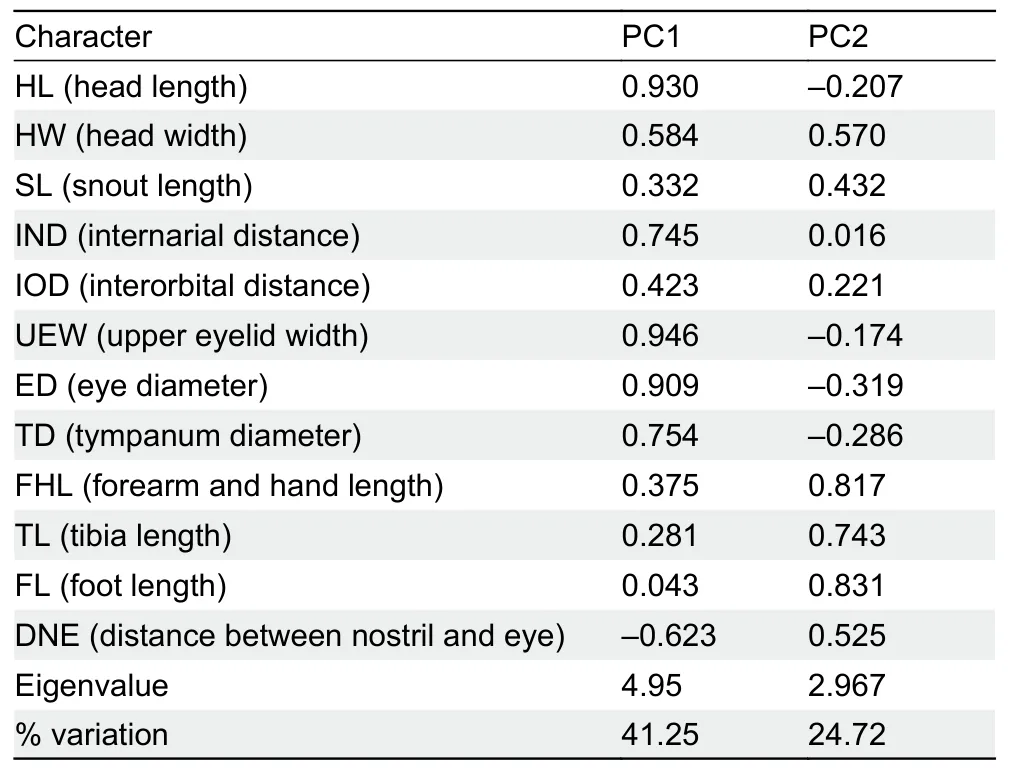
Table 2 Factor loadings of first two principal components of 12 size-adjusted morphometric characteristics of Kurixalus raoi sp.nov.and K.idiootocus
Description of holotype:Adult male, body size small (SVL 30.8 mm); HL 87% of HW (head width); snout obtuse with no dermal prominence on tip; snout longer than eye (SL 4.5 mm,ED 3.9 mm); canthus rostralis blunt and curved; lore region oblique, slightly concave; nostril oval, slightly protuberant,closer to snout tip than eye; IND (internarial distance; 2.7 mm)narrower than IOD (interorbital distance; 3.0 mm); pineal spot absent; pupil oval, horizontal; tympanum distinct (TD 1.7 mm),rounded, less than half ED; supratympanic fold distinct, curves from posterior edge of eye to insertion of arm; vomerine teeth in two oblique series touching inner front edge of choanae,separated by space almost equal to length of each series;tongue attached anteriorly and notched posteriorly; single internal vocal sac, with sac slit opening on floor of mouth at each corner.
Relative length of fingers: I<II<IV<III; tips of all fingers expanded into discs with circummarginal grooves; nuptial pad present on first finger; fingers weakly webbed at base; lateral fringe on free edge of all fingers; subarticular tubercles prominent and rounded, formula 1, 1, 2, 2; supernumerary tubercles present; one inner metacarpal tubercle, outer metacarpal tubercle divided into two; row of white warts forming serrated fringe along outer edge of forearm.

Figure 2 Dorsal view of holotype (GXNU YU140145) in life (A) and preservative (B) and ventral views of holotype (C) and paratypes GXNU YU140141 (D), GXNU YU140143 (E), and GXNU YU20160078 (F)
Heels overlapping when legs positioned at right angles to body; tibiotarsal articulation reaching center of eye; relative length of toes: I<II<V<III<IV; tibia longer than foot (TL 13.7 mm, FL 12.6 mm); tips of toes expanded into discs with circummarginal grooves, smaller than finger discs; toes webbed moderately, formula I2-2II1.5-3III2-3IV3-2V;subarticular tubercles prominent and rounded, formula 1, 1, 2,3, 2; supernumerary tubercles present; series of white tubercles forming serrated dermal fringes along outer edge of tarsus and fifth toe; inner metatarsal tubercle oval, outer metatarsal tubercle absent.
Dorsal surface rough with several small warts on top of head, upper eyelids, dorsum, flank, and dorsal surface of limbs; sides of head scattered with several small tubercles;few white tubercles below anal opening; white conical tubercle on tibiotarsal articulation; throat, chest, belly, and ventral surface of thigh granular, more so on venter.
Coloration in life:Iris brown; dorsal surface grayish-brown,dark brown saddle-shaped mark on dorsum behind eye; dark inverted triangular-shaped mark between eyes; side of head and tympanic region grayish-brown; clear dark brown bar along canthus rostralis; flank gray-brown mottled with faint yellow; limbs dorsally gray-brown with dark brown band;anterior and posterior sides of thigh yolk yellow with small dark patches; chest and abdomen white with faint dark patches;chin completely clouded with black.
Coloration in preservative:Dorsal ground color brown,pattern as in life.Chest white with pair of symmetrical faint dark patches on each side; abdomen white with faint dark speckles.
Morphological variation:The new species is sexually dimorphic, with the female being obviously larger than the male (Table 1).Males possess a single internal vocal sac and slight nuptial pad.Color of ventral surface also varies among individuals: for example, dark blotches and speckles on ventral surface are faint in GXNU YU140145-YU140147, but clear in other paratypes (Figure 2).
Distribution and ecology:The new species is known only from Qingshuihe Nature Reserve, located in a karst area of Southwest Guizhou, China.Males were found sitting on shrubs, rocks, and grass.The gravid female (GXNU YU20160078) was found on the ground.No tadpoles were collected.
Conservation status:Kurixalus raoisp.nov.is distributed in a karst area with low surface water storage ability.Moreover,the type locality is encroached by livestock overgrazing and expanding agriculture.During our field surveys, we only found a few small pools with little water in the lowlands, which were all heavily disturbed by livestock (especially buffalo).Given the likely small distributional range and drought and overgrazing in the area, we recommend that the new species be considered Near Threatened (NT) following IUNC’s Red List categories(IUCN Standards and Petitions Subcommittee, 2016).
Comparisons:Morphological comparisons between the new species and all recognized congeners are summarized in Supplementary Table S3.Morphologically, the new species can be distinguished from its closest relative,K.idiootocus,based on the following characters: larger body size in males(28.2-32.2 mm vs.24.9-29.3 mm); snout obtuse, rounded with no dermal prominence on tip (vs.pointed with small dermal prominence on tip; Figure 3); single internal vocal sac(vs.single external vocal sac); and dorsal surface grayish brown (vs.golden brown).
Kurixalus raoisp.nov.is distinguishable fromK.lenquanensisby larger body size in males (28.2-32.2 mm vs.25.0-28.9 mm); pair of large symmetrical dark blotches on chest present (vs.absent); smoother dorsal surface (vs.many small warts on top of head, upper eyelid, and dorsum;Figure 3); and SL (snout length) greater than ED (vs.SL less than ED).
The new species can be distinguished fromK.absconditusMediyansyah, Hamidy, Munir & Matsui,K.appendiculatus(Güther),K.baliogaster(Inger, Orlov & Darevsky),K.banaensis(Bourret),K.berylliniris,K.bisacculus(Taylor),K.chaseni(Smith),K.eiffingeri,K.hainanus,K.motokawai(Nguyen, Matsui & Eto),K.naso,K.odontotarsus,K.viridescens(Nguyen, Matsui & Hoang),K.wangi, andK.yangiby snout rounded with no projection on tip (vs.pointed with projection on tip) and pair of large symmetrical dark blotches on chest present (vs.absent); fromK.gracilloidesby ground color of dorsal surface grayish brown (vs.golden-brown), pair of large symmetrical dark blotches on chest present (vs.absent), and tibiotarsal articulation reaching center of eye (vs.reaching between eye and nostril); and fromK.verrucosus(Boulenger) by smaller body size in females (38.6 mm vs.43-45 mm), pair of large symmetrical dark blotches on chest present (vs.absent), throat and chest granular (vs.smooth),tibiotarsal articulation reaching center of eye (vs.reaching between eye and nostril), and toe webbing less developed,formula I2-2II1.5-3III2-3IV3-2V (vs.nearly entirely webbed,formula I1-2II1-2III1-2IV2-1V).

Figure 3 Dorsal views of Kurixalus raoi sp.nov.(A; GXNU YU140146), K.lenquanensis (B; GXNU YU20160046, male,collected from Mengzi, Yunnan, China), and K.idiootocus (C;GXNU YU140219, male, collected from Xinbei, Taiwan, China) and ventral view of K.idiootocus (D; GXNU YU140219)
Comments:The taxonomy in the genusKurixalusis complicated due to remarkable morphological similarities among species.Recently, however, a series of taxonomic papers relying on both morphological and molecular evidence have been published (Matsui et al., 2018; Mediyansyah et al.,2019; Van Nguyen et al., 2020; Wu et al., 2016; Yu et al.,2017a, 2017b, 2018).Genetically,Kurixalus raoisp.nov.shows a 16S rRNA sequence divergence of 2.9% from its closest relativeK.idiootocus, which is nearly equal to the conventional threshold of species-level divergence in Anura(3.0% divergence in 16S rRNA gene; Vences et al., 2005).Morphologically, these two species differ from each other in a series of characters, i.e.: vocal sac, body size, and snout shape.Therefore, we consider thatKurixalus raoisp.nov.deserves the rank of independent species.
The new species described here is the third Asian mainlandKurixalusspecies found to be phylogenetically closer to its congeners on the East Asian islands than its mainland congeners, the other two species beingK.lenquanensisfrom Yunnan andK.gracilloidesfrom Vietnam.As expected in Yu et al.(2017b), the discovery ofKurixalus raoisp.nov.suggests the probability that moreKurixalusspecies allied to the East Asian islands clade will be found from southern China pending field surveys, considering the huge geographical gap in distribution between the three mainland species and their relatives from Taiwan Island and the climatic and habitat diversity.In addition, the relationships amongK.gracilloides,the clade consisting ofK.lenquanensis,K.idiootocusandKurixalus raoisp.nov., and the clade consisting ofK.eiffingeri,K.berylliniris, andK.wangias well as the relationships among the three Vietnamese species (K.motokawai,K.banaensis, andK.viridescens) remain unresolved (Figure 1C).Furthermore,K.verrucosus(from the type locality) has never been included in any phylogenetic analysis according to Yu et al.(2017a).Therefore, a phylogenetic study employing more molecular markers andK.verrucosusis necessary to obtain a full understanding of the phylogeny of the genusKurixalus.
TakingKurixalus raoisp.nov.into account, the genusKurixalusnow contains 20 known species.The species diversity ofKurixalusis still underestimated and further morphological studies are needed as several unnamed lineages in theK.odontotarsusspecies group remain according to Yu et al.(2017a).
NOMENCLATURAL ACTS REGISTRATION
The electronic version of this article in portable document format represents a published work according to the International Commission on Zoological Nomenclature (ICZN),and hence the new names contained in the electronic version are effectively published under that Code from the electronic edition alone (see Articles 8.5-8.6 of the Code).This published work and the nomenclatural acts it contains have been registered in ZooBank, the online registration system for the ICZN.The ZooBank LSIDs (Life Science Identifiers) can be resolved and the associated information can be viewed through any standard web browser by appending the LSID to the prefixhttp://zoobank.org/.
Publication LSID:
urn:lsid:zoobank.org:pub:15D25245-AC2B-4DD0-B813-2853 573730DE
Kurixalus raoiLSID:
urn:lsid:zoobank.org:act:9562B6BF-19B9-4924-895E-4C849 D4F7E95
SCIENTIFIC FIELD SURVEY PERMISSION INFORMATION
Permission for field surveys was granted by the Qingshuihe Nature Reserve.
SUPPEMENTARY DATA
Supplementary data to this article can be found online.
COMPETING INTERESTS
The authors declare that they have no competing interests.
AUTHORS’ CONTRIBUTIONS
G.H.Y.and L.N.D.conceived and designed this study.G.H.Y.and J.S.W.conducted the field surveys.J.Z.and J.S.W.performed the experiments.J.Z.measured the specimens,analyzed the data, and wrote the manuscript.All authors read and approved the final version of the manuscript.
ACKNOWLEDGEMENTS
We thank Prof.Si-Min Lin for the donation ofK.idiootocusspecimens collected from Taiwan, China, and Rui Cheng for assistance with laboratory work.
猜你喜欢
杂志排行

Zoological Research的其它文章
- Flexible breeding performance under unstable climatic conditions in a tropical passerine in Southwest China
- Comparative transcriptomics highlights convergent evolution of energy metabolic pathways in group-living spiders
- Long-term trends in a forest ungulate community: park establishment increases numbers, but poaching is a constant threat
- Molecular phylogeny of the genus Muntiacus with special emphasis on the phylogenetic position of Muntiacus gongshanensis
- Parasitism is always costly to the host
- Yunnanilus chuanheensis, a new loach species(Cypriniformes: Nemacheilidae) from the upper Lixianjiang River in Yunnan, China